
Analysis of the Structure and Biosynthesis of the Lipopolysaccharide Core Oligosaccharide of Pseudomonas syringae pv. tomato DC3000

 , and
, and
Abstract
:1. Introduction
2. Results
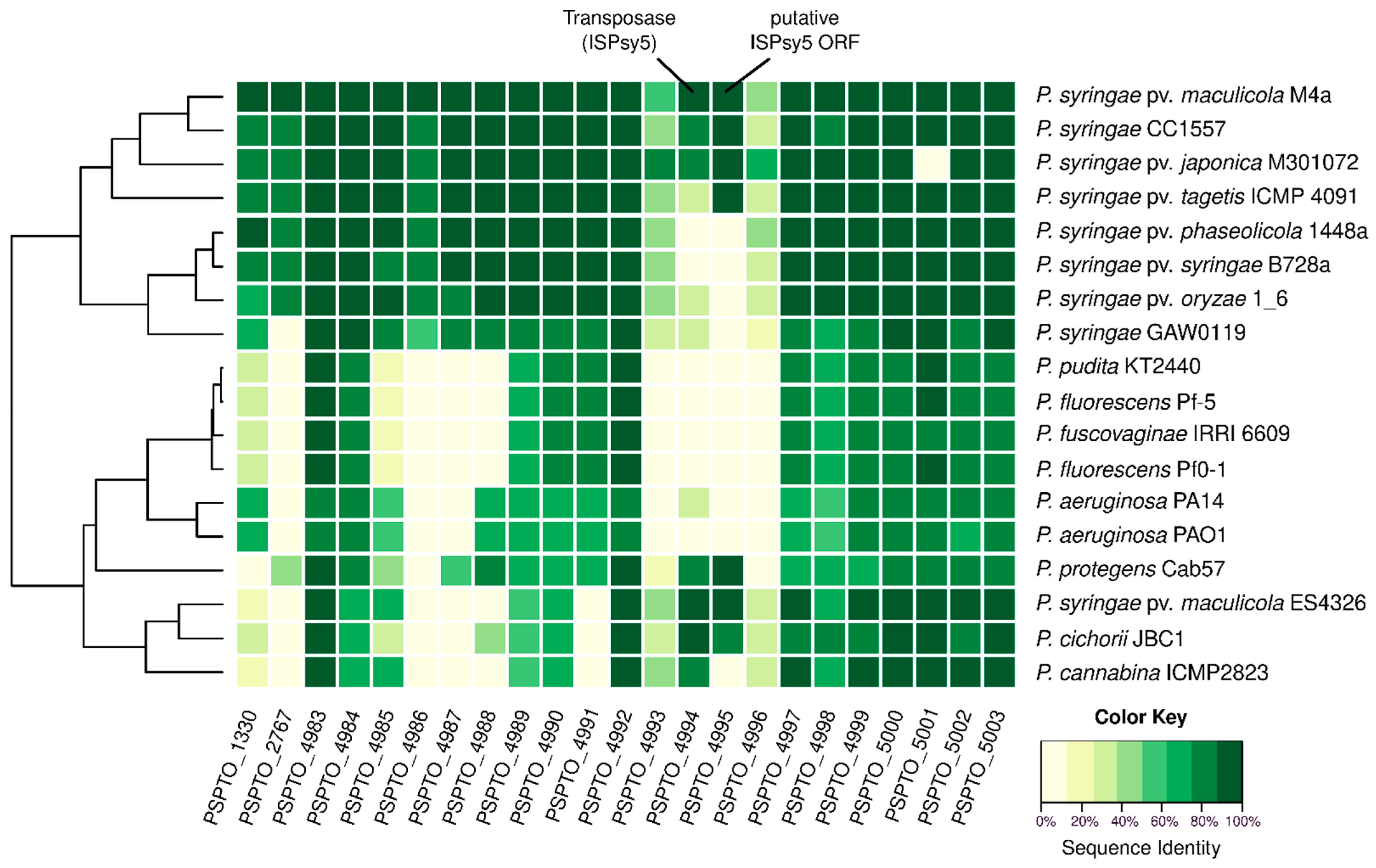
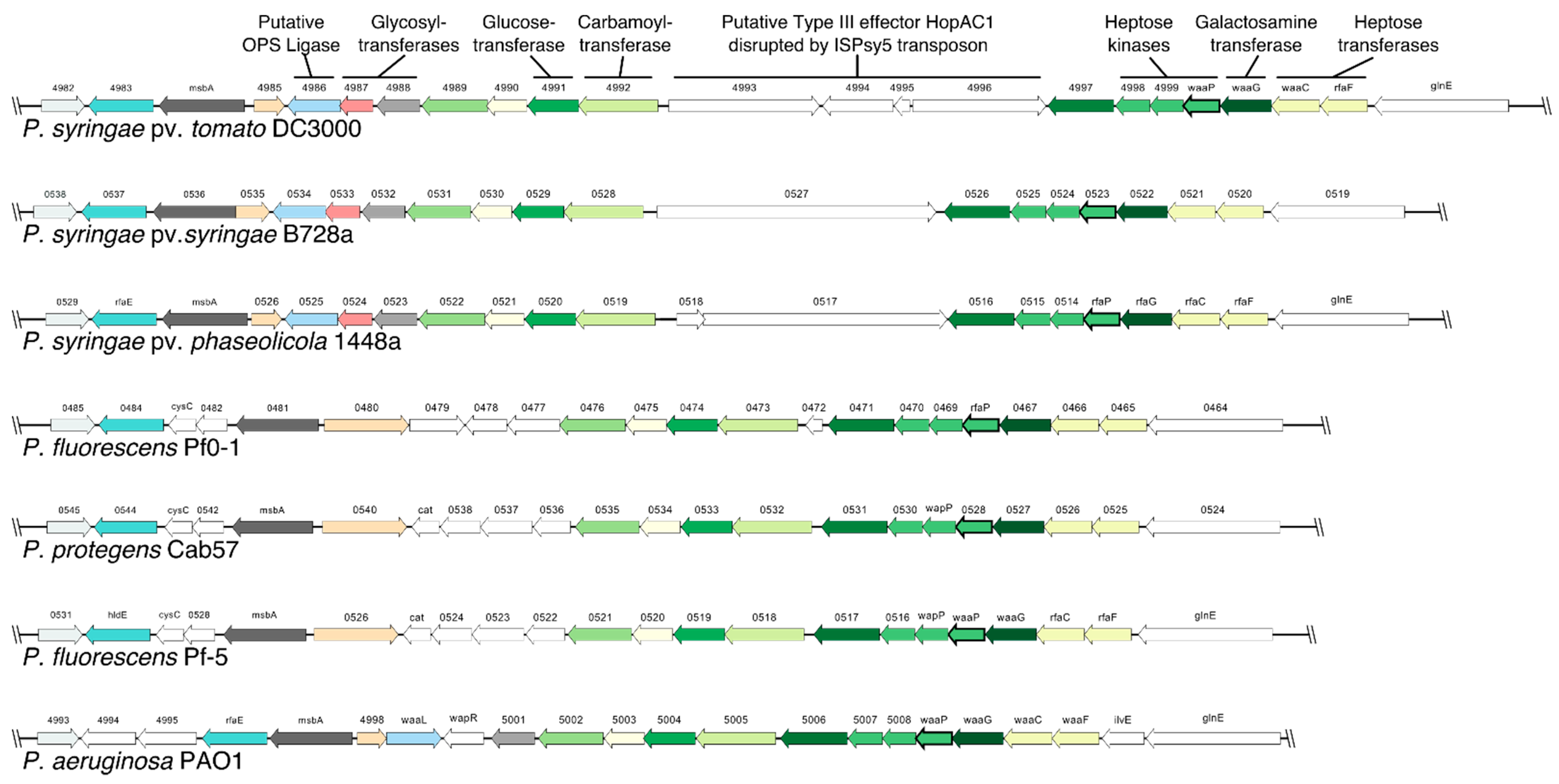
2.1. Pst DC3000 Core-OS Gene Cluster Contains an Insertion Sequence Element
2.2. Genes Involved in Synthesis of the Inner Core-OS Are Conserved in Pseudomonas
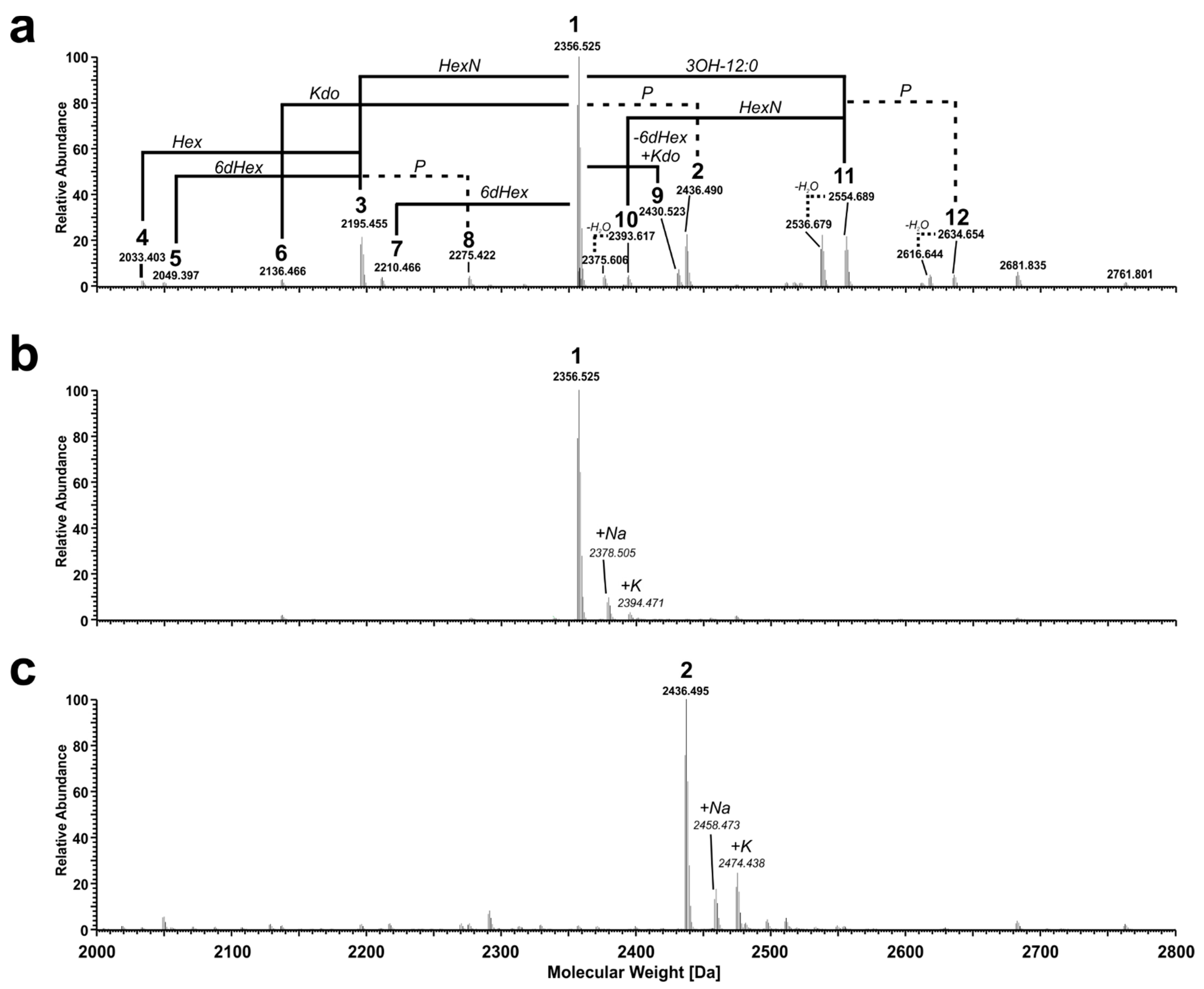
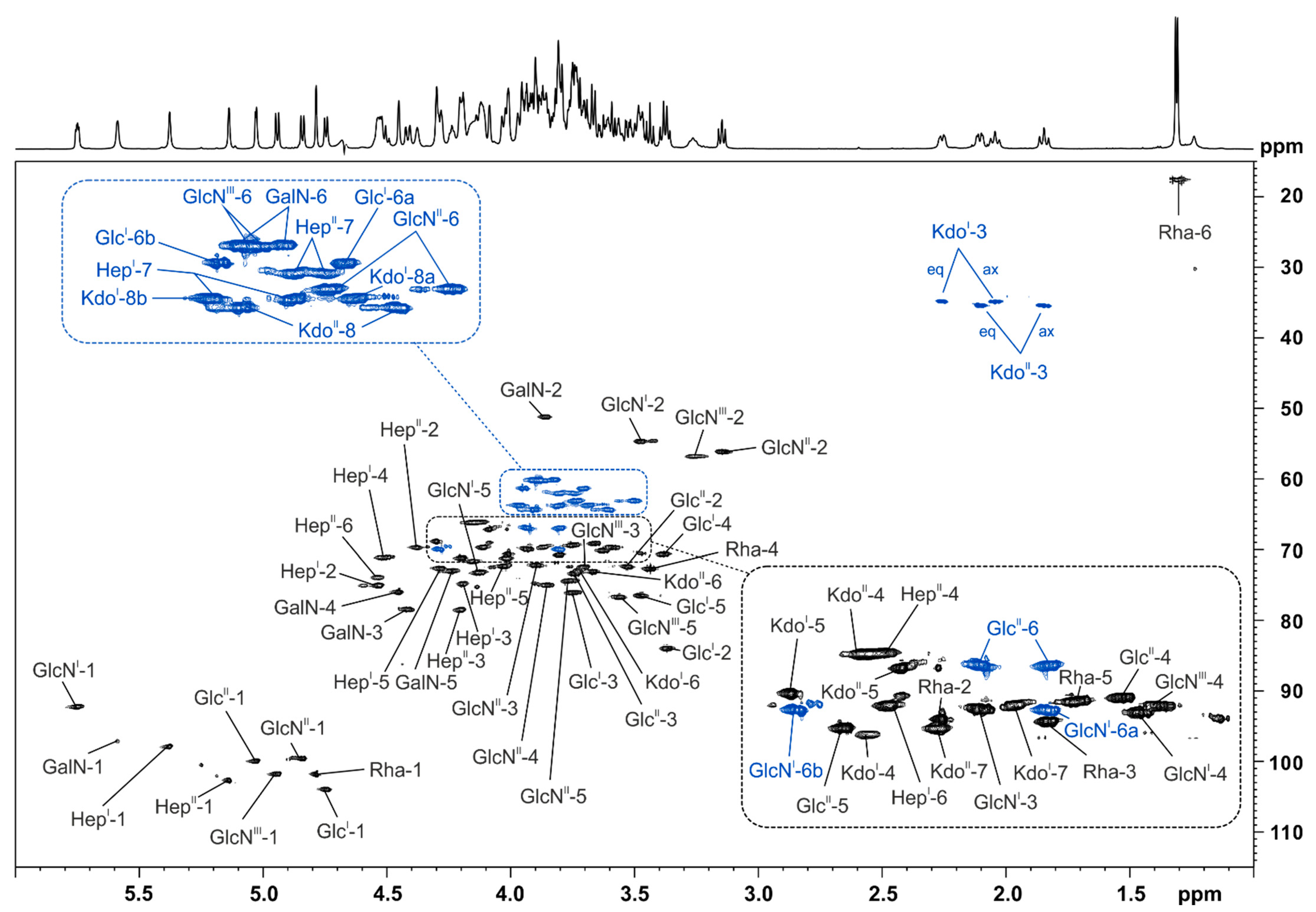
2.3. Structural Analysis of the Pst DC3000 LPS Core-Oligosaccharide
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. Hot Phenol-Water and Phenol-Chloroform-Petroleum Ether Extraction of LPS
4.3. Alkaline Degradation of the Lipopolysaccharide
4.4. Mild-Acid Degradation of the Lipopolysaccharide
4.5. NMR Spectroscopy
4.6. Mass Spectrometry
4.7. Sequence Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Microbiol. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Baltrus, D.A.; McCann, H.C.; Guttman, D.S. Evolution, genomics and epidemiology of Pseudomonas syringae: Challenges in bacterial molecular plant pathology. Mol. Plant Pathol. 2017, 18, 152–168. [Google Scholar] [CrossRef] [Green Version]
- Raetz, C.R.H.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexander, C.; Rietschel, E.T. Bacterial lipopolysaccharides and innate immunity. J. Endotoxin Res. 2001, 7, 167–202. [Google Scholar] [CrossRef]
- Lerouge, I.; Vanderleyden, J. O-antigen structural variation: Mechanisms and possible roles in animal/plant-microbe interactions. FEMS Microbiol. Rev. 2002, 26, 17–47. [Google Scholar] [CrossRef] [Green Version]
- Molinaro, A.; Newman, M.-A.; Lanzetta, R.; Parrilli, M. The structures of lipopolysaccharides from plant-associated Gram-negative bacteria. Eur. J. Org. Chem. 2009, 2009, 5887–5896. [Google Scholar] [CrossRef]
- Trent, M.S.; Stead, C.M.; Tran, A.X.; Hankins, J.V. Diversity of endotoxin and its impact on pathogenesis. J. Endotoxin Res. 2006, 12, 205–223. [Google Scholar] [CrossRef]
- Whitfield, C.; Trent, M.S. Biosynthesis and export of bacterial lipopolysaccharides. Annu. Rev. Biochem. 2014, 83, 99–128. [Google Scholar] [CrossRef]
- Lam, J.S.; Taylor, V.L.; Islam, S.T.; Hao, Y.; Kocíncová, D. Genetic and functional diversity of Pseudomonas aeruginosa lipopolysaccharide. Front. Microbiol. 2011, 2, 118. [Google Scholar] [CrossRef] [Green Version]
- Knirel, Y.A.; Bystrova, O.V.; Kocharova, N.A.; Zähringer, U.; Pier, G.B. Conserved and variable structural features in the lipopolysaccharide of Pseudomonas aeruginosa. J. Endotoxin Res. 2006, 12, 324–336. [Google Scholar] [CrossRef]
