
Molecular, Cellular and Functional Analysis of TRγ Chain along the European Sea Bass Dicentrarchus labrax Development

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
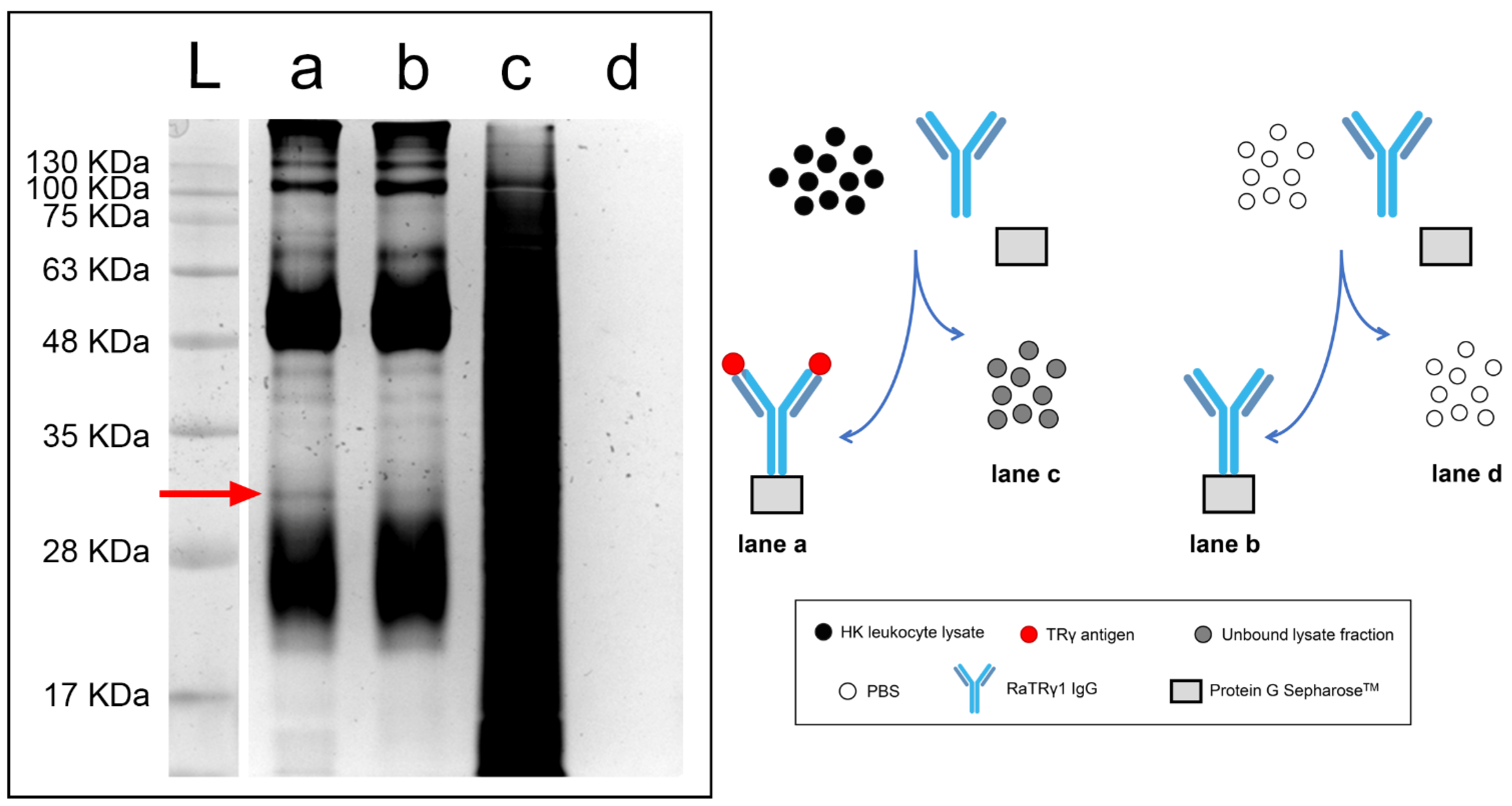
2.1. Antibody Production, ELISA Validation and IgG Enrichment
2.2. Immunoprecipitation
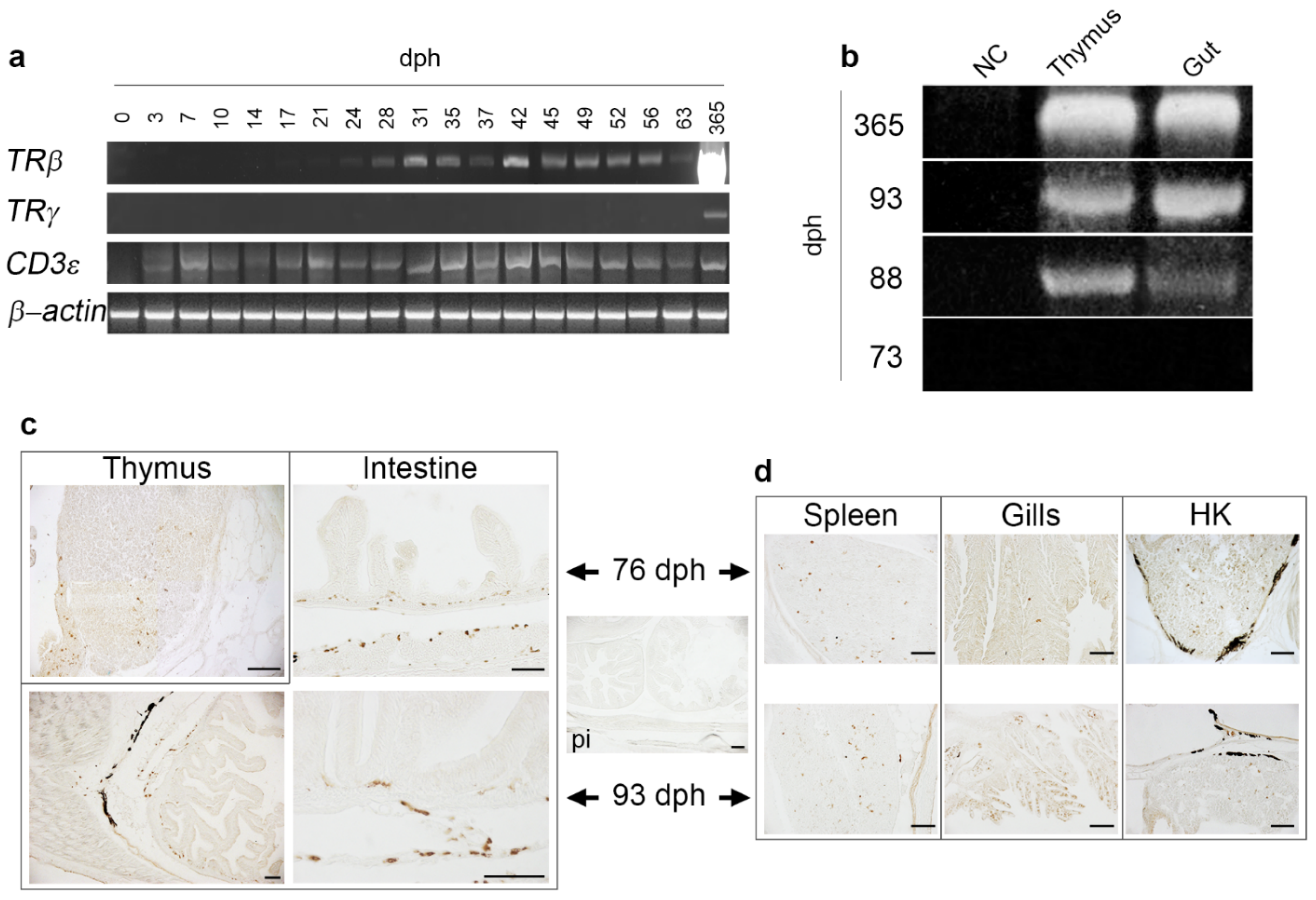
2.3. Expression Analysis of T Cell Markers along the Ontogeny
2.4. Localization of TRγ+ Cells in Developing Lymphoid Organs and Mucosal Tissues of Post-Larvae
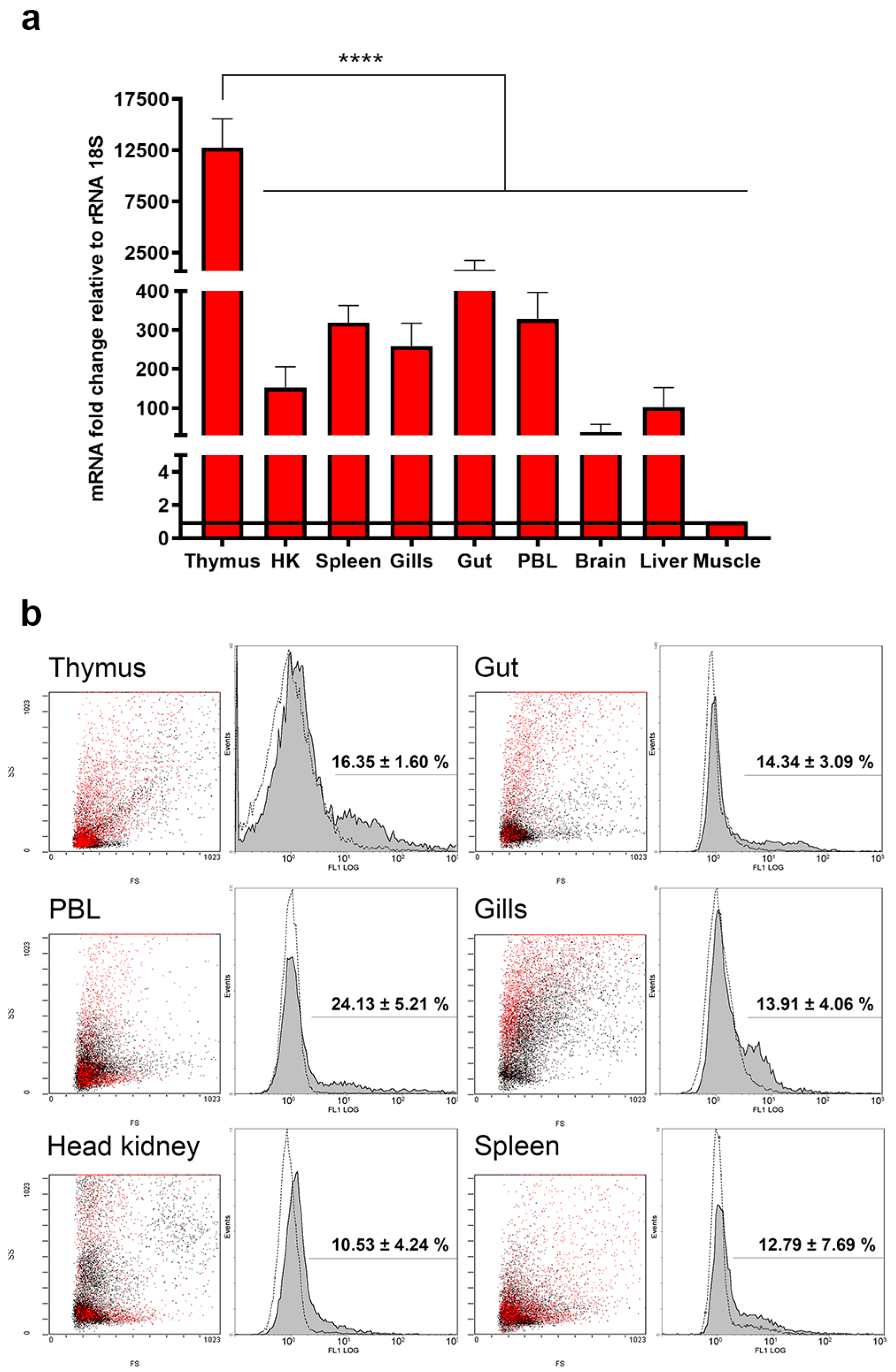
2.5. Transcriptional and Translational Quantification of TRγ in Lymphoid Organs and Mucosal Tissues of One-Year-Old Fish
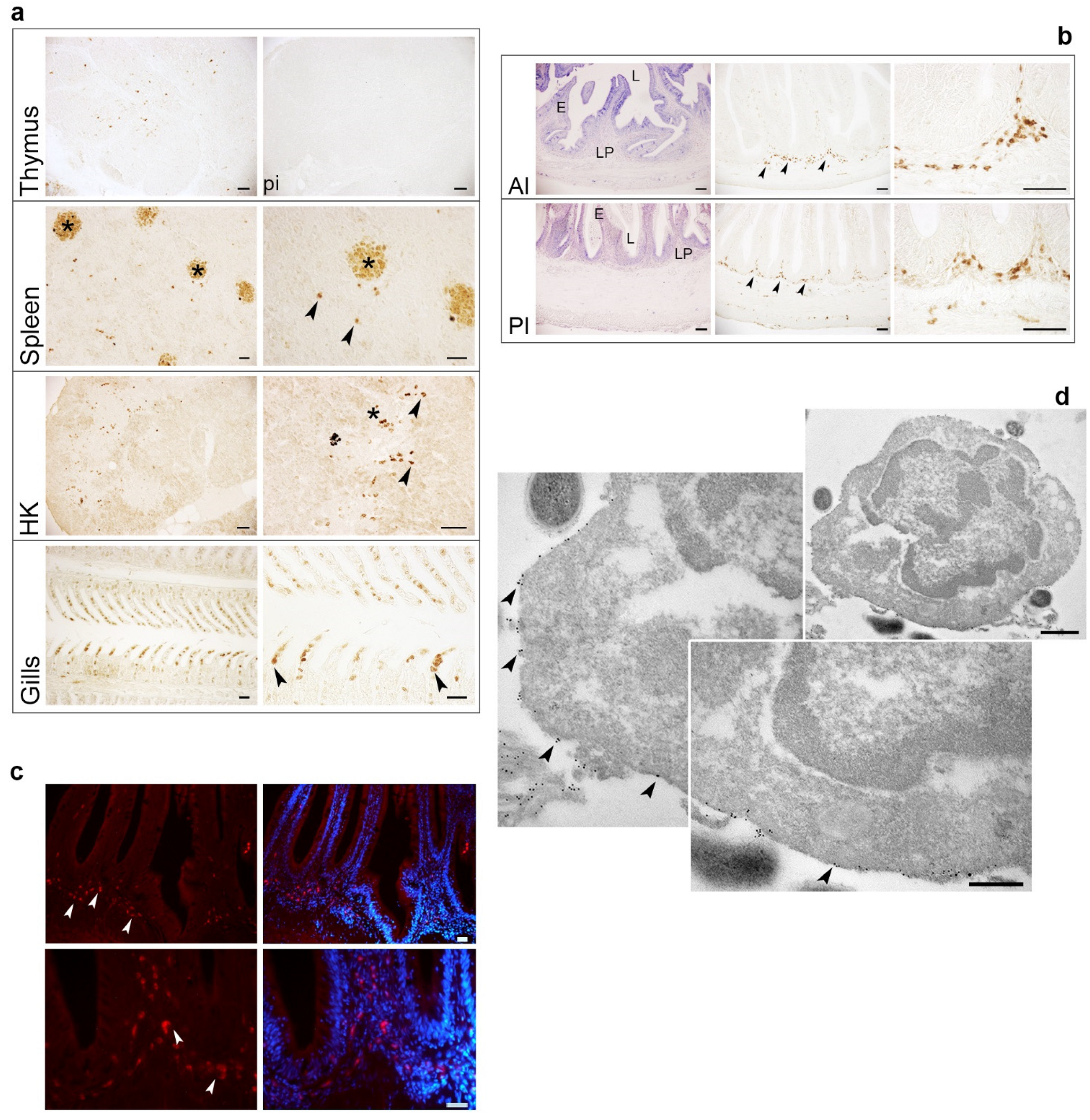
2.6. Localization of TRγ+ Cells in Lymphoid Organs and Mucosal Tissues of One-Year-Old Fish
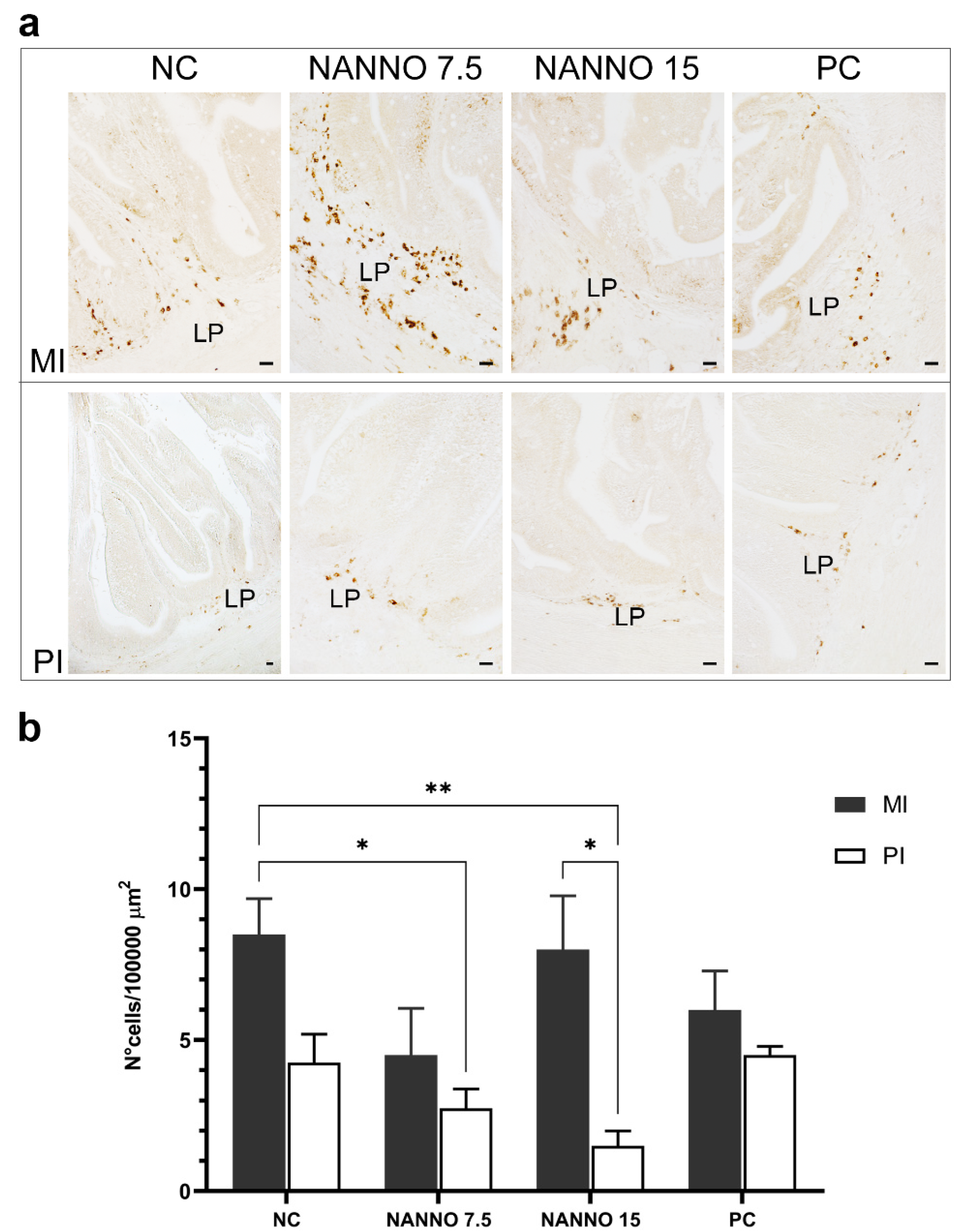
2.7. Effects of Marine Microalgae Nannochloropsis sp. Diets on Intestinal TRγ-Expressing Cells
3. Discussion
4. Materials and Methods
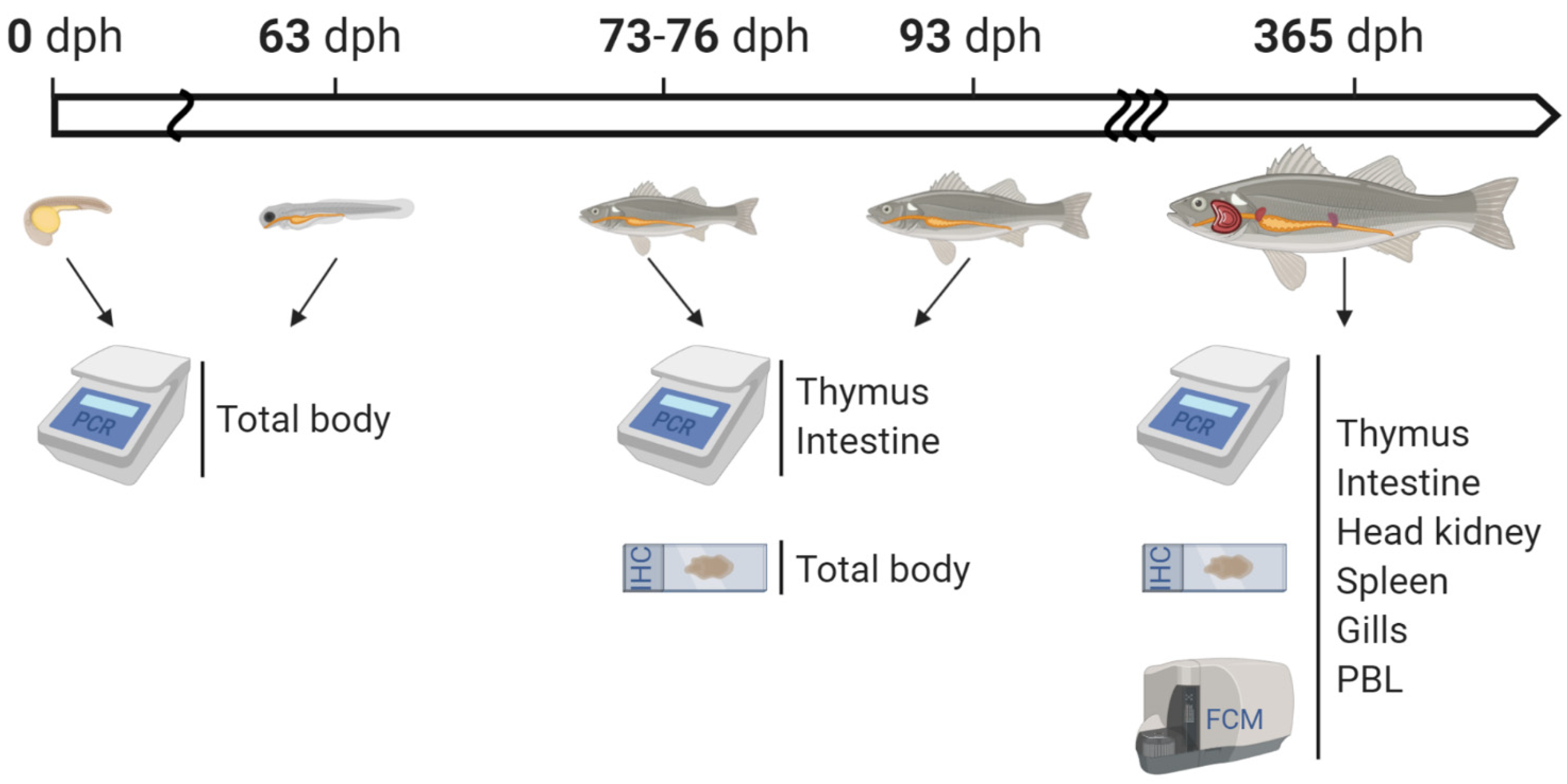
4.1. Animal Husbandry and Sampling
4.2. Production of Antisera Raised against Sea Bass TRγ Peptides
4.3. RNA Extraction and RT-PCR
4.4. Basal Expression Analysis of TRγ Transcripts in One-Year-Old Fish
4.5. Immunoprecipitation of Head Kidney Leukocyte Lysate with RaTRγ1 IgG
4.6. Flow Cytometry
