
Biochemical Characterization of Recombinant UDPG-Dependent IAA Glucosyltransferase from Maize (Zea mays)


Abstract
:1. Introduction
2. Results and Discussion
2.1. Production of Recombinant Maize IAGlc Synthase in Escherichia coli and Purification of the Enzyme
2.2. Kinetic Properties of Maize Recombinant IAGlc Synthase
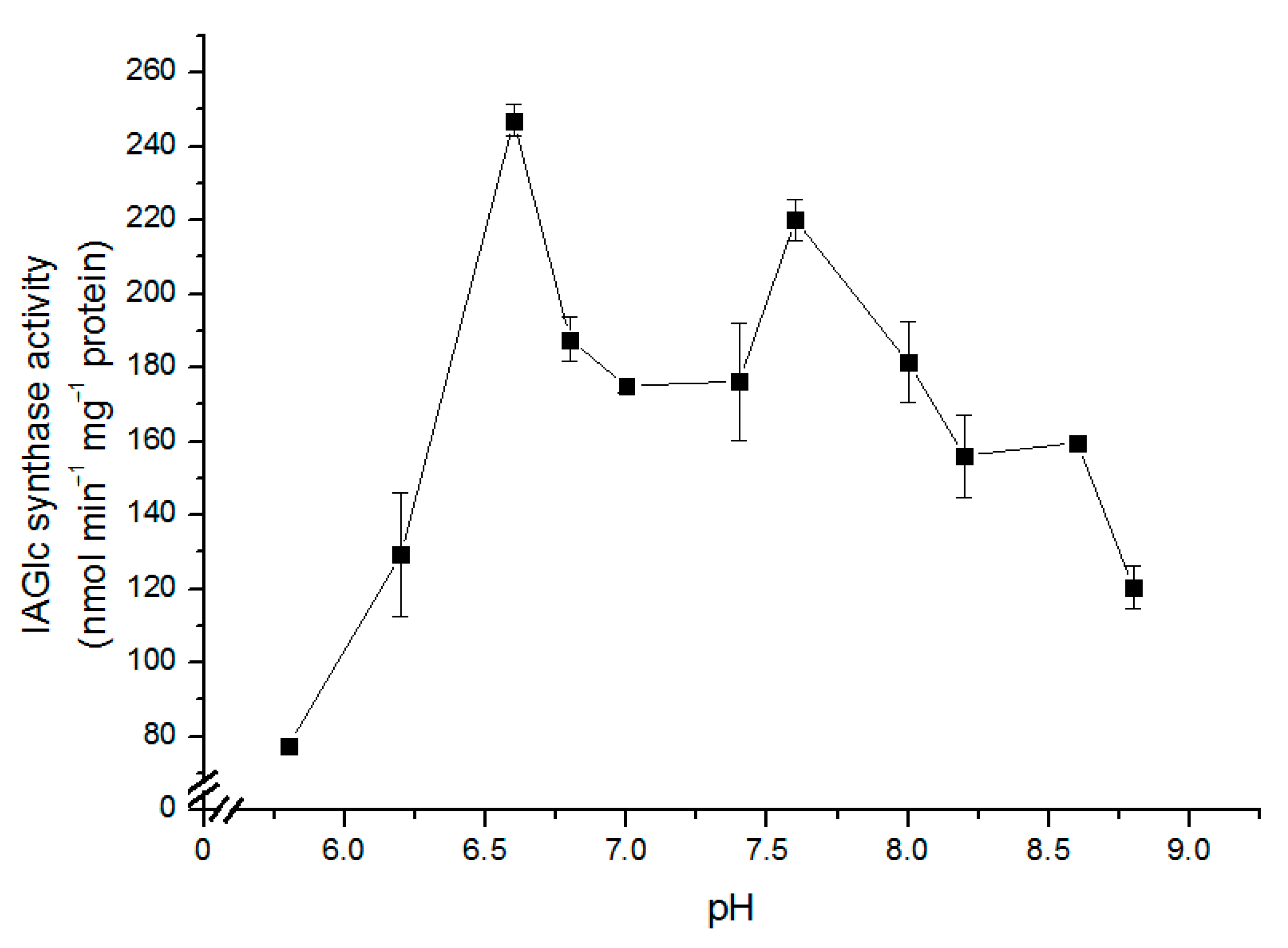
2.2.1. Determination of pH Effect on IAGlc Synthase
2.2.2. Determination of IAGlc Synthase Substrate Specificity
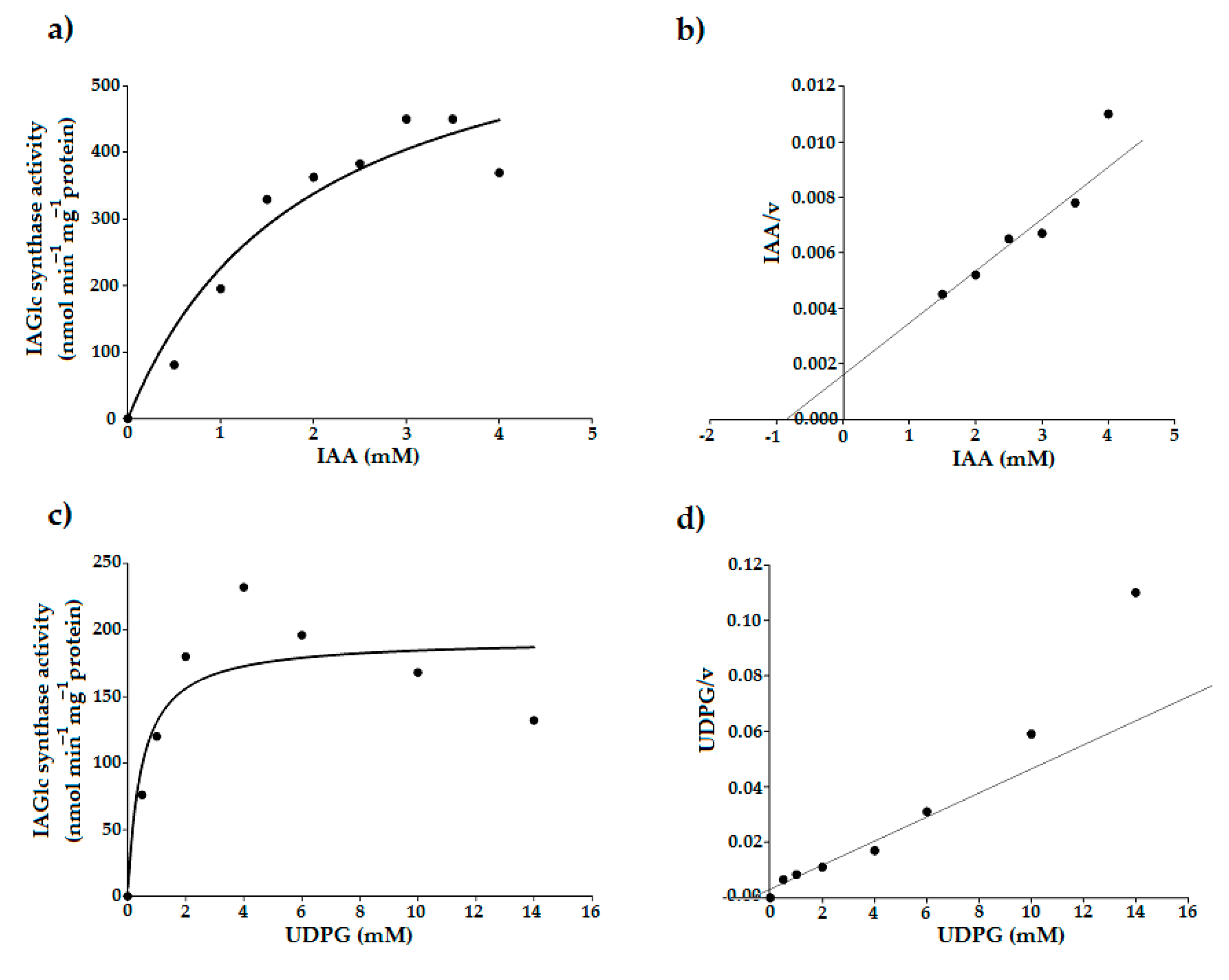
2.2.3. Determination of Kinetic Parameters of IAGlc Synthase
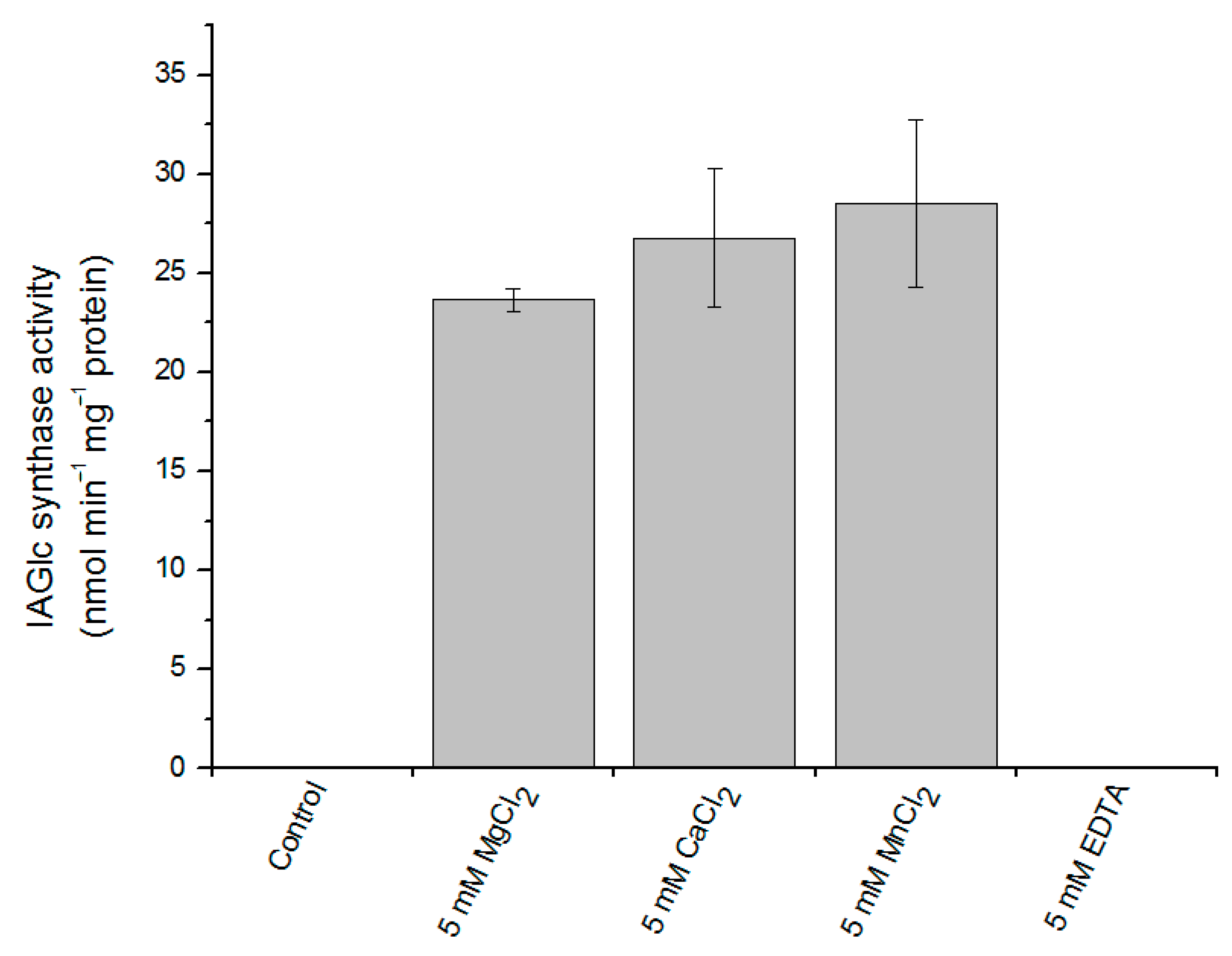
2.2.4. Effect of Divalent Cations on IAGlc Synthase
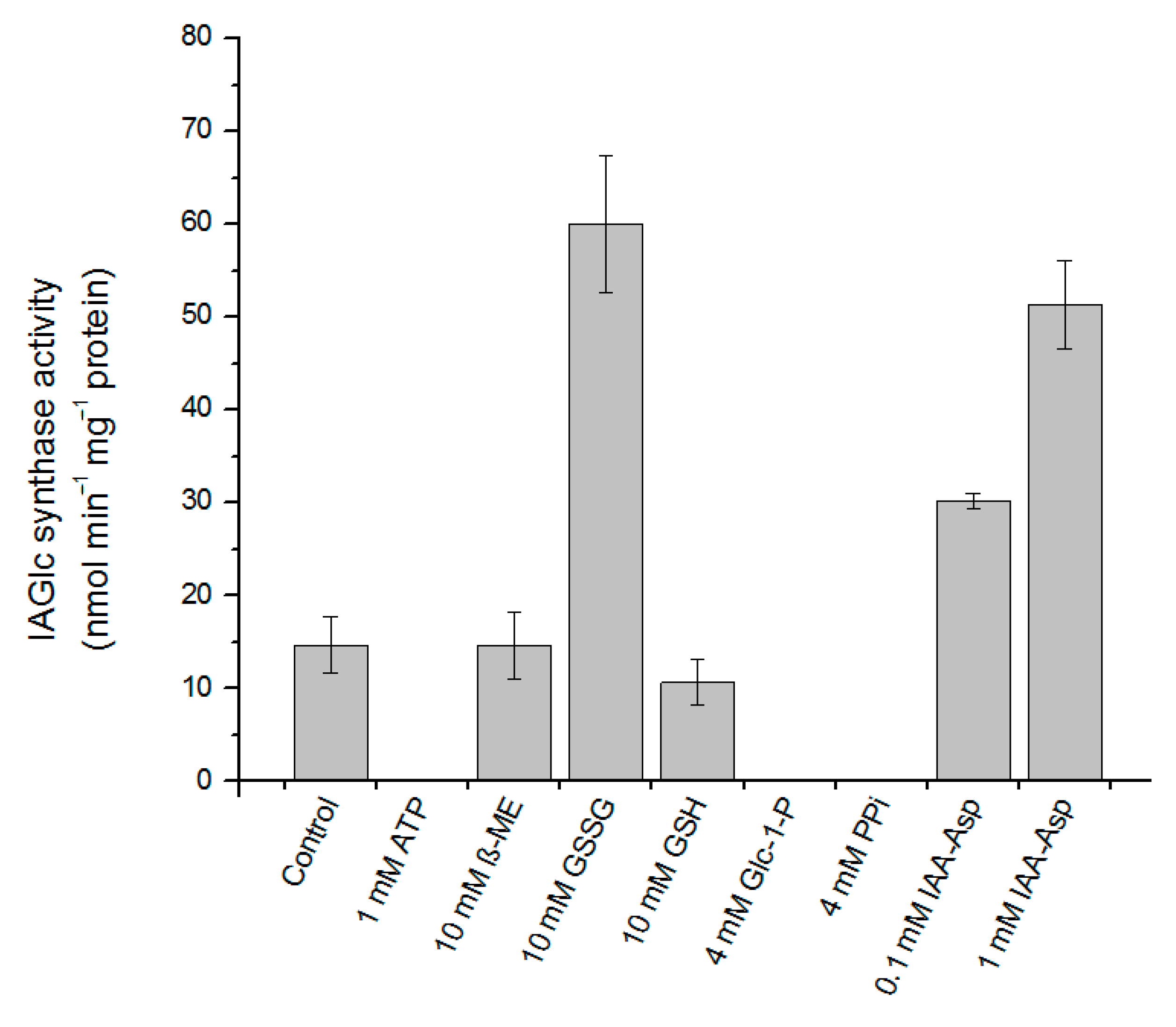
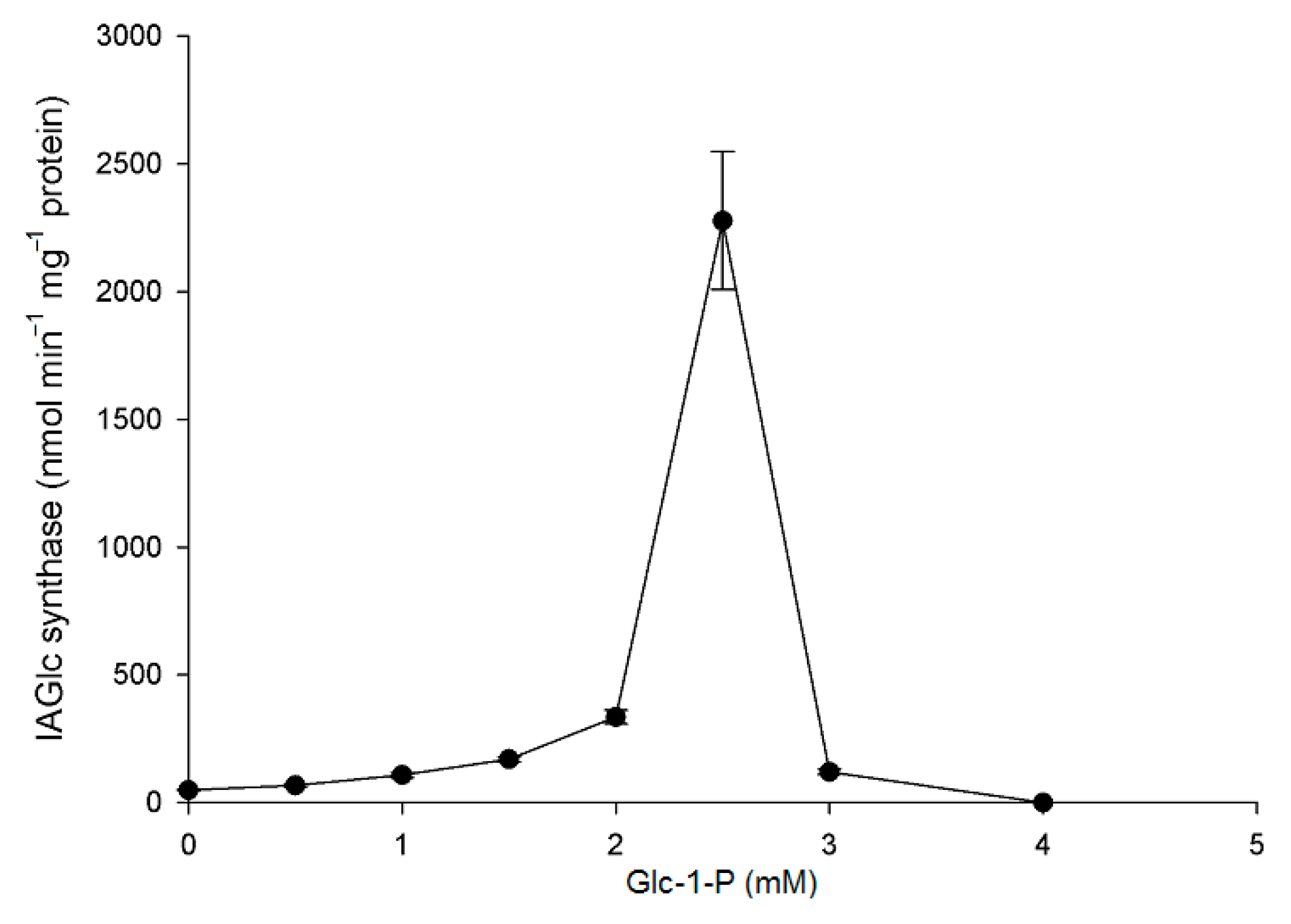
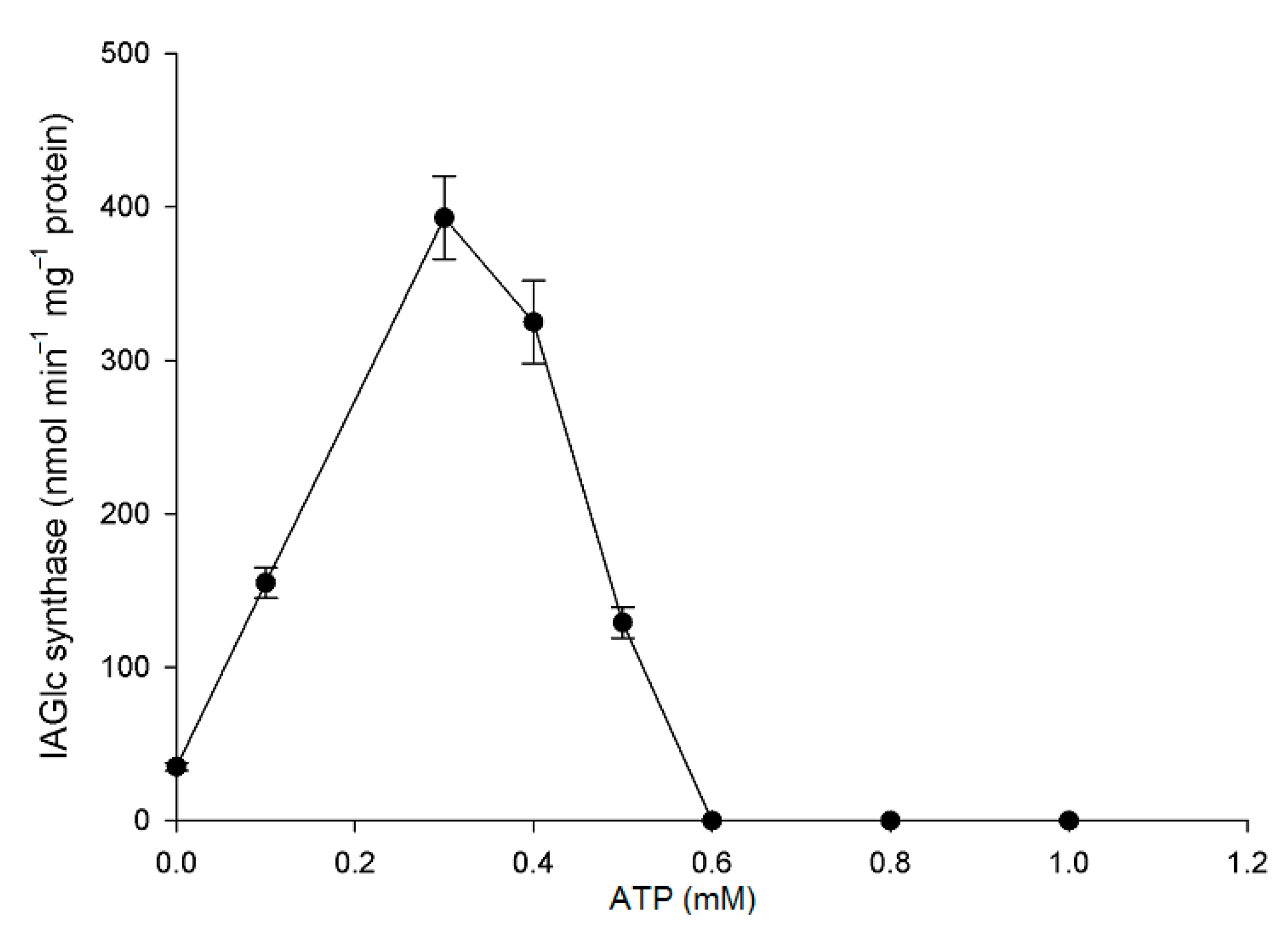
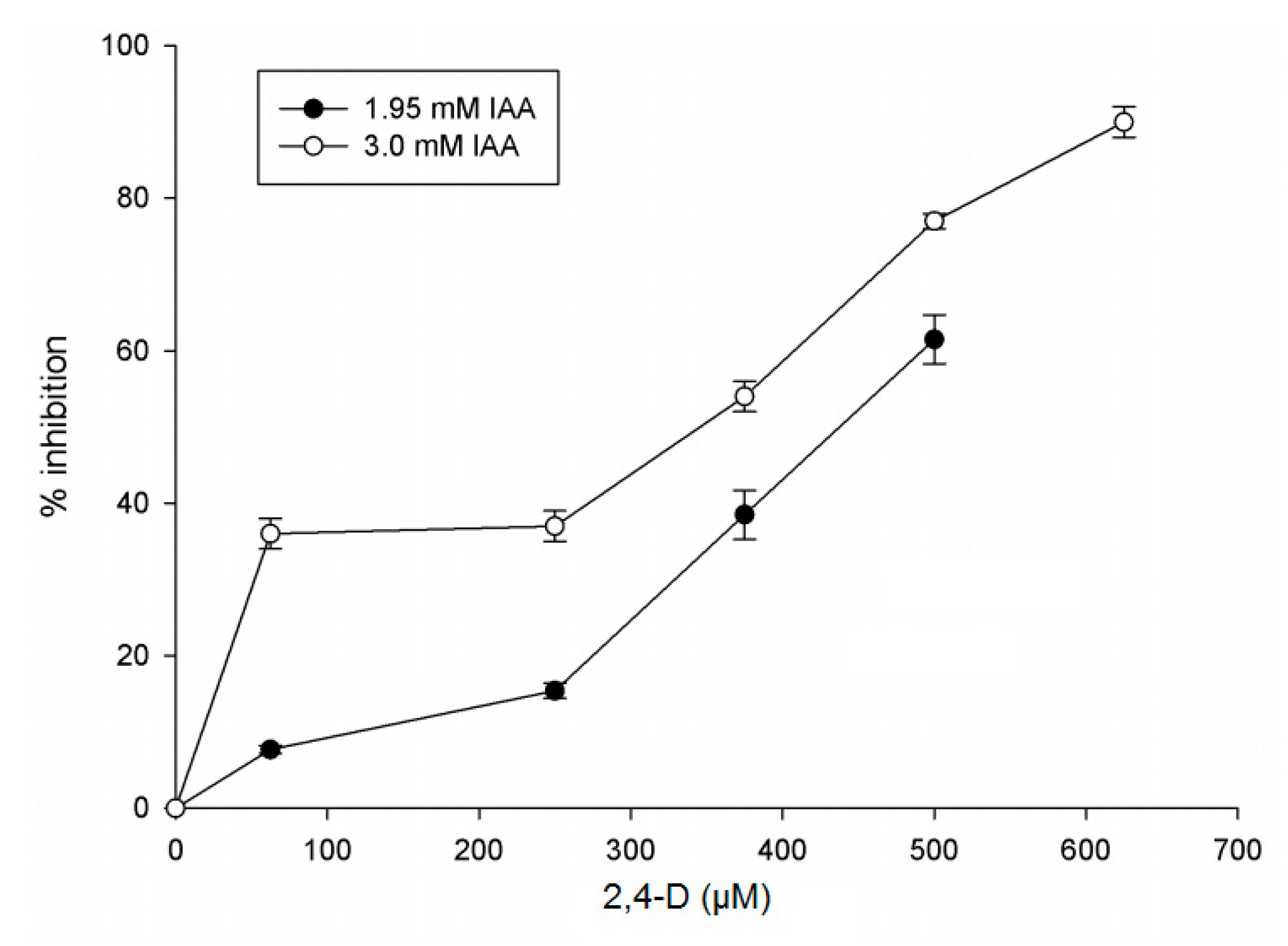
2.2.5. Effect of Modulators on IAGlc Synthase
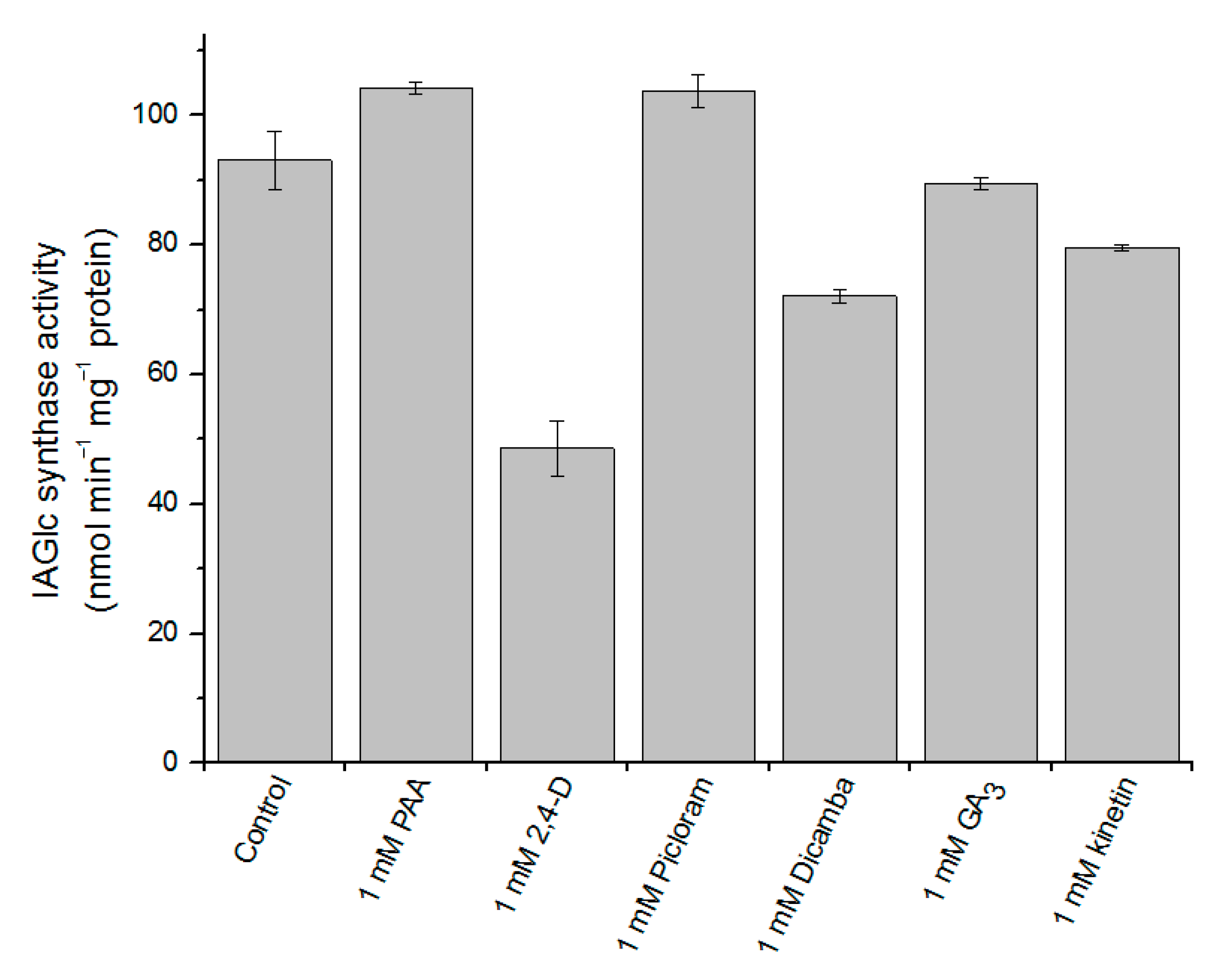
2.2.6. Effect of Phytohormones on IAGlc Synthase
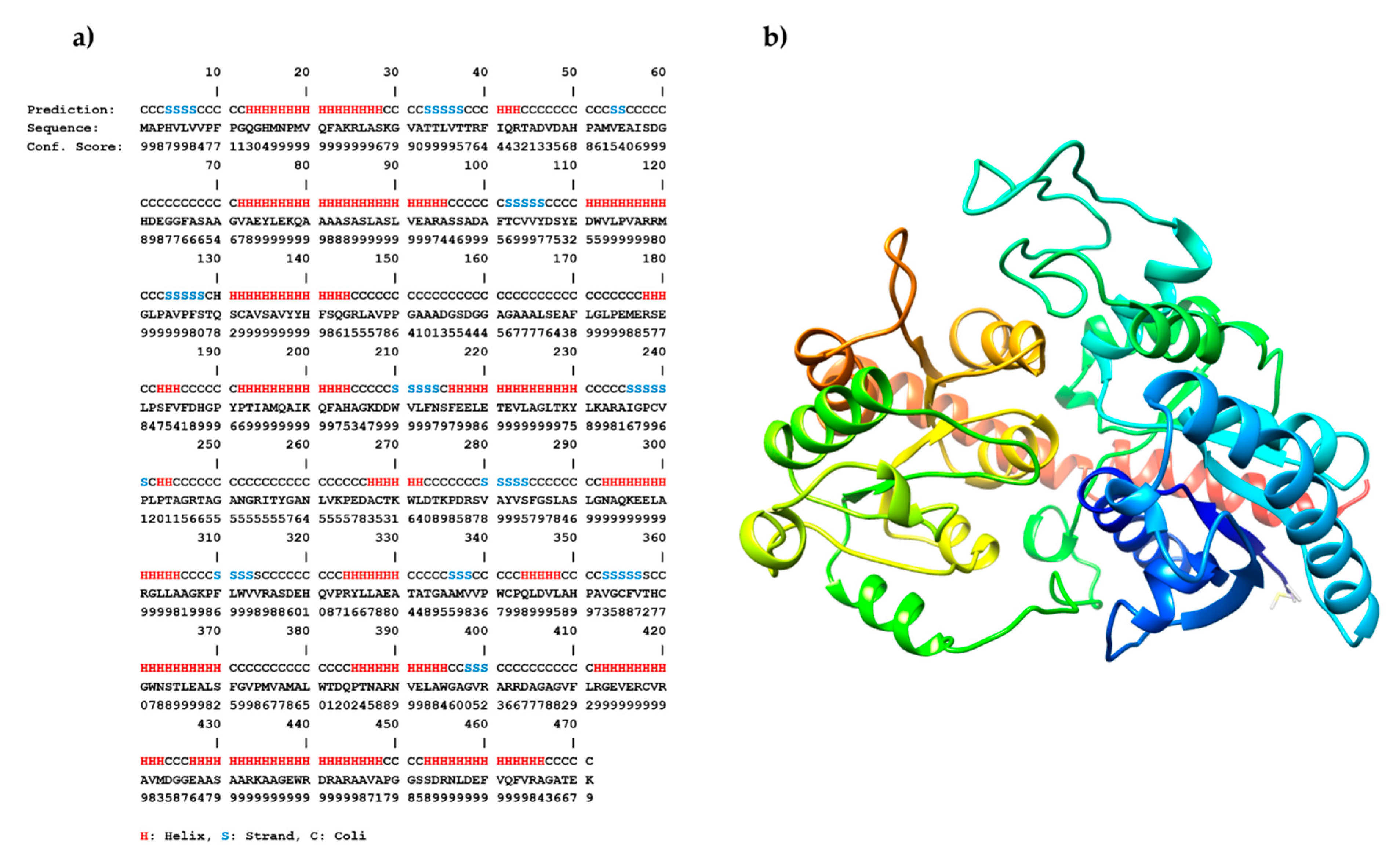
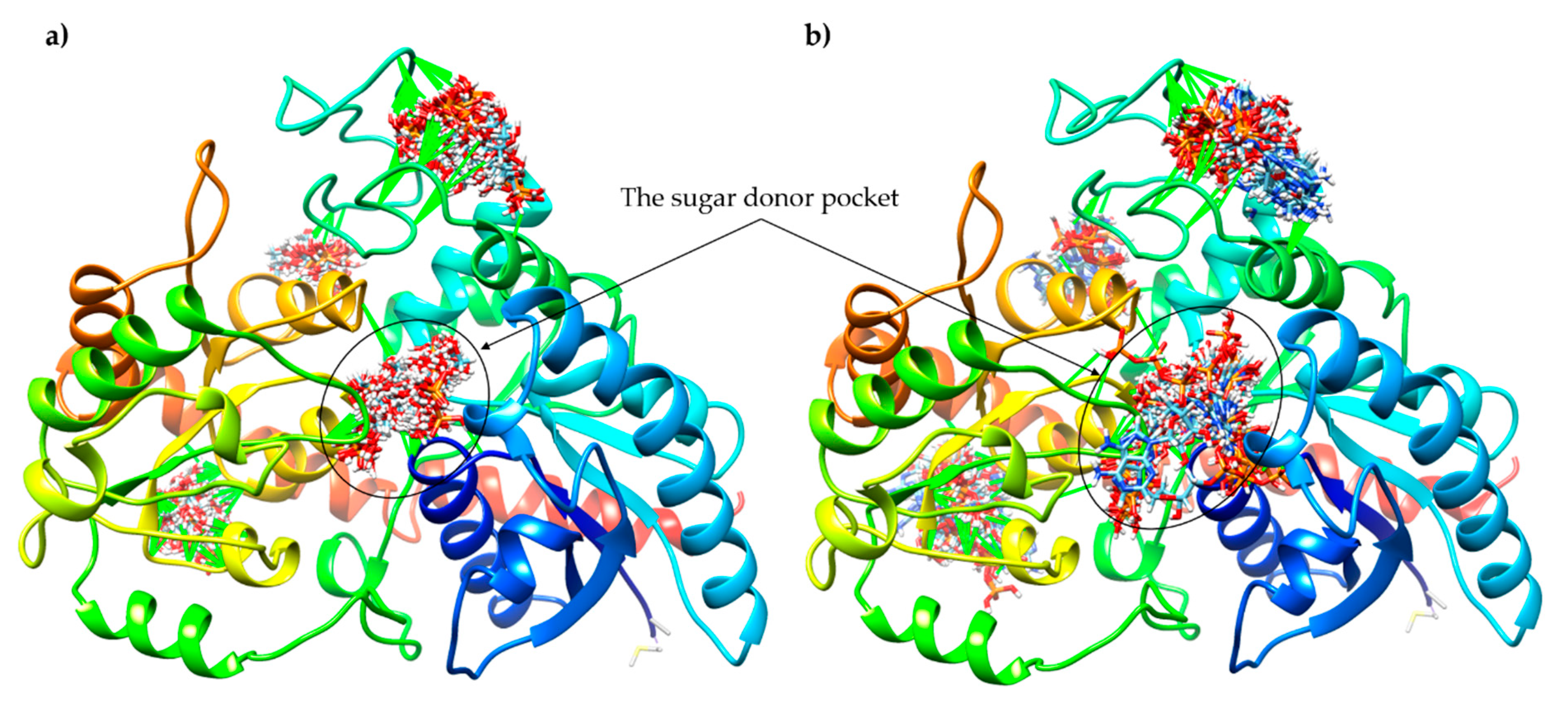
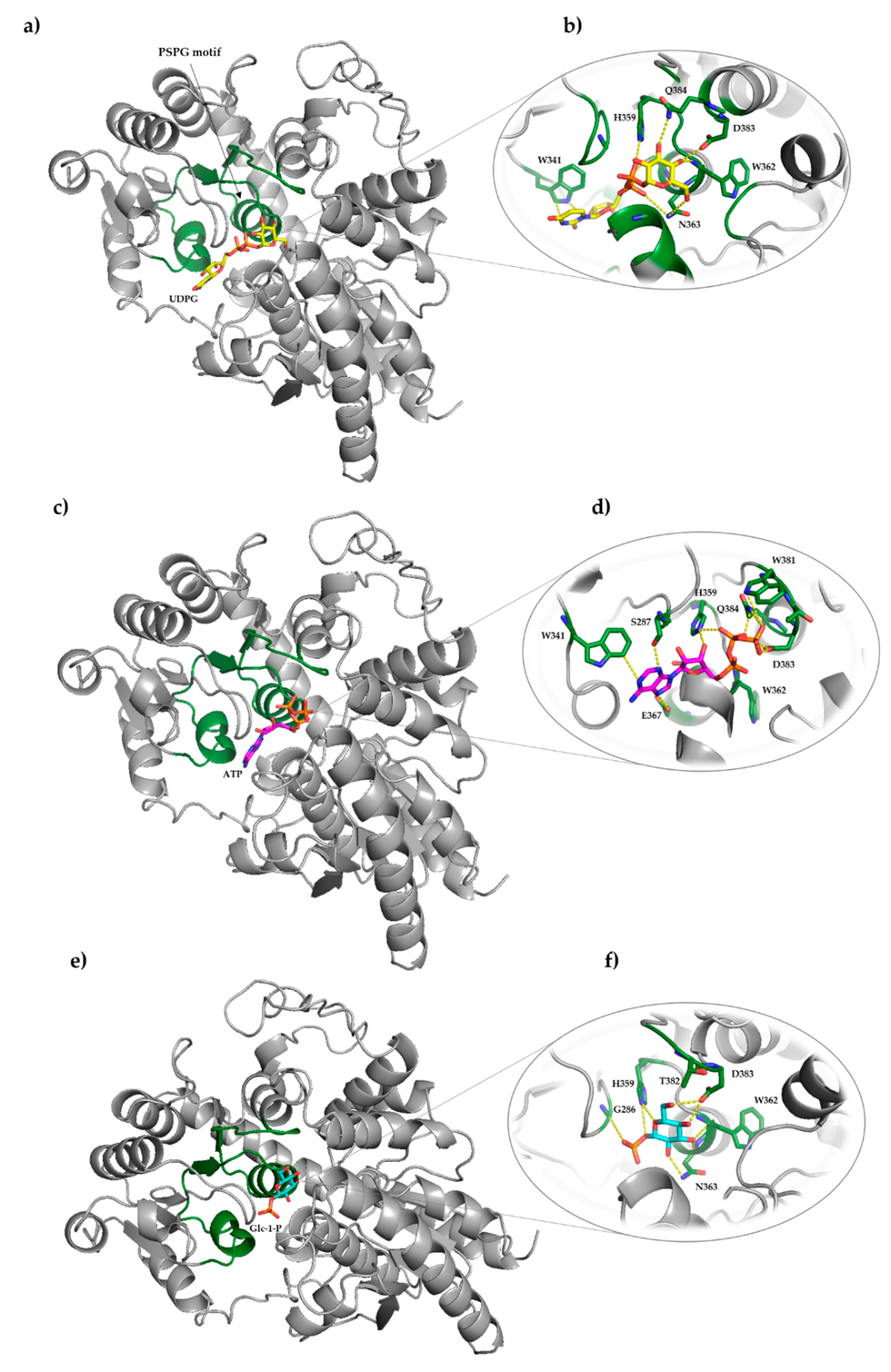
2.3. Sequence Analysis, 3D Structure Prediction and Molecular Docking Studies of Maize IAGlc Synthase
3. Materials and Methods
3.1. Expression of Zm-IAGLU in E. coli and Purification of Recombinant IAGlc Synthase
3.2. Enzyme Activity Assay
3.2.1. Determination of IAGlc Synthase Activity
3.2.2. Determination of UDPG Pyrophosphorylase Activity
3.3. Kinetic Studies
3.3.1. The Effect of pH on IAGlc Synthase Activity
3.3.2. The Effect of Divalent Cations on IAGlc Synthase Activity
3.3.3. The Effect of Modulators on IAGlc Synthase Activity
3.3.4. The Effect of Phytohormones on IAGlc Synthase Activity
3.3.5. Determination of Kinetic Parameters for UDPG
3.3.6. Determination of Kinetic Parameters for IAA
3.3.7. Determination of Substrate Specificity of IAGlc Synthase