- Ernst, R.K.; Hajjar, A.M.; Tsai, J.H.; Moskowitz, S.M.; Wilson, C.B.; Miller, S.I. Pseudomonas aeruginosa lipid A diversity and its recognition by Toll-like receptor 4. J. Endotoxin Res. 2003, 9, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Frirdich, E.; Whitfield, C. Lipopolysaccharide inner core oligosaccharide structure and outer membrane stability in human pathogens belonging to the Enterobacteriaceae. J. Endotoxin Res. 2005, 11, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Kutschera, A.; Ranf, S. The multifaceted functions of lipopolysaccharide in plant-bacteria interactions. Biochimie 2019, 159, 93–98. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Owen, K.A.; Ly, K.T.; Park, D.; Black, S.G.; Wilson, J.M.; Sifri, C.D.; Ravichandran, K.S.; Ernst, P.B.; Casanova, J.E. Brain angiogenesis inhibitor 1 (BAI1) is a pattern recognition receptor that mediates macrophage binding and engulfment of Gram-negative bacteria. Proc. Natl. Acad. Sci. USA 2011, 108, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, T.H.; Lee, M.M.; Yacono, P.W.; Cannon, C.L.; Gerçeker, A.A.; Golan, D.E.; Pier, G.B. CFTR is a pattern recognition molecule that extracts Pseudomonas aeruginosa LPS from the outer membrane into epithelial cells and activates NF-kB translocation. Proc. Natl. Acad. Sci. USA 2002, 99, 6907–6912. [Google Scholar] [CrossRef] [Green Version]
- Silipo, A.; Molinaro, A.; Sturiale, L.; Dow, J.M.; Erbs, G.; Lanzetta, R.; Newman, M.-A.; Parrilli, M. The elicitation of plant innate immunity by lipooligosaccharide of Xanthomonas campestris. J. Biol. Chem. 2005, 280, 33660–33668. [Google Scholar] [CrossRef] [Green Version]
- Braun, S.G.; Meyer, A.; Holst, O.; Pühler, A.; Niehaus, K. Characterization of the Xanthomonas campestris pv. campestris lipopolysaccharide substructures essential for elicitation of an oxidative burst in tobacco cells. Mol. Plant Microbe Interact. 2005, 18, 674–681. [Google Scholar] [CrossRef] [Green Version]
- Needham, B.D.; Trent, M.S. Fortifying the barrier: The impact of lipid a remodelling on bacterial pathogenesis. Nat. Rev. Microbiol. 2013, 11, 467–481. [Google Scholar] [CrossRef]
- Kooistra, O.; Lüneberg, E.; Lindner, B.; Knirel, Y.A.; Frosch, M.; Zähringer, U. Complex O-acetylation in Legionella pneumophila serogroup 1 lipopolysaccharide. Evidence for two genes involved in 8-O-acetylation of legionaminic acid. Biochemistry 2001, 40, 7630–7640. [Google Scholar] [CrossRef]
- Kocincova, D.; Lam, J.S. Structural diversity of the core oligosaccharide domain of Pseudomonas aeruginosa lipopolysaccharide. Biochem. Mosc. 2011, 76, 755–760. [Google Scholar] [CrossRef]