4.7. Immunohistochemical Investigation on Post-Larvae and One-Year-Old Fish, and Immunofluorescence Analysis
4.8. Pre-Embedding Labeling of Cell Surface Antigens for Electron Microscopy
4.9. Diet Trials
4.10. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AI | anterior intestine |
| ANOVA | analysis of variance |
| BSA | bovine serum albumin |
| BW | body weight |
| CHAPS | 3-((3-cholamidopropyl) dimethylammonio)-1-propanesulfonate |
| DAPI | 4′,6-diamidino-2-phenylindole |
| DP | double positive |
| dph | day post hatching |
| DN | double negative |
| E | epithelium |
| HBSS | Hank’s Balanced Salt Solution without calcium and magnesium |
| HK | head kidney |
| kDa | kilo Dalton |
| IIF | indirect immunofluorescence |
| LP | lamina propria |
| L | lumen |
| MHC | major histocompatibility complex |
| MI | mid intestine |
| MW | molecular weight |
| MMCs | melano-macrophage centres |
| NC | negative control |
| NGS | normal goat serum |
| O/N | overnight |
| PAb | polyclonal antibody |
| PBL | peripheral blood lymphocytes |
| PBS | phosphate-buffered saline |
| PBT | PBS, Triton and BSA |
| PC | positive control |
| PFA | paraformaldehyde |
| PI | posterior intestine |
| PMSF | phenylmethylsulfonyl fluoride |
| RT | room temperature |
| TcR | T cell receptor |
| TR | T cell receptor chain |
References
