
Transcriptional Repression of Raf Kinase Inhibitory Protein Gene by Metadherin during Cancer Progression

 ,
,  , ,
, ,  , , and
, , and
Abstract
:1. Introduction
2. Results
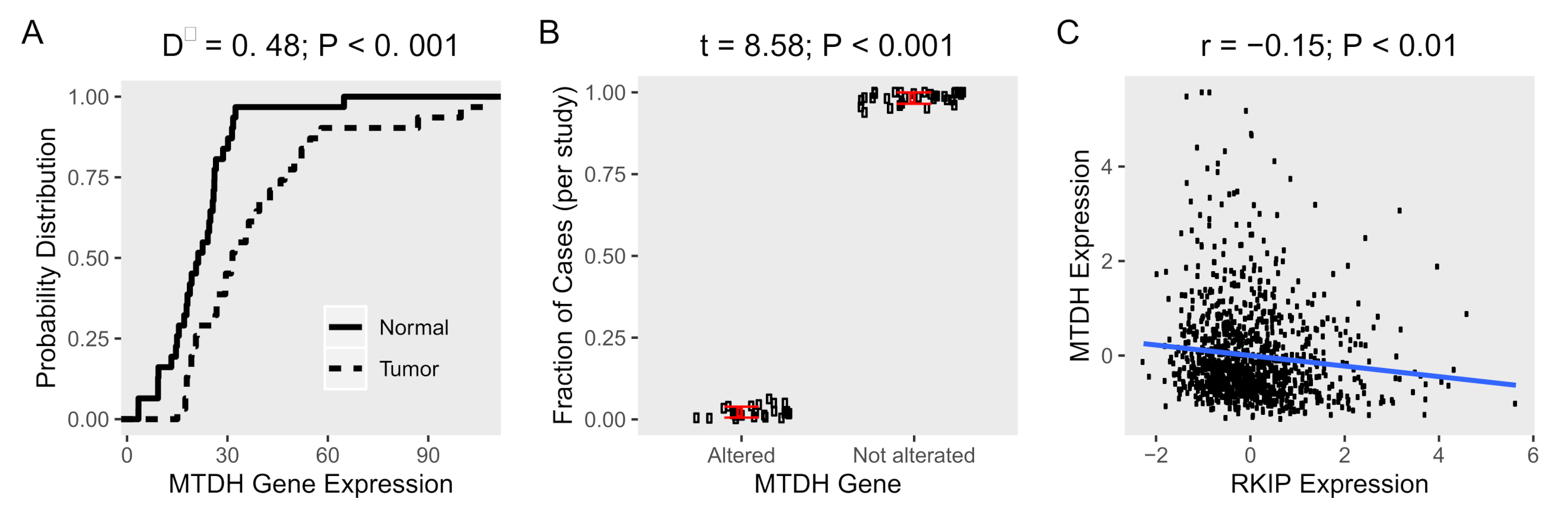
2.1. MTDH Is Upregulated in Cancer Tissues with Low Frequency Of Genetic Alterations
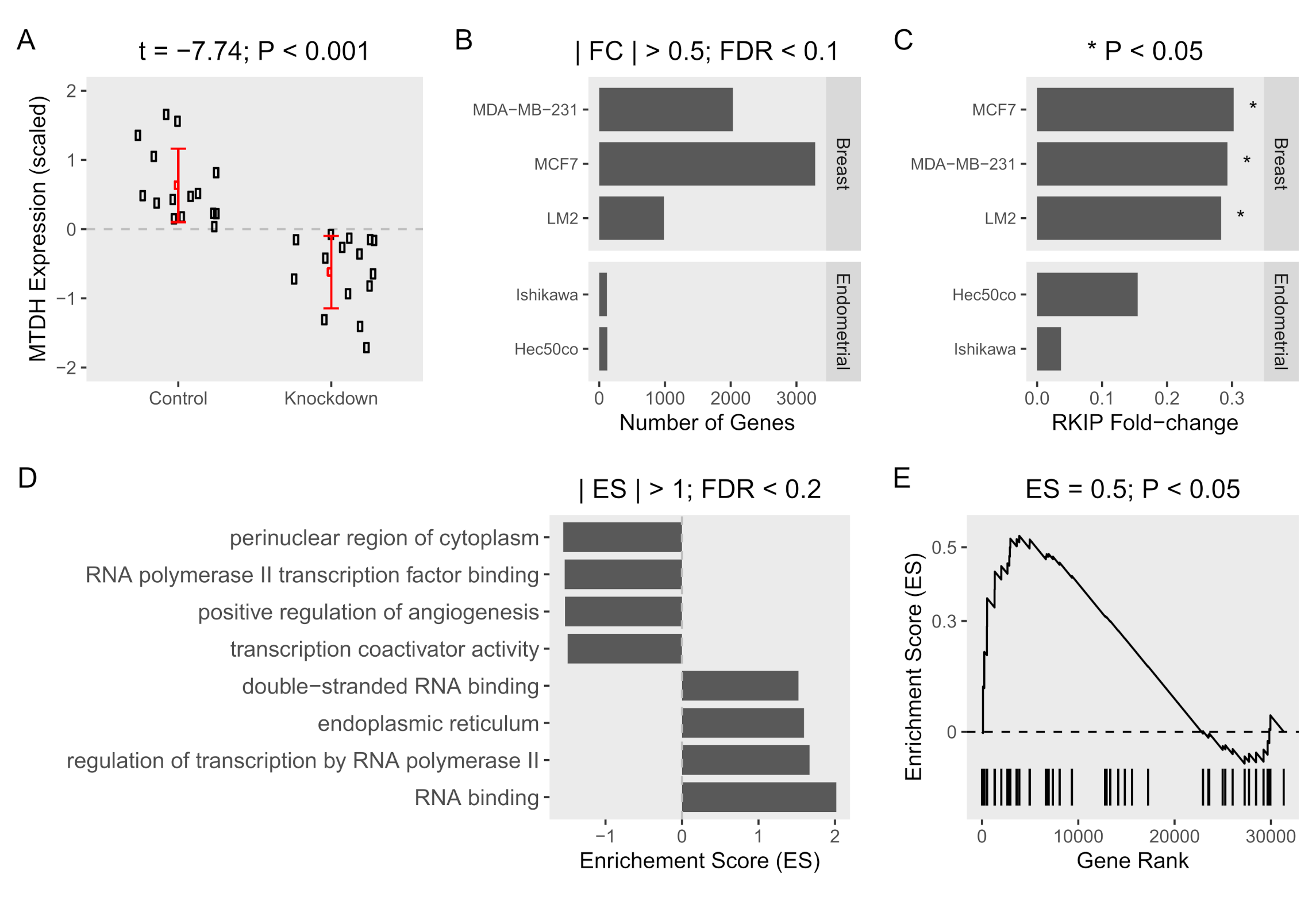
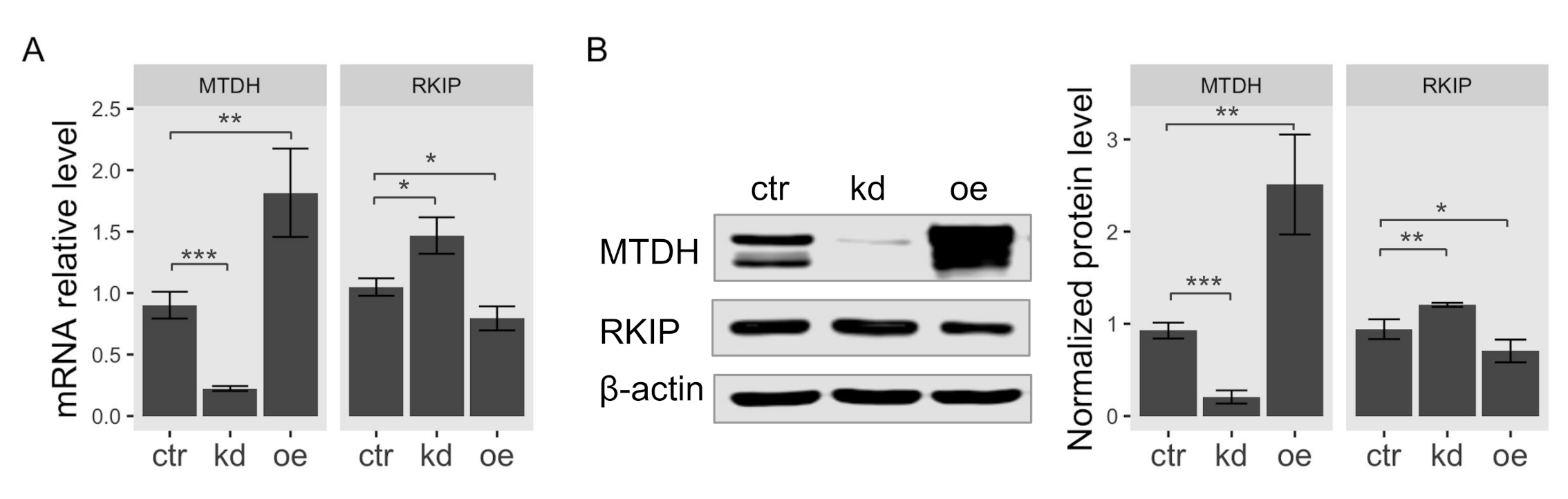
2.2. MTDH Knockdown Induces RKIP Expression in Breast Cancer Cell Lines
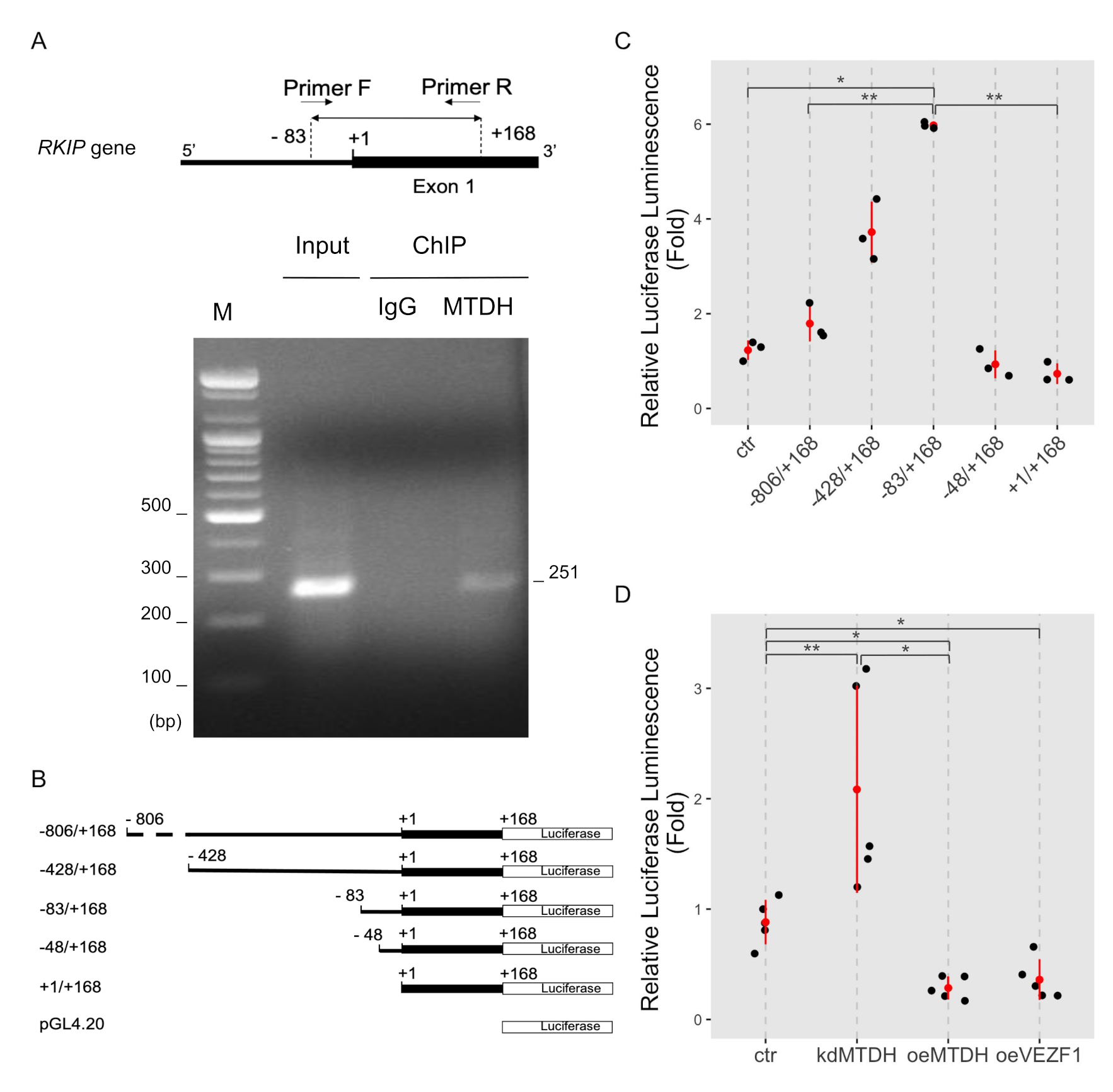
2.3. MTDH Regulates the Transcription of the RKIP Gene
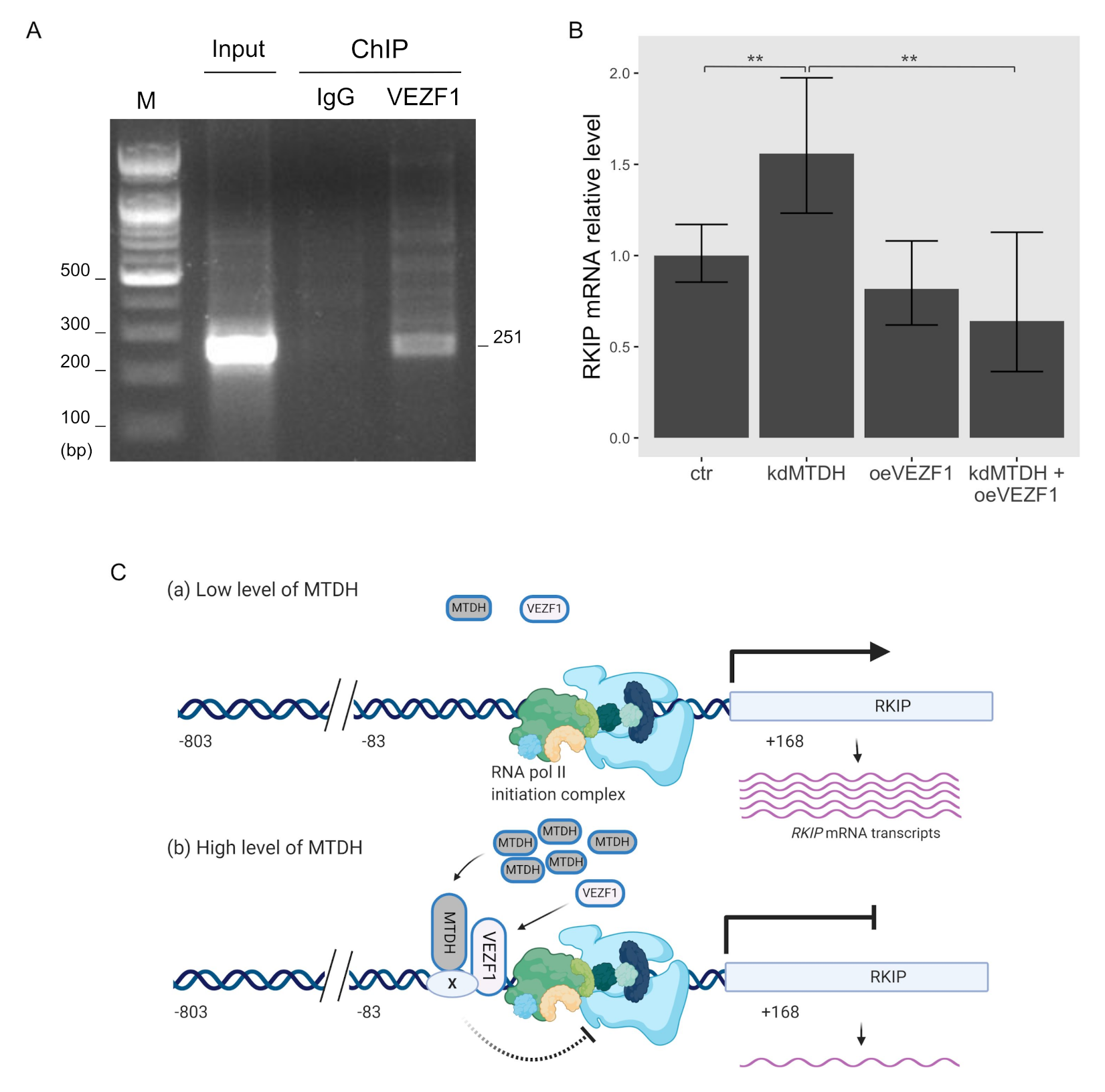
2.4. MTDH Binds to the RKIP Promoter
2.5. Functional Association between MTDH and VEZF1 during Transcriptional Activation of RKIP Gene
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Gene Expression Analysis of MTDH-Knockout Cells

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
4.3. MTDH Gene Expression Analysis in Cancer Tissues
4.4. Dual Cross-Linking Chromatin Immunoprecipitation Assay
4.5. Cell Culture and Transfection
4.6. Production of Recombinant Lentiviral Particles and Infection
4.7. Western Blot Analysis
4.8. RNA Extraction and RT-qPCR Analysis
4.9. Luciferase Promoter Assay
4.10. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yeung, K.; Seitz, T.; Li, S.; Janosch, P.; McFerran, B.; Kaiser, C.; Fee, F.; Katsanakis, K.D.; Rose, D.W.; Mischak, H.; et al. Suppression of Raf-1 kinase activity and MAP kinase signalling by RKIP. Nature 1999, 401, 173–177. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, D.; Bai, Y.; Wang, Z.; Beach, S.; Mott, S.; Roy, R.; Braastad, C.; Sun, Y.; Mukhopadhyay, A.; Aggarwal, B.B.; et al. RKIP Sensitizes Prostate and Breast Cancer Cells to Drug-induced Apoptosis. J. Biol. Chem. 2004, 279, 17515–17523. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Yeung, M.L.; Beach, S.; Shields, J.M.; Yeung, K.C. RKIP downregulates B-Raf kinase activity in melanoma cancer cells. Oncogene 2005, 24, 3535–3540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Funderburk, S.F.; Marcellino, B.K.; Yue, Z. Cell “Self-Eating” (Autophagy) Mechanism in Alzheimer’s Disease. Mt. Sinai J. Med. A J. Transl. Pers. Med. 2010, 77, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Park, S.G.; Jung, H.; Chi, S.W.; Yu, D.Y.; Lee, S.C.; Bae, K.H. RKIP downregulation induces the HBx-mediated Raf-1 mitochondrial translocation. J. Microbiol. Biotechnol. 2011, 21, 525–528. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.z.; Kang, D.c.; Chen, Y.; Pekarskaya, O.; Chao, W.; Volsky, D.J.; Fisher, P.B. Identification and cloning of human astrocyte genes displaying elevated expression after infection with HIV-1 or exposure to HIV-1 envelope glycoprotein by rapid subtraction hybridization, RaSH. Oncogene 2002, 21, 3592–3602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, G.; Chong, R.A.; Yang, Q.; Wei, Y.; Blanco, M.A.; Li, F.; Reiss, M.; Au, J.L.S.; Haffty, B.G.; Kang, Y.; et al. MTDH Activation by 8q22 Genomic Gain Promotes Chemoresistance and Metastasis of Poor-Prognosis Breast Cancer. Cancer Cell 2009, 15, 9–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Fu, D.; Hu, G. Metadherin: An emerging key regulator of the malignant progression of multiple cancers. Thorac. Cancer 2011, 2, 143–148. [Google Scholar] [CrossRef]