3.3.8. Inhibition of IAGlc by ATP and Glucose-1-Phosphate
3.3.9. Inhibition of IAGlc Synthase by 2,4-D
3.4. Western Blot Analysis
3.5. Protein Determination
3.6. In Silico Sequence Analysis
3.7. Homology Modeling and Molecular Docking
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 1-O-IAGlc | 1-O-indole-3-acetyl-β-d-glucose |
| 2,4-D | 2,4-dichlorophenoxyacetic acid |
| ATP | adenosine triphosphate |
| GA | gibberellic acid |
| GSH | reduced glutathione |
| GSSG | oxidized glutathione |
| GT | glycosyltransferase |
| IAA | indole-3-acetic acid |
| IBA | indole-3-butyric acid |
| IPA | indole-3-propionic acid |
| IPyA | indole-3-pyruvic acid |
| IPTG | isopropyl-1-thio-β-d-galactopyranoside |
| NAA | naphthaleneacetic acid |
| OxIAA | oxindole-3-acetic acid |
| PAA | phenylacetic acid |
| PEP | phosphoenolopyruvate |
| PPi | pyrophosphate |
| PSPG | plant secondary product glycosyltransferase |
| UDP | uridine diphosphate |
| UDPG | uridine diphosphate glucose |
| UGT | uridine diphosphate glycosyltransferase |
| Zm-IAGlc | UDPG-dependent IAA glucosyltransferase gene |
References
- Breton, C.; Fournel-Gigleux, S.; Palcic, M.M. Recent structures, evolution and mechanisms of glycosyltransferases. Curr. Opin. Struct. Biol. 2012, 22, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.; Li, Y.; Lim, E.K.; Bowles, D.J. Higher plant glycosyltransferases. Genome Biol. 2001, 2, 3004. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, P.I.; Owens, I.S.; Burchell, B.; Bock, K.W.; Bairoch, A.; Belanger, A.; Gigleux, S.F.; Green, M.; Hum, D.W.; Iyanagi, T.; et al. The UDP glycosyltransferase gene superfamily: Recommended nomenclature update based on evolutionary divergence. Pharmacogenetics 1997, 7, 255–269. [Google Scholar] [CrossRef]