- Birnbaum, M.E.; Berry, R.; Hsiao, Y.-S.; Chen, Z.; Shingu-Vazquez, M.A.; Yu, X.; Waghray, D.; Fischer, S.; McCluskey, J.; Rossjohn, J.; et al. Molecular Architecture of the Aβ T Cell Receptor–CD3 Complex. Proc. Natl. Acad. Sci. USA 2014, 111, 17576–17581. [Google Scholar] [CrossRef]
- Gobel, T.W.; Chen, C.L.; Lahti, J.; Kubota, T.; Kuo, C.L.; Aebersold, R.; Hood, L.; Cooper, M.D. Identification of T-Cell Receptor Alpha-Chain Genes in the Chicken. Proc. Natl. Acad. Sci. USA 1994, 91, 1094–1098. [Google Scholar] [CrossRef]
- Fellah, J.S.; Kerfourn, F.; Dumay, A.-M.; Aubet, G.; Charlemagne, J. Structure and Diversity of the T-Cell Receptor α Chain in the Mexican Axolotl. Immunogenetics 1997, 45, 235–241. [Google Scholar] [CrossRef]
- Smelty, P.; Marchal, C.; Renard, R.; Sinzelle, L.; Pollet, N.; Dunon, D.; Jaffredo, T.; Sire, J.-Y.; Fellah, J.S. Identification of the Pre-T-Cell Receptor Chain in Nonmammalian Vertebrates Challenges the Structure-Function of the Molecule. Proc. Natl. Acad. Sci. USA 2010, 107, 19991–19996. [Google Scholar] [CrossRef]
- Wilson, M.; Zhou, H.; Bengtén, E.; Clem, L.; Stuge, T.; Warr, G.; Miller, N.W. T-Cell Receptors in Channel Catfish: Structure and Expression of TCR α and β Genes. Mol. Immunol. 1998, 35, 545–557. [Google Scholar] [CrossRef]