- Bhasin, N.; Albus, A.; Michon, F.; Livolsi, P.J.; Park, J.S.; Lee, J.C. Identification of a gene essential for O-acetylation of the Staphylococcus aureus type 5 capsular polysaccharide. Mol. Microbiol. 1998, 27, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Amor, K.; Heinrichs, D.E.; Frirdich, E.; Ziebell, K.; Johnson, R.P.; Whitfield, C. Distribution of core oligosaccharide types in lipopolysaccharides from Escherichia coli. Infect. Immun. 2000, 68, 1116–1124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, J.D.; Kocíncová, D.; Westman, E.L.; Lam, J.S. Review: Lipopolysaccharide biosynthesis in Pseudomonas aeruginosa. Innate Immun. 2009, 15, 261–312. [Google Scholar] [CrossRef]
- De Castro, C.; Molinaro, A.; Nunziata, R.; Lanzetta, R.; Parrilli, M.; Holst, O. A novel core region, lacking heptose and phosphate, of the lipopolysaccharide from the Gram-negative bacterium Pseudomonas cichorii (Pseudomonadaceae RNA group 1). Eur. J. Org. Chem. 2004, 2004, 2427–2435. [Google Scholar] [CrossRef]
- Walsh, A.G.; Matewish, M.J.; Burrows, L.L.; Monteiro, M.A.; Perry, M.B.; Lam, J.S. Lipopolysaccharide core phosphates are required for viability and intrinsic drug resistance in Pseudomonas aeruginosa. Mol. Microbiol. 2000, 35, 718–727. [Google Scholar] [CrossRef]
- Zdorovenko, E.L.; Vinogradov, E.; Zdorovenko, G.M.; Lindner, B.; Bystrova, O.V.; Shashkov, A.S.; Rudolph, K.; Zähringer, U.; Knirel, Y.A. Structure of the core oligosaccharide of a rough-type lipopolysaccharide of Pseudomonas syringae pv. phaseolicola. Eur. J. Biochem. FEBS 2004, 271, 4968–4977. [Google Scholar] [CrossRef] [Green Version]
- Zdorovenko, G.M.; Varbanets, L.D.; Zdorovenko, E.L.; Vinarskaya, N.V.; Yakovleva, L.M. Chemical and biological characterization of lipopolysaccharides from the Pseudomonas syringae pv. maculicola IMV 381 collection culture and its dissociants. Microbiology 2004, 73, 678–688. [Google Scholar] [CrossRef]
- Zdorovenko, G.M.; Zdorovenko, E.L.; Varbanets, L.D. Composition, structure, and biological properties of lipopolysaccharides from different strains of Pseudomonas syringae pv. atrofaciens. Microbiology 2007, 76, 683–697. [Google Scholar] [CrossRef]
- Kutschera, A.; Schombel, U.; Wröbel, M.; Gisch, N.; Ranf, S. Loss of wbpL disrupts O-polysaccharide synthesis and impairs virulence of plant-associated Pseudomonas strains. Mol. Plant Pathol. 2019, 20, 1535–1549. [Google Scholar] [CrossRef] [Green Version]
- Knirel, Y.A.; Bystrova, O.V.; Shashkov, A.S.; Lindner, B.; Kocharova, N.A.; Senchenkova, S.N.; Moll, H.; Zähringer, U.; Hatano, K.; Pier, G.B. Structural analysis of the lipopolysaccharide core of a rough, cystic fibrosis isolate of Pseudomonas aeruginosa. Eur. J. Biochem. FEBS 2001, 268, 4708–4719. [Google Scholar] [CrossRef] [Green Version]