- Song, H.; Li, C.; Lu, R.; Zhang, Y.; Geng, J. Expression of astrocyte elevated gene-1: A novel marker of the pathogenesis, progression, and poor prognosis for endometrial cancer. Int. J. Gynecol. Cancer 2010, 20, 19. [Google Scholar] [CrossRef]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [Green Version]
- Dhiman, G.; Srivastava, N.; Goyal, M.; Rakha, E.; Lothion-Roy, J.; Mongan, N.P.; Miftakhova, R.R.; Khaiboullina, S.F.; Rizvanov, A.A.; Baranwal, M. Metadherin: A therapeutic target in multiple cancers. Front. Oncol. 2019. [Google Scholar] [CrossRef]
- Hou, Y.; Yu, L.; Mi, Y.; Zhang, J.; Wang, K.; Hu, L. Association of MTDH immunohistochemical expression with metastasis and prognosis in female reproduction malignancies: A systematic review and meta-analysis. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Z.; Li, C.; Kang, B.; Gao, G.; Li, C.; Zhang, Z. GEPIA: A web server for cancer and normal gene expression profiling and interactive analyses. Nucleic Acids Res. 2017, 45, W98–W102. [Google Scholar] [CrossRef] [Green Version]
- Zou, Q.; Wu, H.; Fu, F.; Yi, W.; Pei, L.; Zhou, M. RKIP suppresses the proliferation and metastasis of breast cancer cell lines through up-regulation of miR-185 targeting HMGA2. Arch. Biochem. Biophys. 2016, 610, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Noh, H.S.; Hah, Y.S.; Zada, S.; Ha, J.H.; Sim, G.; Hwang, J.S.; Lai, T.H.; Nguyen, H.Q.; Park, J.Y.; Kim, H.J.; et al. PEBP1, a RAF kinase inhibitory protein, negatively regulates starvation-induced autophagy by direct interaction with LC3. Autophagy 2016, 12, 2183–2196. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, O.; Qin, J.; Liu, S.; Sun, S.; Liu, H.; Kuang, J.; Jiang, G.; Zhang, W. Cis-acting elements and trans-acting factors in the transcriptional regulation of Raf kinase inhibitory protein expression. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Lai, T.H.; Zada, S.; Hwang, J.S.; Pham, T.M.; Yun, M.; Kim, D.R. Functional linkage of RKIP to the epithelial to mesenchymal transition and autophagy during the development of prostate cancer. Cancers 2018, 10, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, D.M.; Ruoslahti, E. Metadherin, a cell surface protein in breast tumors that mediates lung metastasis. Cancer Cell 2004, 5, 365–374. [Google Scholar] [CrossRef] [Green Version]
- Guo, F.; Wan, L.; Zheng, A.; Stanevich, V.; Wei, Y.; Satyshur, K.A.; Shen, M.; Lee, W.; Kang, Y.; Xing, Y. Structural Insights into the Tumor-Promoting Function of the MTDH-SND1 Complex. Cell Rep. 2014, 8, 1704–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emdad, L.; Lee, S.G.; Su, Z.Z.; Jeon, H.Y.; Boukerche, H.; Sarkar, D.; Fisher, P.B. Astrocyte elevated gene-1 (AEG-1) functions as an oncogene and regulates angiogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 21300–5. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, D.; Eun, S.P.; Emdad, L.; Lee, S.G.; Su, Z.Z.; Fisher, P.B. Molecular basis of nuclear factor-κB activation by astrocyte elevated gene-1. Cancer Res. 2008, 68, 1478–1484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emdad, L.; Sarkar, D.; Su, Z.z.Z.Z.z.; Randolph, A.; Boukerche, H.; Valerie, K.; Fisher, P.B.; Eun, S.P.; Emdad, L.; Lee, S.g.G.; et al. Activation of the nuclear factor κB pathway by