- Osmani, S.A.; Bak, S.; Møller, B.L. Substrate specificity of plant UDP-dependent glycosyltransferases predicted from crystal structures and homology modeling. Phytochemistry 2009, 70, 325–347. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Hanada, K. An evolutionary view of functional diversity in family 1 glycosyltransferases. Plant J. 2011, 66, 182–193. [Google Scholar] [CrossRef]
- Korasick, D.A.; Enders, T.A.; Strader, L.C. Auxin biosynthesis and storage forms. J. Exp. Bot. 2013, 64, 2541–2555. [Google Scholar] [CrossRef] [Green Version]
- Bajguz, A.; Piotrowska, A. Conjugates of auxin and cytokinin. Phytochemistry 2009, 70, 957–969. [Google Scholar] [CrossRef]
- Ludwig-Müller, J. Auxin conjugates: Their role for plant development and in the evolution of land plants. J. Exp. Bot. 2011, 62, 1757–1773. [Google Scholar] [CrossRef] [Green Version]
- Michalczuk, L.; Bandurski, R.S. UDP-glucose: Indoleacetic acid glucosyl transferase and indoleacetyl-glucose: Myo-inositol indoleacetyl transferase. Biochem. Biophys. Res. Commun. 1980, 93, 588–592. [Google Scholar] [CrossRef]
- Kęsy, J.M.; Bandurski, R.S. Partial purification and characterization of indol-3-ylacetylglucose: Myo-inositol in-dol-3-ylacetyltransferase (indoleacetic acid-inositol synthase). Plant. Physiol. 1990, 94, 1598–1604. [Google Scholar] [CrossRef] [Green Version]
- Kowalczyk, S.; Jakubowska, A.; Zielińska, E.; Bandurski, R.S. Bifunctional indole-3-acetyl transferase catalyses synthesis and hydrolysis of indole-3-acetyl-myo-inositol in immature endosperm of Zea mays. Physiol. Plant. 2003, 119, 165–174. [Google Scholar] [CrossRef]
- Leźnicki, A.J.; Bandurski, R.S. Enzymic synthesis of indole-3-acetyl-1-O-β-D-glucose. I. Partial purification and characterization of the enzyme from Zea mays. Plant. Physiol. 1988, 88, 1474–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leznicki, A.J.; Bandurski, R.S. Enzymic Synthesis of Indole-3-Acetyl-1-O-β-d-Glucose. II. Metabolic Characteristics of the Enzyme. Plant Physiol. 1988, 88, 1481–1485. [Google Scholar] [CrossRef] [PubMed]
