
Cytokinin Type-B Response Regulators Promote Bulbil Initiation in Lilium lancifolium

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Full-Length Cloning and Sequence Analysis of Type-B ARRs
2.2. Subcellular Localization of Type-B ARRs
2.3. Expression Pattern of Five LlRRs
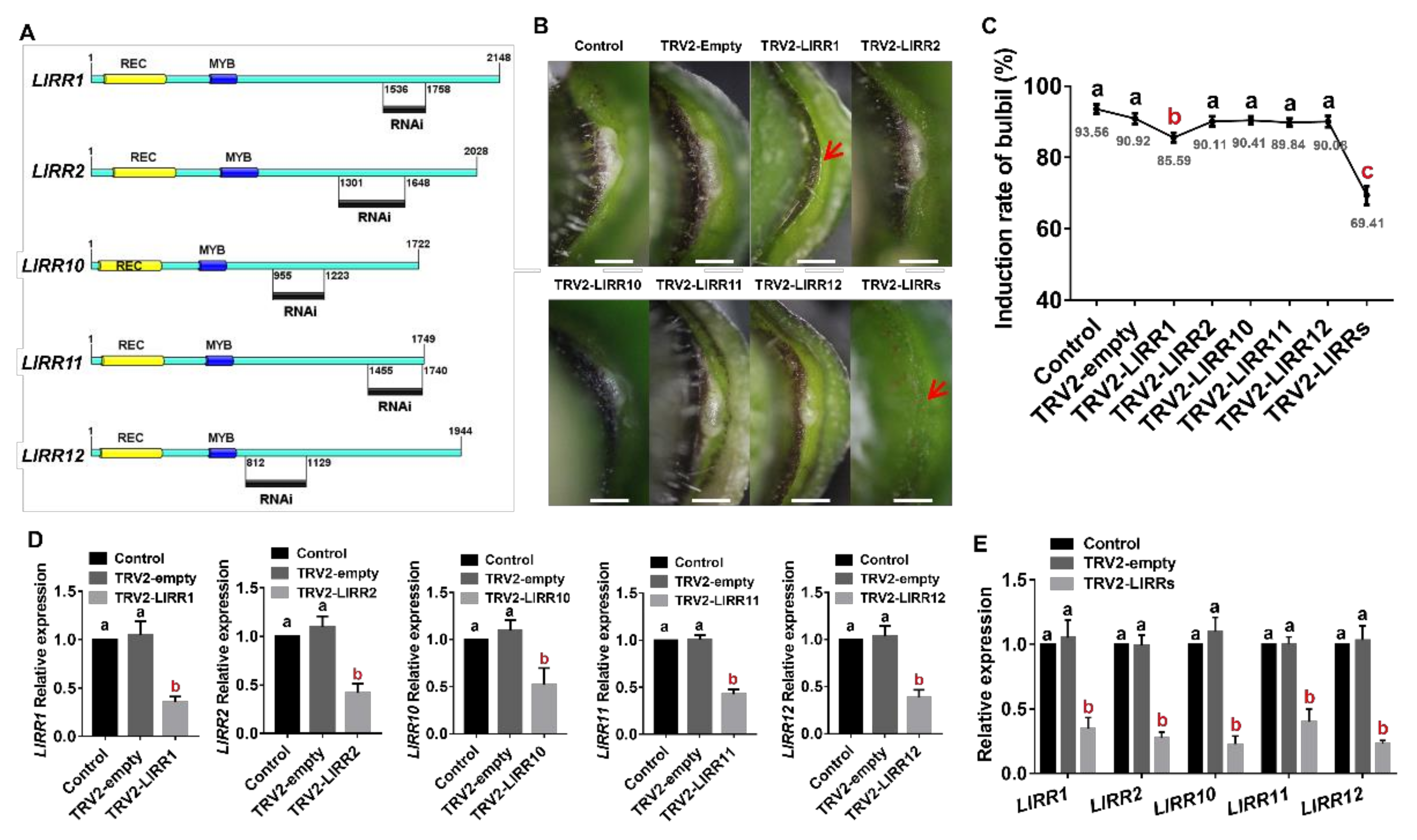
2.4. Type-B LlRRs as Positive Regulators of Bulbil Formation
2.5. A Cytokinin Feedback Loop Involved in the Initiation of the AM
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Isolation of Type-B LlRRs and the LlRR9 Promoter
4.3. Real-Time RT-PCR (qRT-PCR)
4.4. Subcellular Localization
4.5. Virus-Induced Gene Silencing (VIGS)
4.6. Dual-Luciferase Reporter Assay
4.7. Electrophoretic Mobility Shift Assay (EMSA)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- China Pharmacopoeia Committee. Chinese Pharmacopoeia; Chemical Industry Press: Beijing, China, 2005.