- Silipo, A.; Leone, S.; Molinaro, A.; Lanzetta, R.; Parrilli, M. The structure of the phosphorylated carbohydrate backbone of the lipopolysaccharide of the phytopathogen bacterium Pseudomonas tolaasii. Carbohydr. Res. 2004, 339, 2241–2248. [Google Scholar] [CrossRef] [PubMed]
- DeLucia, A.M.; Six, D.A.; Caughlan, R.E.; Gee, P.; Hunt, I.; Lam, J.S.; Dean, C.R. Lipopolysaccharide (LPS) inner-core phosphates are required for complete LPS synthesis and transport to the outer membrane in Pseudomonas aeruginosa PAO1. mBio 2011, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [Green Version]
- Jimenez-Barbero, J.; de Castro, C.; Evidente, A.; Molinaro, A.; Parrilli, M.; Surico, G. Structural determination of the O-specific chain of the lipopolysaccharide from Pseudomonas cichorii. Eur. J. Org. Chem. 2002, 2002, 1770–1775. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Helbig, J.H.; Zähringer, U. Structure of a decasaccharide isolated by mild acid degradation and dephosphorylation of the lipopolysaccharide of Pseudomonas fluorescens strain ATCC 49271. Carbohydr. Res. 1996, 283, 129–139. [Google Scholar] [CrossRef]
- Bystrova, O.V.; Lindner, B.; Moll, H.; Kocharova, N.A.; Knirel, Y.A.; Zähringer, U.; Pier, G.B. Structure of the lipopolysaccharide of Pseudomonas aeruginosa O-12 with a randomly O-acetylated core region. Carbohydr. Res. 2003, 338, 1895–1905. [Google Scholar] [CrossRef]
- Bystrova, O.V.; Knirel, Y.A.; Lindner, B.; Kocharova, N.A.; Kondakova, A.N.; Zähringer, U.; Pier, G.B. Structures of the core oligosaccharide and O-units in the R- and SR-type lipopolysaccharides of reference strains of Pseudomonas aeruginosa O-serogroups. FEMS Immunol. Med. Microbiol. 2006, 46, 85–99. [Google Scholar] [CrossRef] [Green Version]
- Wilkinson, S.G. 31P N.m.r. evidence for the presence of triphosphate residues in lipopolysaccharides from Pseudomonas aeruginosa. Biochem. J. 1981, 199, 833–835. [Google Scholar] [CrossRef] [Green Version]
- Schechter, L.M.; Vencato, M.; Jordan, K.L.; Schneider, S.E.; Schneider, D.J.; Collmer, A. Multiple approaches to a complete inventory of Pseudomonas syringae pv. tomato DC3000 type III secretion system effector proteins. Mol. Plant Microbe Interact. 2006, 19, 1180–1192. [Google Scholar] [CrossRef] [Green Version]
- Kutschera, A.; Dawid, C.; Gisch, N.; Schmid, C.; Raasch, L.; Gerster, T.; Schäffer, M.; Smakowska-Luzan, E.; Belkhadir, Y.; Vlot, A.C.; et al. Bacterial medium-chain 3-hydroxy fatty acid metabolites trigger immunity in Arabidopsis plants. Science 2019, 364, 178–181. [Google Scholar] [CrossRef]
- Ranf, S.; Gisch, N.; Schäffer, M.; Illig, T.; Westphal, L.; Knirel, Y.A.; Sánchez-Carballo, P.M.; Zähringer, U.; Hückelhoven, R.; Lee, J.; et al. A lectin S-domain receptor kinase mediates lipopolysaccharide sensing in Arabidopsis thaliana. Nat. Immunol. 2015, 16, 426–433. [Google Scholar] [CrossRef]