- Kowalczyk, S.; Bandurski, R.S. Enzymic synthesis of 1-O-(indol-3-ylacetyl)-β-d-glucose. Purification of the enzyme from Zea mays, and preparation of antibodies to the enzyme. Biochem. J. 1991, 279, 509–514. [Google Scholar] [CrossRef]
- Szerszen, J.B.; Szczyglowski, K.; Bandurski, R.S. iaglu, a gene from Zea mays involved in conjugation of growth hormone indole-3-acetic acid. Science 1994, 265, 1699–1701. [Google Scholar] [CrossRef]
- Jackson, R.G.; Lim, E.K.; Li, Y.; Kowalczyk, M.; Sandberg, G.; Hoggett, J.; Ashford, D.A.; Bowles, D.J. Identification and Biochemical Characterization of an Arabidopsis Indole-3-acetic Acid Glucosyltransferase. J. Biol. Chem. 2001, 276, 4350–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tognetti, V.B.; Van Aken, O.; Morreel, K.; Vandenbroucke, K.; Van De Cotte, B.; De Clercq, I.; Chiwocha, S.; Fenske, R.; Prinsen, E.; Boerjan, W.; et al. Perturbation of Indole-3-Butyric Acid Homeostasis by the UDP-Glucosyltransferase UGT74E2 Modulates Arabidopsis Architecture and Water Stress Tolerance. Plant Cell 2010, 22, 2660–2679. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, K.; Hayashi, K.I.; Natsume, M.; Kamiya, Y.; Sakakibara, H.; Kawaide, H.; Kasahara, H. UGT74D1 Catalyzes the Glucosylation of 2-Oxindole-3-Acetic Acid in the Auxin Metabolic Pathway in Arabidopsis. Plant Cell Physiol. 2014, 55, 218–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.Z.; Jin, S.H.; Jiang, X.Y.; Dong, R.R.; Li, P.; Li, Y.J.; Hou, B.K. Ectopic expression of UGT75D1, a glycosyltransferase preferring indole-3-butyric acid, modulates cotyledon development and stress tolerance in seed germination of Arabidopsis thaliana. Plant Mol. Biol. 2015, 90, 77–93. [Google Scholar] [CrossRef]