- Yu, X.R.; Zhang, J.; Shao, S.S.; Yu, H.; Xiong, F.; Wang, Z. Morphological and physicochemical properties of bulb and bulbil starches from Lilium lancifolium. Starch-Stärke 2015, 67, 448–458. [Google Scholar] [CrossRef]
- Bach, A.; Sochacki, D. Propagation of ornamental geophytes: Physiology and management. In Ornamental Geophytes: From Basic Science to Sustainable Production; Kamenetsky, R., Okubo, H., Eds.; CRC Press: Boca Raton, FL, USA, 2012; pp. 261–286. [Google Scholar]
- Noda, S. Cytogenetic behavior, chromosomal differentiations, and geographic distribution in Lilium lancifolium (Liliaceae). Plant Species Biol. 1986, 1, 69–78. [Google Scholar] [CrossRef]
- Kim, J.H.; Kyung, H.Y.; Choi, Y.S.; Lee, J.K.; Hiramatsu, M.; Okubo, H. Geographic distribution and habitat differentiation in diploid and triploid Lilium lancifolium of South Korea. J. Fac. Agric. 2006, 51, 239–243. [Google Scholar] [CrossRef]
- Yang, P.P.; Xu, L.F.; Xu, H.; Tang, Y.C.; He, G.R.; Cao, Y.W.; Feng, Y.Y.; Yuan, S.X.; Ming, J. Histological and transcriptomic analysis during bulbil formation in Lilium lancifolium. Front. Plant Sci. 2017, 8, 1508. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.N.; Möller, M.; Cronk, Q.C. Altered expression of GFLO, the Gesneriaceae homologue of FLORICAULA/LEAFY, is associated with the transition to bulbil formation in Titanotrichum oldhamii. Dev. Genes Evol. 2004, 214, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.D.; Bryan, A. Plant Form: An Illustrated Guide to Flowering Plant Morphology; Timber Press: Portland, OR, USA, 2008. [Google Scholar]
- Abraham-Juárez, M.J.; Martínez-Hernández, A.; Leyva-González, M.A.; Herrera-Estrella, L.; Simpson, J. Class I KNOX genes are associated with organogenesis during bulbil formation in Agave tequilana. J. Exp. Bot. 2010, 61, 4055–4067. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, S.; Juárez, M.J.; Simpson, J. Agave tequilana MADS genes show novel expression patterns in meristems, developing bulbils and floral organs. Sex. Plant Reprod. 2012, 25, 11–26. [Google Scholar] [CrossRef]
- Yang, P.P.; Xu, L.F.; Xu, H.; He, G.R.; Feng, Y.Y.; Cao, Y.W.; Tang, Y.C.; Yuan, S.X.; Ming, J. Morphological and anatomical observation during the formation of bulbils in Lilium lancifolium. Caryologia 2018, 71, 146–149. [Google Scholar] [CrossRef]
- He, G.R.; Yang, P.P.; Tang, Y.C.; Cao, Y.W.; Qi, X.Y.; Xu, L.F.; Ming, J. Mechanism of exogenous cytokinins inducing bulbil formation in Lilium lancifolium In Vitro. Plant Cell Rep. 2020, 39, 861–872. [Google Scholar] [CrossRef]
- Wang, C.N.; Cronk, Q.C. Meristem fate and bulbil formation in Titanotrichum (Gesneriaceae). Am. J. Bot. 2003, 90, 1696–1707. [Google Scholar] [CrossRef]
- Abraham-Juárez, M.J.; Hernández Cárdenas, R.; Santoyo Villa, J.N.; O’Connor, D.; Sluis, A.; Hake, S.; Ordaz-Ortiz, J.; Terry, L.; Simpson, J. Functionally different PIN proteins control auxin flux during bulbil development in Agave tequilana. J. Exp. Bot. 2015, 66, 3893–3905. [Google Scholar] [CrossRef] [PubMed]
- PENG, X.Y.; ZHOU, P.H.; ZHANG, L.B.; JIANG, D.S.; LIU, Y.F. The Induction of Bulbils of Dioscorea zingiberensis. J. Trop. Subtrop. Bot. 2005, 4, 319–323. [Google Scholar]