- Gottlieb, H.E.; Kotlyar, V.; Nudelman, A. NMR chemical shifts of common laboratory solvents as trace impurities. J. Org. Chem. 1997, 62, 7512–7515. [Google Scholar] [CrossRef]
- Oberto, J. SyntTax: A web server linking synteny to prokaryotic taxonomy. BMC Bioinform. 2013, 14, 4. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Identifier | Annotation 1 | Putative function in Pst DC3000 | PAO1 Equivalent | Putative/Proven Function in PAO1 | Identity % |
|---|---|---|---|---|---|
| Genes within core-OS cluster | |||||
| PSPTO_4983 | Lipopolysaccharide biosynthesis protein RfaE | Heptose biosynthesis | hldE (PA4996) | Heptose biosynthesis | 86.9 |
| PSPTO_4984 | Lipid A ABC transporter, ATP-binding/permease protein | Lipid-A:core-OS transport | msbA (PA4997) | Lipid-A:core-OS transport | 83.8 |
| PSPTO_4985 | Toluene tolerance protein | Unknown | PA4998 | Kinase | 55.6 |
| PSPTO_4986 | Membrane protein | Putative OPS ligase | waaL (PA4999) 2 | OPS ligase | No hit/19.4 3 |
| PSPTO_4987 | Hypothetical protein | WbcX-like glycosyltransferase | wapR (PA5000) 2 | Glycosyltransferase (Rha) | No hit/7.9 3 |
| PSPTO_4988 | Hypothetical protein | RfaB family glycosyltransferase | PA5001 | Glycosyltransferase | 73.6 |
| PSPTO_4989 | Hypothetical protein | PIG-L family deacetylase | PA5002 | Unknown | 65.5 |
| PSPTO_4990 | Hypothetical protein | GNAT family N-acetyltransferase | PA5003 | Unknown | 69.1 |
| PSPTO_4991 | Glycoside hydrolase family protein | Glycosyltransferase (GlcII) | wapH (PA5004) | Glycosyltransferase (GlcII) | 71.2 |
| PSPTO_4992 | Carbamoyltransferase family protein | Cm-(→7) carbamoyltransferase | wapO (PA5005) | Carbamoyltransferase | 86.5 |
| PSPTO_4993 | Hypothetical protein | Type III effector HopAC1 (segment) | No hit | ||
| PSPTO_4994 | ISPsy5, transposase | ISPsy5, transposase | No hit | ||
| PSPTO_4995 | ISPsy5, Orf1 | ISPsy5, Orf1 | No hit | ||
| PSPTO_4996 | Hypothetical protein | Type III effector HopAC1 (segment) | No hit | ||
| PSPTO_4997 | Hypothetical protein | Unknown | PA5006 | Kinase | 67.7 |
| PSPTO_4998 | Lipopolysaccharide biosynthesis protein | Heptose kinase | wapQ (PA5007) | Heptose kinase | 57.5 |
| PSPTO_4999 | Lipopolysaccharide core biosynthesis protein | Heptose kinase | wapP (PA5008) | Heptose kinase | 76.2 |
| PSPTO_5000 | Lipopolysaccharide core biosynthesis protein WaaP | Heptose kinase | waaP (PA5009) | Heptose kinase | 79.5 |
| PSPTO_5001 | Lipopolysaccharide core biosynthesis protein WaaG | α-GalN-(1→3) Glycosyltransferase | wapG (PA5010) | Glycosyltransferase (GalN) | 78.6 |