astrocyte elevated gene-1: Implications for tumor progression and metastasis. Cancer Res. 2011, 66, 1509–1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanco, M.A.; Alečković, M.; Hua, Y.; Li, T.; Wei, Y.; Xu, Z.; Cristea, I.M.; Kang, Y. Identification of staphylococcal nuclease domain-containing 1 (SND1) as a metadherin-interacting protein with metastasis-promoting functions. J. Biol. Chem. 2011, 286, 19982–19992. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Du, X.; Zang, L.; Song, N.; Yang, T.; Dong, R.; Wu, T.; He, X.; Lu, J. Prognostic impact of Metadherin-SND1 interaction in colon cancer. Mol. Biol. Rep. 2012, 39, 10497–10504. [Google Scholar] [CrossRef]
- Yoo, B.K.; Emdad, L.; Lee, S.G.; Su, Z.Z.; Santhekadur, P.; Chen, D.; Gredler, R.; Fisher, P.B.; Sarkar, D. Astrocyte elevated gene-1 (AEG-1): A multifunctional regulator of normal and abnormal physiology. Pharmacol. Ther. 2011, 130, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.; Drapkin, R.; Yalamanchili, R.; Mosialos, G.; Kieff, E. The Epstein-Barr virus nuclear protein 2 acidic domain forms a complex with a novel cellular coactivator that can interact with TFIIE. Mol. Cell. Biol. 1995, 15, 4735–4744. [Google Scholar] [CrossRef] [Green Version]
- Beach, S.; Tang, H.; Park, S.; Dhillon, A.S.; Keller, E.T.; Kolch, W.; Yeung, K.C. Snail is a repressor of RKIP transcription in metastatic prostate cancer cells. Oncogene 2008, 27, 2243–2248. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; He, K.; Yan, S.; Yang, Y.; Gao, X.; Zhang, M.; Xia, Z.; Huang, Z.; Huang, S.; Zhang, N. Metadherin/astrocyte elevated gene-1 positively regulates the stability and function of forkhead box M1 during tumorigenesis. Neuro-Oncology 2017, 19, 352–363. [Google Scholar] [CrossRef] [Green Version]