- Navarro, C.; Cruz-Oró, E.; Prat, S. Conserved function of FLOWERING LOCUS T (FT) homologues as signals for storage organ differentiation. Curr. Opin. Plant Biol. 2015, 23, 45–53. [Google Scholar] [CrossRef]
- Long, J.; Barton, M.K. Initiation of axillary and floral meristems in Arabidopsis. Dev. Biol. 2000, 218, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, J.; Shi, B.H.; Yu, T.; Qi, J.Y.; Meyerowitz, E.M.; Jiao, Y.L. The stem cell niche in leaf axils is established by auxin and cytokinin in Arabidopsis. Plant Cell 2014, 26, 2055–2067. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Tian, C.H.; Zhang, C.; Shi, B.H.; Cao, X.W.; Zhang, T.Q.; Zhao, Z.; Wang, J.W.; Jiao, Y.L. Cytokinin signaling activates WUSCHEL expression during axillary meristem initiation. Plant Cell 2017, 29, 1373–1387. [Google Scholar] [CrossRef]
- Mok, D.W.; Mok, M.C. Cytokinin metabolism and action. Annu. Rev. Plant Biol. 2001, 52, 89–118. [Google Scholar] [CrossRef]
- Davies, P.J. Plant Hormones: Biosynthesis, Signal Transduction, Action! Springer Science & Business Media: Berlin/Heidelberg, Germany, 2004. [Google Scholar]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [CrossRef]
- Hwang, I.; Sheen, J.; Müller, B. Cytokinin signaling networks. Annu. Rev. Plant Biol. 2012, 63, 353–380. [Google Scholar] [CrossRef]
- O’Brien, J.A.; Benková, E. Cytokinin cross-talking during biotic and abiotic stress responses. Front. Plant Sci. 2013, 4, 451. [Google Scholar] [CrossRef]
- Kieber, J.J.; Schaller, G.E. Cytokinins. Arab. Book 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, T. Compilation of all genes encoding two-component phosphotransfer signal transducers in the genome of Escherichia coli. DNA Res. 1997, 4, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Stock, A.M.; Robinson, V.L.; Goudreau, P.N. Two-component signal transduction. Annu. Rev. Biochem. 2000, 69, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Kieber, J.J.; Shiu, S.H. Two-component signaling elements and histidyl-aspartyl phosphorelays. Arab. Book 2008, 6, e0112. [Google Scholar] [CrossRef]
- Brandstatter, I.; Kieber, J.J. Two genes with similarity to bacterial response regulators are rapidly and specifically induced by cytokinin in Arabidopsis. Plant Cell 1998, 10, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- D’Agostino, I.B.; Deruere, J.; Kieber, J.J. Characterization of the response of the Arabidopsis response regulator gene family to cytokinin. Plant Physiol. 2000, 124, 1706–1717. [Google Scholar] [CrossRef] [PubMed]
- Kiba, T.; Yamada, H.; Sato, S.; Kato, T.; Tabata, S.; Yamashino, T.; Mizuno, T. The type-A response regulator, ARR15, acts as a negative regulator in the cytokinin-mediated signal transduction in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 868–874. [Google Scholar] [CrossRef]
- To, J.P.; Haberer, G.; Ferreira, F.J.; Deruère, J.; Mason, M.G.; Schaller, G.E.; Alonso, J.M.; Ecker, J.R.; Kieber, J.J. Type-A ARRs are partially redundant negative regulators of cytokinin signaling. Plant Cell 2004, 16, 658–671. [Google Scholar] [CrossRef]
- Kiba, T.; Aoki, K.; Sakakibara, H.; Adn Mizuno, T. Arabidopsis response regulator, ARR22, ectopic expression of which results in phenotypes similar to the wol cytokinin-receptor mutant. Plant Cell Physiol. 2004, 45, 1063–1077. [Google Scholar] [CrossRef]