- Rast, J.P.; Litman, G.W. T-Cell Receptor Gene Homologs Are Present in the Most Primitive Jawed Vertebrates. Proc. Natl. Acad. Sci. USA 1994, 91, 9248–9252. [Google Scholar] [CrossRef] [PubMed]
- Scapigliati, G.; Romano, N.; Abelli, L.; Meloni, S.; Ficca, A.G.; Buonocore, F.; Bird, S.; Secombes, C.J. Immunopurification of T-Cells from Sea Bass Dicentrarchus labrax (L.). Fish Shellfish Immunol. 2000, 10, 329–341. [Google Scholar] [CrossRef]
- Nam, B.-H.; Hirono, I.; Aoki, T. The Four TCR Genes of Teleost Fish: The CDNA and Genomic DNA Analysis of Japanese Flounder (Paralichthys olivaceus) TCR α-, β-, γ-, and δ-Chains. J. Immunol. 2003, 170, 3081–3090. [Google Scholar] [CrossRef] [PubMed]
- Shang, N.; Sun, X.; Hu, W.; Wang, Y.; Guo, Q. Molecular Cloning and Characterization of Common Carp (Cyprinus carpio L.) TCRγ and CD3γ/δ Chains. Fish Shellfish Immunol. 2008, 24, 412–425. [Google Scholar] [CrossRef]
- Yazawa, R.; Cooper, G.A.; Beetz-Sargent, M.; Robb, A.; McKinnel, L.; Davidson, W.S.; Koop, B.F. Functional Adaptive Diversity of the Atlantic Salmon T-Cell Receptor Gamma Locus. Mol. Immunol. 2008. [Google Scholar] [CrossRef]
- Buonocore, F.; Castro, R.; Randelli, E.; Lefranc, M.-P.; Six, A.; Kuhl, H.; Reinhardt, R.; Facchiano, A.; Boudinot, P.; Scapigliati, G. Diversity, Molecular Characterization and Expression of T Cell Receptor γ in a Teleost Fish, the Sea Bass (Dicentrarchus labrax, L.). PLoS ONE 2012, 7, e47957. [Google Scholar] [CrossRef]