- Bruderer, M.; Alini, M.; Stoddart, M.J. Role of HOXA9 and VEZF1 in endothelial biology. J. Vasc. Res. 2013, 50, 265–278. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Yang, Q.; Norvil, A.B.; Sherris, D.; Gowher, H. Characterization of small molecules inhibiting the pro-angiogenic activity of the zinc finger transcription factor vezf1. Molecules 2018, 23, 1615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyashita, H.; Sato, Y. Metallothionein 1 is a downstream target of vascular endothelial zinc finger 1 (VEZF1) in endothelial cells and participates in the regulation of angiogenesis. Endothel. J. Endothel. Cell Res. 2005, 12, 163–170. [Google Scholar] [CrossRef]
- AlAbdi, L.; He, M.; Yang, Q.; Norvil, A.B.; Gowher, H. The transcription factor Vezf1 represses the expression of the antiangiogenic factor Cited2 in endothelial cells. J. Biol. Chem. 2018, 293, 11109–11118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keller, E.T. Metastasis suppressor genes: A role for raf kinase inhibitor protein (RKIP). Anti-Cancer Drugs 2004, 15, 663–669. [Google Scholar] [CrossRef]
- Feng, L.; Zhang, C.; Liu, G.; Wang, F. RKIP negatively regulates the glucose induced angiogenesis and endothelial-mesenchymal transition in retinal endothelial cells. Exp. Eye Res. 2019, 189, 107851. [Google Scholar] [CrossRef]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. Limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 43, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gollavilli, P.; Kanugula, A.; Pemmasani, S.; Kotamraju, S. A