- Hosoda, K.; Imamura, A.; Katoh, E.; Hatta, T.; Tachiki, M.; Yamada, H.; Mizuno, T.; Yamazaki, T. Molecular structure of the GARP family of plant Myb-related DNA binding motifs of the Arabidopsis response regulators. Plant Cell 2002, 14, 2015–2029. [Google Scholar] [CrossRef]
- Sakakibara, H.; Suzuki, M.; Takei, K.; Deji, A.; Taniguchi, M.; Sugiyama, T. A response-regulator homologue possibly involved in nitrogen signal transduction mediated by cytokinin in maize. Plant J. 1998, 14, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Mason, M.G.; Li, J.; Mathews, D.E.; Kieber, J.J.; Schaller, G.E. Type-B response regulators display overlapping expression patterns in Arabidopsis. Plant Physiol. 2004, 135, 927–937. [Google Scholar] [CrossRef] [PubMed]
- Schaller, G.E.; Doi, K.; Hwang, I.; Kieber, J.J.; Khurana, J.P.; Kurata, N.; Mizuno, T.; Pareek, A.; Shiu, S.H.; Wu, P.; et al. Nomenclature for two-component signaling elements of rice. Plant Physiol. 2007, 143, 555–557. [Google Scholar] [CrossRef]
- Tsai, Y.C.; Weir, N.R.; Hill, K.; Zhang, W.; Kim, H.J.; Shiu, S.H.; Schaller, G.E.; Kieber, J.J. Characterization of genes involved in cytokinin signaling and metabolism from rice. Plant Physiol. 2012, 158, 1666–1684. [Google Scholar] [CrossRef]
- Sakai, H.; Aoyama, T.; Oka, A. Arabidopsis ARR1 and ARR2 response regulators operate as transcriptional activators. Plant J. 2000, 24, 703–711. [Google Scholar] [CrossRef]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Sakai, H.; Honma, T.; Aoyama, T.; Sato, S.; Kato, T.; Tabata, S.; Oka, A. ARR1, a transcription factor for genes immediately responsive to cytokinins. Science 2001, 294, 1519–1521. [Google Scholar] [CrossRef]
- Mason, M.G.; Mathews, D.E.; Argyros, D.A.; Maxwell, B.B.; Kieber, J.J.; Alonso, J.M.; Ecker, J.R.; Schaller, G.E. Multiple type-B response regulators mediate cytokinin signal transduction in Arabidopsis. Plant Cell 2005, 17, 3007–3018. [Google Scholar] [CrossRef] [PubMed]
- Yokoyama, A.; Yamashino, T.; Amano, Y.I.; Tajima, Y.; Imamura, A.; Sakakibara, H.; Mizuno, T. Type-B ARR transcription factors, ARR10 and ARR12, are implicated in cytokinin-mediated regulation of protoxylem differentiation in roots of Arabidopsis thaliana. Plant Cell Physiol. 2007, 48, 84–96. [Google Scholar] [CrossRef]
- Argyros, R.D.; Mathews, D.E.; Chiang, Y.H.; Palmer, C.M.; Thibault, D.M.; Etheridge, N.; Argyros, D.A.; Mason, M.G.; Kieber, J.J.; Schaller, G.E. Type B response regulators of Arabidopsis play key roles in cytokinin signaling and plant development. Plant Cell 2008, 20, 2102–2116. [Google Scholar] [CrossRef]
- Ishida, K.; Yamashino, T.; Yokoyama, A.; Mizuno, T. Three type-B response regulators, ARR1, ARR10 and ARR12, play essential but redundant roles in cytokinin signal transduction throughout the life cycle of Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Dello Ioio, R.; Galinha, C.; Fletcher, A.G.; Grigg, S.P.; Molnar, A.; Willemsen, V.; Scheres, B.; Sabatini, S.; Baulcombe, D.; Maini, P.K.; et al. A PHABULOSA/cytokinin feedback loop controls root growth in Arabidopsis. Curr. Biol. 2012, 22, 1699–1704. [Google Scholar] [CrossRef] [PubMed]