- Moulana, M.; Taylor, E.B.; Edholm, E.S.; Quiniou, S.M.A.; Wilson, M.; Bengtén, E. Identification and Characterization of TCRγ and TCRδ Chains in Channel Catfish, Ictalurus punctatus. Immunogenetics 2014, 66, 545–561. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.Y.; Qi, Z.T.; Wu, N.; Chang, M.X.; Nie, P. Complementary DNA Sequences of the Constant Regions of T-Cell Antigen Receptors α, β and γ in Mandarin Fish, Siniperca chuatsi Basilewsky, and Their Transcriptional Changes after Stimulation with Flavobacterium columnare. J. Fish Dis. 2014, 37, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Wan, F.; Hu, C.; Ma, J.; Gao, K.; Xiang, L.; Shao, J. Characterization of Γδ T Cells from Zebrafish Provides Insights into Their Important Role in Adaptive Humoral Immunity. Front. Immunol. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yu, W.; Yu, Y.; Dong, S.; Yin, Y.; Huang, Z.; Wan, X.; Zhang, L.; Yu, Y.; Ai, T.; et al. Molecular Characterization and Expression Analysis of T Cell Receptor (TCR) γ and δ Genes in Dojo Loach (Misgurnus anguillicaudatus) in Response to Bacterial, Parasitic and Fungal Challenge. Fish Shellfish Immunol. 