| PSPTO_5002 | Lipopolysaccharide heptosyltransferase | α-HepI-(1→5) Glycosyltransferase | waaC (PA5011) | Glycosyltransferase (HepI) | 74.9 |
| PSPTO_5003 | ADP-heptose--LPS heptosyltransferase II | α-HepII-(1→3) Glycosyltransferase | waaF (PA5012) | Glycosyltransferase (HepII) | 83.5 |
| Genes outside of the cluster | |||||
| PSPTO_1330 | Glycosyltransferase family protein | Glycosyltransferase α-l-Rha-(1→6) | migA (PA0705) | Glycosyltransferase (Rha1→6) | 63.9 |
| PSPTO_2767 | Lipopolysaccharide core biosynthesis domain protein | No hit | |||
| Molecule | Composition | Calculated Mass [Da] | Observed Mass [Da] | Error [ppm] | HPAEC * |
|---|---|---|---|---|---|
| 4 | Kdo2Hep2Hex16dHex1HexN3P5 | 2033.403 | 2033.403 | 0.0 | pool 4 |
| 5 | Kdo2Hep2Hex2HexN3P5 | 2049.398 | 2049.397 | −0.5 | pool 3 |
| 6 | Kdo1Hep2Hex26dHex1HexN4P5 | 2136.466 | 2136.466 | 0.0 | pool 1 |
| 3 | Kdo2Hep2Hex26dHex1HexN3P5 | 2195.456 | 2195.455 | −0.5 | pool 4 |
| 7 | Kdo2Hep2Hex2HexN4P5 | 2210.467 | 2210.466 | −0.5 | pool 3 |
| 8 | Kdo2Hep2Hex26dHex1HexN3P6 | 2275.422 | 2275.422 | 0.0 | pool 6 |
| 1 | Kdo2Hep2Hex26dHex1HexN4P5 | 2356.524 | 2356.525 | 0.4 | pool 2/3 |
| 10anh | Kdo2Hep2Hex26dHex1HexN3P5[12:0(3-OH)]–H2O | 2375.607 | 2375.606 | −0.4 | ** |
| 10 | Kdo2Hep2Hex26dHex1HexN3P5[12:0(3-OH)] | 2393.618 | 2393.617 | −0.4 | ** |
| 9 | Kdo3Hep2Hex2HexN4P5 | 2430.525 | 2430.523 | −0.8 | pool 4 |
| 2 | Kdo2Hep2Hex26dHex1HexN4P6 | 2436.491 | 2436.490 | −0.4 | pool 5 |
| 11anh | Kdo2Hep2Hex26dHex1HexN4P5[12:0(3-OH)]–H2O | 2536.676 | 2536.679 | 1.2 | ** |
| 11 | Kdo2Hep2Hex26dHex1HexN4P5[12:0(3-OH)] | 2554.686 | 2554.689 | 1.2 | ** |
| 12anh | Kdo2Hep2Hex26dHex1HexN4P6[12:0(3-OH)]–H2O | 2616.642 | 2616.644 | 0.8 | ** |
| 12 | Kdo2Hep2Hex26dHex1HexN4P6[12:0(3-OH)] | 2634.653 | 2634.654 | 0.4 | ** |
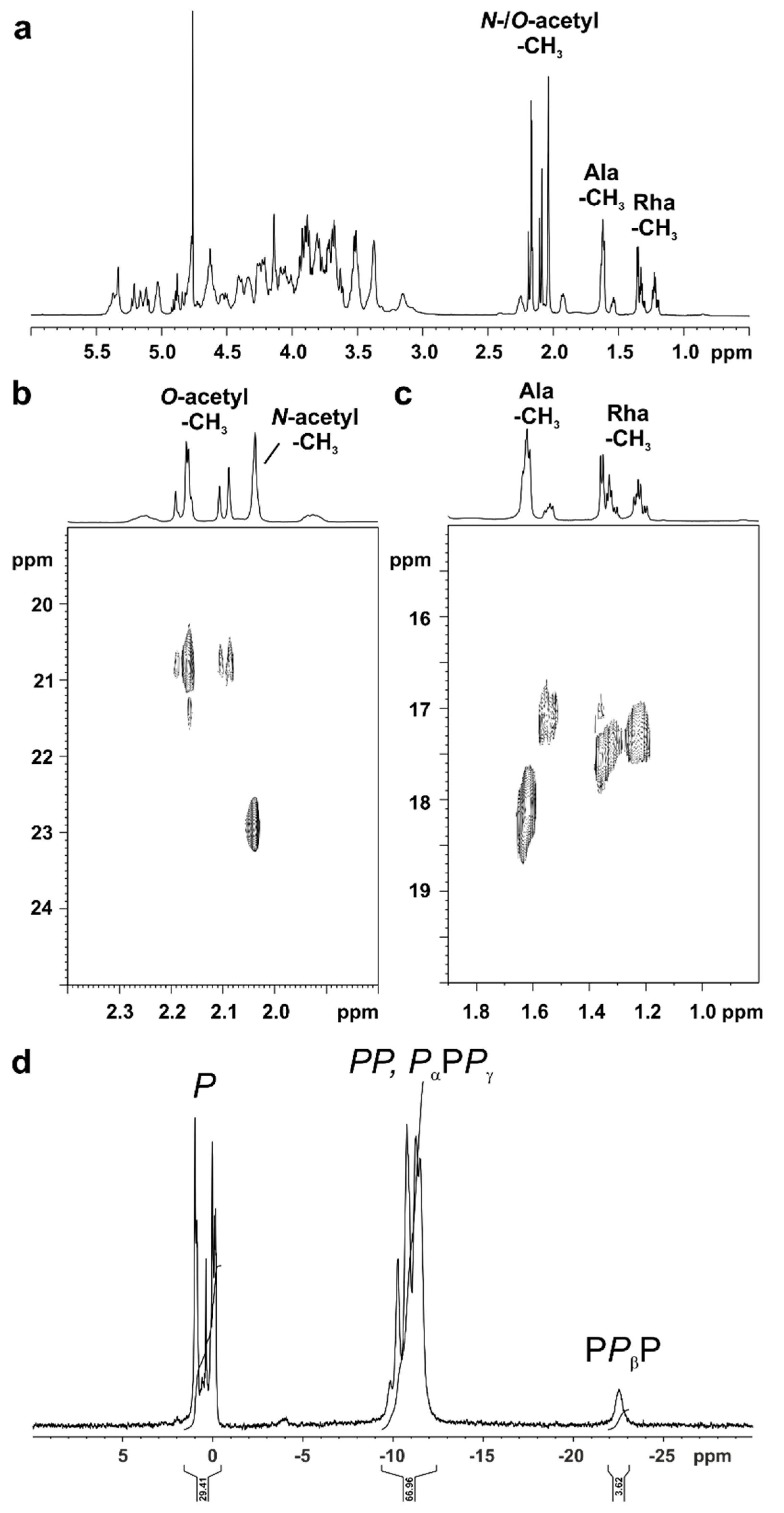
| Residue | H1 | H2 | H3 | H4 | H5 | H6a | H6b | H7a | H7b |
| →6)-α-GlcNI-(1→P | 5.77–5.73 | 3.50–3.45 | 3.96–3.91 | 3.65–3.60 | 4.15–4.10 | 3.83–3.78 | 4.31–4.27 | ||
| →6)-β-GlcNII4P-(1→ | 4.84 [d, 8.5 Hz] | 3.17–3.12 | 3.92–3.87 | 3.88–3.83 | 3.79–3.73 | 3.52–3.48 | 3.76-3.71 | ||
| →3)-α-HepI2P4P-(1→ | 5.39–5.36 | 4.54–4.51 | 4.22–4.17 | 4.54–4.48 | 4.30–4.26 | 4.14–4.09 | 3.83–3.79 | 3.99–3.94 | |
| →3)-α-HepII6P-(1→ | 5.15–5.12 | 4.39–4.36 | 4.21–4.18 | 4.13–4.09 | 4.05–4.00 | 4.56–4.50 | 3.77–3.71 | 3.82–3.77 | |
| →3,4)-α-GalN-(1→ | 5.60–5.57 | 3.88–3.83 | 4.43–4.39 | 4.46–4.44 | 4.24–4.21 | 3.84–3.80 | 3.93–3.88 | ||
| →2)-β-GlcI-(1→ | 4.75 [d, 7.9 Hz] | 3.38–3.34 | 3.77–3.73 | 3.40–3.36 | 3.50–3.45 | 3.73–3.69 | 3.97–3.93 | ||
| →6)-α-GlcII-(1→ | 5.03 [d, 7.9 Hz] | 3.55–3.50 | 3.76–3.72 | 3.68–3.63 | 4.22–4.18 | 3.81–3.78 | 3.95–3.92 | ||