- Meng, W.J.; Cheng, Z.J.; Sang, Y.L.; Zhang, M.M.; Rong, X.F.; Wang, Z.W.; Tang, Y.Y.; Zhang, X.S. Type-B Arabidopsis response regulators specify the shoot stem cell niche by dual regulation of WUSCHEL. Plant Cell 2017, 29, 1357–1372. [Google Scholar] [CrossRef] [PubMed]
- Olivia, G.; Audrey, O.; Grégory, G.; Mouadh, C.; Vincent, C.; Gaëlle, G.; Nicolas, P.; Joel, C.; Martine, C. A type-B response regulator drives the expression of the hydroxymethylbutenyl diphosphate synthase gene in periwinkle. J. Plant Physiol. 2012, 169, 1571–1574. [Google Scholar] [CrossRef]
- Ramiírez-Carvajal, G.A.; Morse, A.M.; Dervinis, C.; Davis, J.M. The Cytokinin Type-B Response Regulator PtRR13 Is a Negative Regulator of Adventitious Root Development in Populus. Plant Physiol. 2009, 150, 759–771. [Google Scholar] [CrossRef]
- Aki, S.S.; Mikami, T.; Naramoto, S.; Nishihama, R.; Ishizaki, K.; Kojima, M.; Takebayashi, Y.; Sakakibara, H.; Kyozuka, J.; Kohchi, T.; et al. Cytokinin Signaling Is Essential for Organ Formation in Marchantia polymorpha. Plant Cell Physiol. 2019, 60, 1842–1854. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.H.; Ha, C.V.; Nishiyama, R.; Watanabe, Y.; Leyva-González, M.A.; Fujita, Y.; Tran, U.T.; Li, W.Q.; Tanaka, M.; Seki, M.; et al. Arabidopsis type B cytokinin response regulators ARR1, ARR10, and ARR12 negatively regulate plant responses to drought. Proc. Natl. Acad. Sci. USA 2016, 113, 3090–3095. [Google Scholar] [CrossRef]
- Kim, H.J.; Ryu, H.; Hong, S.H.; Woo, H.R.; Lim, P.O.; Lee, L.C.; Sheen, J.; Nam, H.G.; Hwang, I. Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis. Proc. Natl. Acad. Sci. USA 2006, 103, 814–819. [Google Scholar] [CrossRef]
- Dai, X.H.; Liu, Z.H.; Qiao, M.; Li, J.; Li, S.; Xiang, F.N. ARR12 promotes de novo shoot regeneration in Arabidopsis thaliana via activation of WUSCHEL expression. J. Integr. Plant Biol. 2017, 59, 747–758. [Google Scholar] [CrossRef]
- Liu, Z.H.; Dai, X.H.; Li, J.; Liu, N.; Liu, X.Z.; Li, S.; Xiang, F.N. The Type-B Cytokinin Response Regulator ARR1 Inhibits Shoot Regeneration in an ARR12-Dependent Manner in Arabidopsis. Plant Cell 2020, 32, 2271–2291. [Google Scholar] [CrossRef]
- Yan, Z.W.; Liu, X.; Ljung, K.; Li, S.N.; Zhao, W.Y.; Yang, F.; Wang, M.L.; Tao, Y. Type B response regulators act as central integrators in transcriptional control of the auxin biosynthesis enzyme TAA1. Plant Physiol. 2017, 175, 1438–1454. [Google Scholar] [CrossRef]
- Taniguchi, M.; Sasaki, N.; Tsuge, T.; Aoyama, T.; Oka, A. ARR1 directly activates cytokinin response genes that encode proteins with diverse regulatory functions. Plant Cell Physiol. 2007, 48, 263–277. [Google Scholar] [CrossRef]
- Leibfried, A.; To, J.P.; Busch, W.; Stehling, S.; Kehle, A.; Demar, M.; Kieber, J.J.; Lohmann, J.U. WUSCHEL controls meristem size by direct transcriptional regulation of cytokinin inducible response regulators. Nature 2005, 438, 1172–1175. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.J.; Park, J.Y.; Ku, S.J.; Ha, Y.M.; Kim, S.; Kim, M.D.; Oh, M.; Kim, J. Genome-wide expression profiling of Arabidopsis response regulator 7 (ARR7) overexpression in cytokinin response. Mol. Genet. Genom. 2007, 277, 115–137. [Google Scholar] [CrossRef] [PubMed]
- To, J.P.; Deruère, J.; Maxwell, B.B.; Morris, V.F.; Hutchison, C.E.; Schaller, G.E.; Kieber, J.J. Cytokinin regulates type-A Arabidopsis response regulator activity and protein stability via two- component phosphorelay. Plant Cell 2007, 19, 3901–3914. [Google Scholar] [CrossRef]