- Chen, L.; Huang, X.X.; Zhao, S.M.; Xiao, D.W.; Xiao, L.T.; Tong, J.H.; Wang, W.S.; Li, Y.J.; Ding, Z.; Hou, B.K. IPyA glucosylation mediates light and temperature signaling to regulate auxin-dependent hypocotyl elongation in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 6910–6917. [Google Scholar] [CrossRef]
- Ostrowski, M.; Hetmann, A.; Jakubowska, A. Indole-3-acetic acid UDP-glucosyltransferase from immature seeds of pea is involved in modification of glycoproteins. Phytochemistry 2015, 117, 25–33. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, T.T.; Xiao, D.W.; Zhao, S.M.; Lin, J.S.; Wang, T.; Li, Y.J.; Hou, B.K. OsIAGT1 is a glucosyltransferase gene involved in the glucose conjugation of auxin in rice. Rice 2019, 12, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostrowski, M.; Ciarkowska, A.; Dalka, A.; Wilmowicz, E.; Jakubowska, A. Biosynthesis pathway of indole-3-acetyl-myo-inositol during development of maize (Zea mays L.) seeds. J. Plant Physiol. 2020, 245, 153082. [Google Scholar] [CrossRef]
- Choi, M.S.; Koh, E.B.; Woo, M.O.; Piao, R.; Oh, C.S.; Koh, H.J. Tiller formation in rice is altered by overexpression of OsIAGLU gene encoding an IAA-conjugating enzyme or exogenous treatment of free IAA. J. Plant Biol. 2012, 55, 429–435. [Google Scholar] [CrossRef]
- Yu, X.L.; Wang, H.Y.; Leung, D.W.M.; He, Z.D.; Zhang, J.J.; Peng, X.X.; Liu, E.E. Overexpression of OsIAAGLU reveals a role for IAA–glucose conjugation in modulating rice plant architecture. Plant Cell Rep. 2019, 38, 731–739. [Google Scholar] [CrossRef]
- Jin, S.; Hou, B.; Zhang, G. The ectopic expression of Arabidopsis glucosyltransferase UGT74D1 affects leaf positioning through modulating indole-3-acetic acid homeostasis. Sci. Rep. 2021, 11, 1154. [Google Scholar] [CrossRef] [PubMed]
- Michalczuk, L.; Bandurski, R.S.; Corcuera, L.J.; Kowalczyk, S.; Keglevic, D.; Pokorný, M.; Gazarian, I.G.; Lagrimini, L.M.; Mellon, F.A.; Naldrett, M.J.; et al. Enzymic synthesis of 1-O-indol-3-ylacetyl-β-d-glucose and indol-3-ylacetyl-myo-inositol. Biochem. J. 1982, 207, 273–281. [Google Scholar] [CrossRef]