2019, 86, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Boschi, I.; Randelli, E.; Buonocore, F.; Casani, D.; Bernini, C.; Fausto, A.M.; Scapigliati, G. Transcription of T Cell-Related Genes in Teleost Fish, and the European Sea Bass (Dicentrarchus labrax) as a Model. Fish Shellfish Immunol. 2011, 31, 655–662. [Google Scholar] [CrossRef]
- Lee, Y.M.; Lee, J.-H.; Noh, J.K.; Kim, H.C.; Park, C.-J.; Park, J.-W.; Hwang, I.J.; Kim, S.Y. Stage and Tissue Specific Expression of Four TCR Subunits in Olive Flounder (Paralichthys olivaceus). Dev. Reprod. 2013, 17, 329–335. [Google Scholar] [CrossRef]
- James, K.D.; Jenkinson, W.E.; Anderson, G. T-Cell Egress from the Thymus: Should I Stay or Should I Go? J. Leukoc. Biol. 2018. [Google Scholar] [CrossRef]
- Zhou, H.; Stuge, T.B.; Miller, N.W.; Bengten, E.; Naftel, J.P.; Bernanke, J.M.; Chinchar, V.G.; Clem, L.W.; Wilson, M. Heterogeneity of Channel Catfish CTL with Respect to Target Recognition and Cytotoxic Mechanisms Employed. J. Immunol. 2001, 167, 1325–1332. [Google Scholar] [CrossRef]
- Hayday, A.C. Γδ Cells: A Right Time and a Right Place for a Conserved Third Way of Protection. Annu. Rev. Immunol. 2000, 18, 975–1026. [Google Scholar] [CrossRef]
- Vavassori, S.; Kumar, A.; Wan, G.S.; Ramanjaneyulu, G.S.; Cavallari, M.; El Daker, S.; Beddoe, T.; Theodossis, A.; Williams, N.K.; Gostick, E.; et al. Butyrophilin 3A1 Binds Phosphorylated Antigens and Stimulates Human Γδ T Cells. Nat. Immunol. 2013, 14, 908–916. [Google Scholar] [CrossRef]