- Ramireddy, E.; Brenner, W.G.; Pfeifer, A.; Heyl, A.; Schmülling, T. In planta analysis of a cis-regulatory cytokinin response motif in Arabidopsis and identification of a novel enhancer sequence. Plant Cell Physiol. 2013, 54, 1079–1092. [Google Scholar] [CrossRef]
- Gordon, S.P.; Chickarmane, V.S.; Ohno, C.; Meyerowitz, E.M. Multiple feedback loops through cytokinin signaling control stem cell number within the Arabidopsis shoot meristem. Proc. Natl. Acad. Sci. USA 2009, 106, 16529–16534. [Google Scholar] [CrossRef]
- Liu, Z.H.; Li, J.; Wang, L.; Li, Q.; Lu, Q.; Yu, Y.C.; Li, S.; Bai, M.Y.; Hu, Y.X.; Xiang, F.N. Repression of callus initiation by the miRNA-directed interaction of auxin–cytokinin in Arabidopsis thaliana. Plant J. 2016, 87, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. 20 years of the SMART protein domain annotation resource. Nucleic Acids Res. 2018, 46, D493–D496. [Google Scholar] [CrossRef]
- Kenichi, H.; Yoshihiro, U.; Masao, I.; Tomoko, K. Plant cis-acting regulatory DNA elements (PLACE) database. Nucleic Acids Res. 1999, 27, 297–300. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for In Silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.F.; Xu, H.; Cao, Y.W.; Yang, P.P.; Feng, Y.Y.; Tang, Y.C.; Yuan, S.X.; Ming, J. Validation of reference genes for quantitative real-time PCR during bicolor tepal development in asiatic hybrid lilies (Lilium spp.). Front. Plant Sci. 2017, 8, 669. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Wang, S.H.; Yang, X.Y.; Xu, M.N.; Lin, X.Z.; Lin, T.; Qi, J.J.; Shao, G.J.; Tian, N.N.; Yang, Q.; Zhang, Z.H.; et al. A rare SNP identified a TCP transcription factor essential for tendril development in cucumber. Mol. Plant 2015, 8, 1795–1808. [Google Scholar] [CrossRef]
- Tian, J.; Pei, H.X.; Zhang, S.; Chen, J.W.; Chen, W.; Yang, R.Y.; Meng, Y.L.; You, J.; Gao, J.P.; Ma, N. TRV–GFP: A modified Tobacco rattle virus vector for efficient and visualizable analysis of gene function. J. Exp. Bot. 2014, 65, 311–322. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Wang, X.C.; Hong, S.L.; Li, Y.S.; Zuo, J.R. Deletion of the initial 45 residues of ARR18 induces cytokinin response in Arabidopsis. J. Genet. Genom. 2012, 39, 37–46. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, G.; Yang, P.; Cao, Y.; Tang, Y.; Wang, L.; Song, M.; Wang, J.; Xu, L.; Ming, J. Cytokinin Type-B Response Regulators Promote Bulbil Initiation in Lilium lancifolium. Int. J. Mol. Sci. 2021, 22, 3320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073320
He G, Yang P, Cao Y, Tang Y, Wang L, Song M, Wang J, Xu L, Ming J. Cytokinin Type-B Response Regulators Promote Bulbil Initiation in Lilium lancifolium. International Journal of Molecular Sciences. 2021; 22(7):3320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073320
Chicago/Turabian StyleHe, Guoren, Panpan Yang, Yuwei Cao, Yuchao Tang, Ling Wang, Meng Song, Jing Wang, Leifeng Xu, and Jun Ming. 2021. "Cytokinin Type-B Response Regulators Promote Bulbil Initiation in Lilium lancifolium" International Journal of Molecular Sciences 22, no. 7: 3320. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073320