- Gasser, B.; Saloheimo, M.; Rinas, U.; Dragosits, M.; Rodríguez-Carmona, E.; Baumann, K.; Giuliani, M.; Parrilli, E.; Branduardi, P.; Lang, C.; et al. Protein folding and conformational stress in microbial cells producing recombinant proteins: A host comparative overview. Microb. Cell Factories 2008, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, S.H.; Ma, X.M.; Han, P.; Wang, B.; Sun, Y.G.; Zhang, G.Z.; Li, Y.J.; Hou, B.K. UGT74D1 Is a Novel Auxin Glycosyltransferase from Arabidopsis thaliana. PLoS ONE 2013, 8, e61705. [Google Scholar] [CrossRef]
- Ostrowski, M.; Świdziński, M.; Ciarkowska, A.; Jakubowska, A. IAA-amido synthetase activity and GH3 expression during development of pea seedlings. Acta Physiol. Plant. 2014, 36, 3029–3037. [Google Scholar] [CrossRef]
- Kowalczyk, S.; Jakubowska, A.; Bandurski, R.S. 1-Naphtalene acetic acid induces indole-3-ylacetylglucose synthase in Zea mays seedling tissue. Plant Growth Regul. 2002, 38, 127–134. [Google Scholar] [CrossRef]
- Aoi, Y.; Hira, H.; Hayakawa, Y.; Liu, H.; Fukui, K.; Dai, X.; Tanaka, K.; Hayashi, K.-I.; Zhao, Y.; Kasahara, H. UDP-glucosyltransferase UGT84B1 regulates the levels of indole-3-acetic acid and phenylacetic acid in Arabidopsis. Biochem. Biophys. Res. Commun. 2020, 532, 244–250. [Google Scholar] [CrossRef]
- Berlec, A.; Štrukelj, B. Current state and recent advances in biopharmaceutical production in Escherichia coli, yeasts and mammalian cells. J. Ind. Microbiol. Biotechnol. 2013, 40, 257–274. [Google Scholar] [CrossRef]
- Caputi, L.; Malnoy, M.; Goremykin, V.; Nikiforova, S.; Martens, S. A genome-wide phylogenetic reconstruction of family 1 UDP-glycosyltransferases revealed the expansion of the family during the adaptation of plants to life on land. Plant J. 2012, 69, 1030–1042. [Google Scholar] [CrossRef] [PubMed]