| β-GlcNIII-(1→ | 4.94 [d, 8.4 Hz] | 3.29–3.22 | 3.74–3.68 | 3.61–3.55 | 3.58–3.53 | 3.95–3.85 | 3.95–3.85 | ||
| α-l-Rha-(1→ | 4.80–4.77 | 4.03–4.00 | 3.81–3.79 | 3.46–3.42 | 3.77–3.72 | 1.31 [d, 6.1 Hz] | |||
| Residue | H3eq | H3ax | H4 | H5 | H6 | H7 | H8a | H8b | |
| →4,5)-α-KdoI-(2→ | 2.30–2.23 | 2.04 [dd, 12.3, 12.0 Hz] | 4.17–4.12 | 4.31–4.27 | 3.74–3.71 | 3.89–3.85 | 3.63–3.58 | 3.93–3.88 | |
| α-KdoII-(2→ | 2.11 [dd, 12.9, 4.3 Hz] | 1.85 [dd, 12.9, 12.6 Hz] | 4.18–4.13 | 4.09–4.07 | 3.68–3.65 | 4.04–3.99 | 3.71–3.66 | 3.99–3.94 |
| Residue | C1 | C2 | C3 | C4 | C5 | C6 | C7 | C8 |
|---|---|---|---|---|---|---|---|---|
| →6)-α-GlcNI-(1→P | 92.4–92.2 | 54.6 | 69.9 | 70.1 | 73.3 | 69.9 | ||
| →6)-β-GlcNII4P-(1→ | 99.6 | 56.1 | 72.1 | 75.0 | 74.4 | 63.1 | ||
| →4,5)-α-KdoI-(2→ | n.d. | n.d. | 34.8 | 71.6 | 68.8 | 72.9 | 69.7 | 64.3 |
| α-KdoII-(2→ | n.d. | n.d. | 35.5 | 66.1 | 67.1 | 73.1 | 71.2 | 63.7 |
| →3)-α-HepI2P4P-(1→ | 98.0 | 75.0 | 74.8 | 71.1 | 72.6 | 69.7 | 63.7 | |
| →3)-α-HepII6P-(1→ | 102.8 | 69.7 | 78.3 | 66.1 | 72.3 | 74.0 | 62.0 | |
| →3,4)-α-GalN-(1→ | 97.2 | 51.1 | 78.4 | 75.9 | 73.0 | 60.1 | ||
| →2)-β-GlcI-(1→ | 104.0 | 84.0 | 76.0 | 70.6 | 76.4 | 61.3 | ||
| →6)-α-GlcII-(1→ | 100.0 | 72.3 | 73.3 | 69.1 | 71.1 | 66.9 | ||
| β-GlcNIII-(1→ | 101.8 | 56.7 | 72.4 | 69.6 | 76.6 | 60.2 | ||
| α-l-Rha-(1→ | 101.9 | 70.6 | 70.8 | 72.7 | 69.4 | 17.7 |
| Residue | 31P Chemical Shift [ppm] |
|---|---|
| →6)-α-GlcNI-(1→P | -1.44 |
| →6)-β-GlcNII4P-(1→ | 0.48 |
| →3)-α-HepI2P4P-(1→ | 0.28 |
| →3)-α-HepI2P4P-(1→ | 1.57 |
| →3)-α-HepII6P-(1→ | 1.70 |
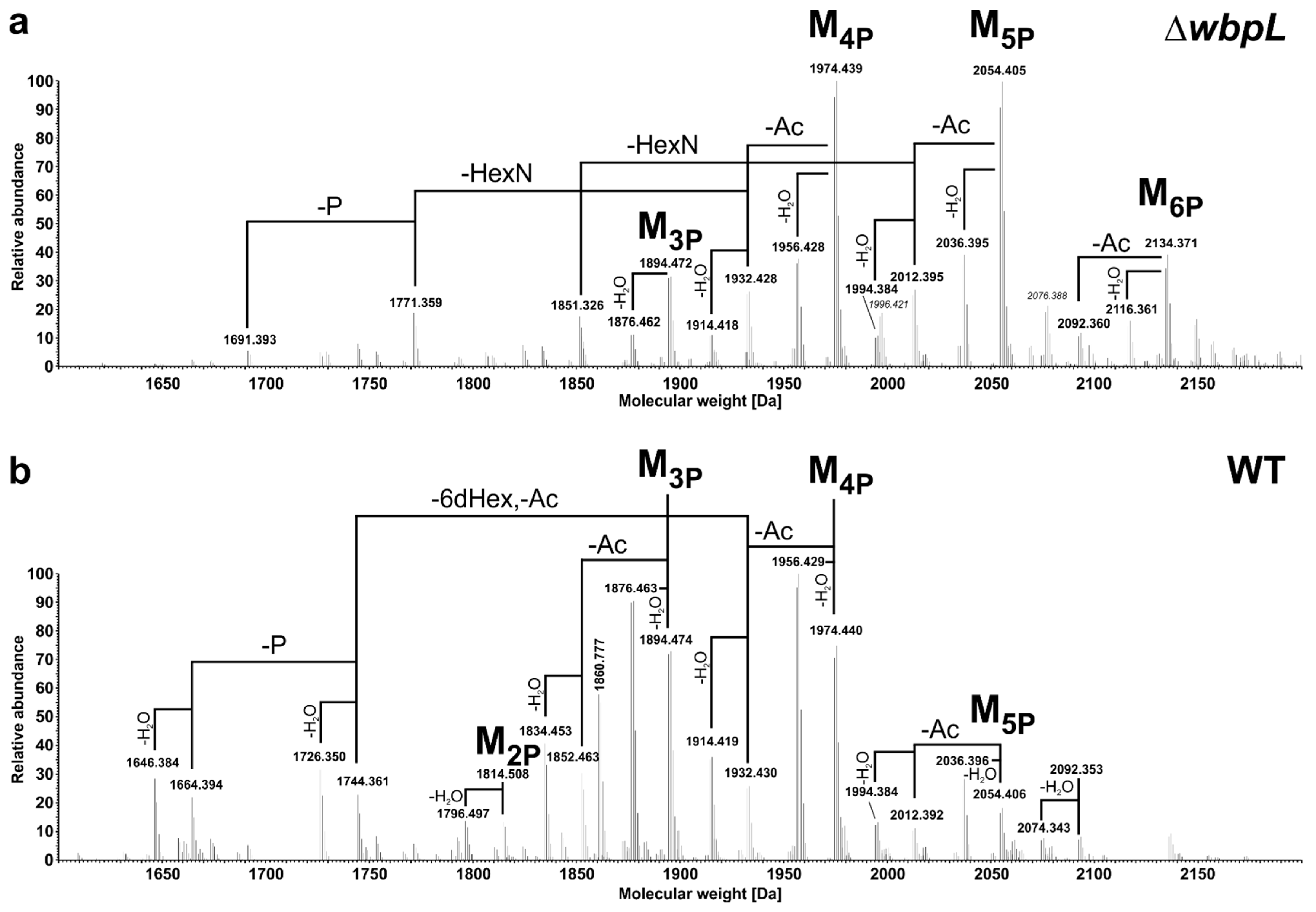
| Molecule | Pst DC3000 ΔwbpL | Pst DC3000 WT | |||
|---|---|---|---|---|---|
| Calculated Mass [Da] | Observed Mass [Da] | Error [Δppm] | Observed Mass [Da] | Error [Δppm] | |
| Kdo1Hep1HepCm1Hex2HexN2Ala1Ac1P3–H2O | 1646.381 | 1646.383 * | 1.2 | 1646.384 | 1.8 |
| Kdo1Hep1HepCm1Hex2HexN2Ala1Ac1P3 | 1664.390 | 1664.394 * | 2.4 | 1664.394 | 2.4 |
| Kdo1Hep1HepCm1Hex26dHex1HexN1Ala1Ac2P3–H2O | 1673.380 | 1673.383 * | 1.8 | 1673.384 | 2.4 |
| Kdo1Hep1HepCm1Hex26dHex1HexN1Ala1Ac2P3 | 1691.391 | 1691.393 | 1.2 | 1691.395 | 2.4 |
| Kdo1Hep1HepCm1Hex2HexN2Ala1Ac1P4–H2O | 1726.347 | 1726.349 | 1.2 | 1726.350 | 1.7 |
| Kdo1Hep1HepCm1Hex26dHex1HexN1Ala1Ac1P4 | 1729.347 | 1729.348 | 0.6 | n.d. | - |
| Kdo1Hep1HepCm1Hex2HexN2Ala1Ac1P4 | 1744.358 | 1744.360 | 1.1 | 1744.361 | 1.7 |
| Kdo1Hep1HepCm1Hex26dHex1HexN1Ala1Ac2P4 | 1771.357 | 1771.359 | 1.1 | 1771.361 | 2.3 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P2–H2O | 1796.494 | n.d. | - | 1796.497 | 1.7 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P2 (M2P) | 1814.504 | 1814.509 * | 2.8 | 1814.508 | 2.2 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac2P3–H2O | 1834.449 | 1834.452 * | 1.6 | 1834.453 | 2.2 |