Newly Identified Spliced Isoform of Metadherin Differently Regulates the Global Transcriptomic Profile in MCF-7 Breast Cancer Cells; CSIR-Indian Institute of Chemical Technology: Secunderabad, India, 2014. [Google Scholar]
- Meng, X.; Brachova, P.; Yang, S.; Xiong, Z.; Zhang, Y.; Thiel, K.W.; Leslie, K.K. Knockdown of mtdh sensitizes endometrial cancer cells to cell death induction by death receptor ligand trail and HDAC inhibitor LBH589 Co-treatment. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.; Kim, D.R. pcr: An R package for quality assessment, analysis and testing of qPCR data. PeerJ 2018, 2018. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, T.H.; Ahmed, M.; Hwang, J.S.; Zada, S.; Pham, T.M.; Elashkar, O.; Kim, D.R. Transcriptional Repression of Raf Kinase Inhibitory Protein Gene by Metadherin during Cancer Progression. Int. J. Mol. Sci. 2021, 22, 3052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063052
Lai TH, Ahmed M, Hwang JS, Zada S, Pham TM, Elashkar O, Kim DR. Transcriptional Repression of Raf Kinase Inhibitory Protein Gene by Metadherin during Cancer Progression. International Journal of Molecular Sciences. 2021; 22(6):3052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063052
Chicago/Turabian StyleLai, Trang Huyen, Mahmoud Ahmed, Jin Seok Hwang, Sahib Zada, Trang Minh Pham, Omar Elashkar, and Deok Ryong Kim. 2021. "Transcriptional Repression of Raf Kinase Inhibitory Protein Gene by Metadherin during Cancer Progression" International Journal of Molecular Sciences 22, no. 6: 3052. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063052