- Ono, E.; Homma, Y.; Horikawa, M.; Kunikane-Doi, S.; Imai, H.; Takahashi, S.; Kawai, Y.; Ishiguro, M.; Fukui, Y.; Nakayama, T. Functional Differentiation of the Glycosyltransferases That Contribute to the Chemical Diversity of Bioactive Flavonol Glycosides in Grapevines (Vitis vinifera). Plant Cell 2010, 22, 2856–2871. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y. I-TASSER server for protein 3D structure prediction. BMC Bioinform. 2008, 9, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Roy, A.; Zhang, Y. Protein–ligand binding site recognition using complementary binding-specific substructure comparison and sequence profile alignment. Bioinformatics 2013, 29, 2588–2595. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Grosdidier, A.; Zoete, V.; Michielin, O. SwissDock, a protein-small molecule docking web service based on EADock DSS. Nucleic Acids Res. 2011, 3, W270–W277. [Google Scholar] [CrossRef] [Green Version]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ogita, Z.I.; Markert, C.L. A miniaturized system for electrophoresis on polyacrylamide gels. Anal. Biochem. 1979, 99, 233–241. [Google Scholar] [CrossRef]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass Spectrometric Sequencing of Proteins from Silver-Stained Polyacrylamide Gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef]
- Ehmann, A. The Van-Urk-Salkowski reagent: A sensitive and specific chromogenic reagent for silica gel thin-layer chromato-graphic detection and identification of indole derivatives. J. Chromatogr. 1977, 132, 267–276. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera: A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeLano, W.L. Pymol: An open-source molecular graphics tool. CCP4 Newsl. Protein Crystallogr. 2002, 40, 82–92. [Google Scholar]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate | IAGlc Synthase (nmol min−1 mg−1 protein) | Relative Activity (%) |