| Kdo1Hep1HepCm1Hex26dHex1HexN1Ala1Ac2P5 | 1851.324 | 1851.326 | 1.1 | n.d. | - |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac2P3 | 1852.460 | 1852.462 | 1.1 | 1852.463 | 1.6 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac3P3–H2O | 1876.460 | 1876.462 | 1.1 | 1876.463 | 1.6 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P3 (M3P) | 1894.470 | 1894.472 | 1.1 | 1894.474 | 2.1 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac2P4–H2O | 1914.416 | 1914.418 | 1.0 | 1914.419 | 1.6 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac2P4 | 1932.426 | 1932.428 | 1.0 | 1932.430 | 2.1 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac3P4–H2O | 1956.426 | 1956.428 | 1.0 | 1956.429 | 1.5 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P4 (M4P) | 1974.437 | 1974.439 | 1.0 | 1974.440 | 1.5 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac2P5–H2O | 1994.382 | 1994.384 | 1.0 | 1994.384 | 1.0 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac2P5 | 2012.393 | 2012.395 | 1.0 | 2012.392 | −0.5 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac3P5–H2O | 2036.393 | 2036.395 | 1.0 | 2036.396 | 1.5 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P5 (M5P) | 2054.403 | 2054.405 | 1.0 | 2054.406 | 1.5 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac2P6–H2O | 2074.348 | n.d. | - | 2074.343 | −2.4 |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac2P6 | 2092.359 | 2092.360 | 0.5 | 2092.353 | −3.3 |
| Kdo1Hep1HepCm1Hex26dHex1 HexN2Ala1Ac3P6–H2O | 2116.359 | 2116.361 | 0.9 | n.d. | - |
| Kdo1Hep1HepCm1Hex26dHex1HexN2Ala1Ac3P6 (M6P) | 2134.369 | 2134.371 | 0.9 | n.d. | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutschera, A.; Schombel, U.; Schwudke, D.; Ranf, S.; Gisch, N. Analysis of the Structure and Biosynthesis of the Lipopolysaccharide Core Oligosaccharide of Pseudomonas syringae pv. tomato DC3000. Int. J. Mol. Sci. 2021, 22, 3250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063250
Kutschera A, Schombel U, Schwudke D, Ranf S, Gisch N. Analysis of the Structure and Biosynthesis of the Lipopolysaccharide Core Oligosaccharide of Pseudomonas syringae pv. tomato DC3000. International Journal of Molecular Sciences. 2021; 22(6):3250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063250
Chicago/Turabian StyleKutschera, Alexander, Ursula Schombel, Dominik Schwudke, Stefanie Ranf, and Nicolas Gisch. 2021. "Analysis of the Structure and Biosynthesis of the Lipopolysaccharide Core Oligosaccharide of Pseudomonas syringae pv. tomato DC3000" International Journal of Molecular Sciences 22, no. 6: 3250. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22063250