|---|---|---|
| IAA | 210.46 ± 0.5 | 100 |
| IBA | 129.53 ± 1.04 | 66 |
| IPA | 104.65 ± 3.8 | 49.7 |
| IPyA | 5.81 ± 0.2 | 2.76 |
| PAA | 0 | 0 |
| 2,4-D | 0 | 0 |
| Dicamba | 0 | 0 |
| Picloram | 0 | 0 |
| Tryptophan | 0 | 0 |
| Substrate | Cosubstrate | KM (mM) | Vmax (nmol min−1 mg−1 protein) |
|---|---|---|---|
| IAA | 4 mM UDPG | 0.8 ± 0.1 | 519 ± 27 |
| UDPG | 3 mM IAA | 0.7 ± 0.09 | 243 ± 18 |
| Inhibitor | Inhibition Type | Ki (μM) | IC50 (μM) | KMapp (mM) |
|---|---|---|---|---|
| 2,4-D | Competitive (IAA) | 117 ± 10 | 350 ± 28 (for 3 mM IAA) 439 ± 33 (for 1.95 mM IAA) | 1.22 ± 0.08 (1.53-fold decrease of affinity) |
| UGT | Sequence Identity with Maize IAGlc Synthase | PSPG Motif Identity with Maize IAGlc Synthase |
|---|---|---|
| UGT76F1 | 27.55% | 59.09% |
| UGT84B1 | 34.10% | 61.36% |
| UGT75D1 | 35.27% | 65.91% |
| UGT74D1 | 39.82% | 65.91% |
| UGT74E2 | 40.41% | 75.00% |
| OsIAGT1 | 69.35% | 88.64% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciarkowska, A.; Ostrowski, M.; Kozakiewicz, A. Biochemical Characterization of Recombinant UDPG-Dependent IAA Glucosyltransferase from Maize (Zea mays). Int. J. Mol. Sci. 2021, 22, 3355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073355
Ciarkowska A, Ostrowski M, Kozakiewicz A. Biochemical Characterization of Recombinant UDPG-Dependent IAA Glucosyltransferase from Maize (Zea mays). International Journal of Molecular Sciences. 2021; 22(7):3355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073355
Chicago/Turabian StyleCiarkowska, Anna, Maciej Ostrowski, and Anna Kozakiewicz. 2021. "Biochemical Characterization of Recombinant UDPG-Dependent IAA Glucosyltransferase from Maize (Zea mays)" International Journal of Molecular Sciences 22, no. 7: 3355. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073355