- Mak, T.W.; Ferrick, D.A. The Γδ T-Cell Bridge: Linking Innate and Acquired Immunity. Nat. Med. 1998, 4, 764–765. [Google Scholar] [CrossRef] [PubMed]
- Scapigliati, G.; Fausto, A.M.; Picchietti, S. Fish Lymphocytes: An Evolutionary Equivalent of Mammalian Innate-like Lymphocytes? Front. Immunol. 2018, 9, 1–8. [Google Scholar] [CrossRef]
- Becker, E.W. Micro-Algae as a Source of Protein. Biotechnol. Adv. 2007, 25, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Burnell, G.; Allan, G.L. New Technologies in Aquaculture: Improving Production Efficiency, Quality and Environmental Management; Woodhead Publishing: Sawston, Cambridge, UK, 2009; ISBN 9781845693848. [Google Scholar]
- Nakagawa, H.; Montgomery, W. Algae. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D., Eds.; CABI Publishing: Wallingford, Oxfordshire, UK, 2007; ISBN 1845931998. [Google Scholar]
- Dawood, M.A.O. Nutritional Immunity of Fish Intestines: Important Insights for Sustainable Aquaculture. Rev. Aquac. 2020, 1–22. [Google Scholar] [CrossRef]
- Cerezuela, R.; Guardiola, F.; Meseguer, J.; Esteban, M.Á. Enrichment of Gilthead Seabream (Sparus aurata L.) Diet with Microalgae: Effects on the Immune System. Fish Physiol. Biochem. 2012, 38, 1729–1739. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Buonocore, F.; Guerra, L.; Belardinelli, M.C.; De Wolf, T.; Couto, A.; Fausto, A.M.; Saraceni, P.R.; Miccoli, A.; Scapigliati, G. Molecular and Cellular Characterization of European Sea Bass CD3ε+ T Lymphocytes and Their Modulation by Microalgal Feed Supplementation. Cell Tissue Res. 2021. [Google Scholar] [CrossRef]
- Gustafsson, K.; Herrmann, T.; Dieli, F. Editorial: Understanding Gamma Delta T Cell Multifunctionality-Towards Immunotherapeutic Applications. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef]
- Flajnik, M.F.; Kasahara, M. Origin and Evolution of the Adaptive Immune System: Genetic Events and Selective Pressures. Nat. Rev. Genet. 2010, 11, 47–59. [Google Scholar] [CrossRef]
- Litman, G.W.; Rast, J.P.; Fugmann, S.D. The Origins of Vertebrate Adaptive Immunity. Nat. Rev. Immunol. 2010, 10, 543–553. [Google Scholar] [CrossRef]
- Rast, J.P.; Anderson, M.K.; Strong, S.J.; Luer, C.; Litman, R.T.; Litman, G.W. A, Β, Γ, and Δ T Cell Antigen Receptor Genes Arose Early in Vertebrate Phylogeny. Immunity 1997, 6, 1–11. [Google Scholar] [CrossRef]
- Parra, Z.E.; Baker, M.L.; Schwarz, R.S.; Deakin, J.E.; Lindblad-Toh, K.; Miller, R.D. A Unique T Cell Receptor Discovered in Marsupials. Proc. Natl. Acad. Sci. USA 2007, 104, 9776–9781. [Google Scholar] [CrossRef]
- Parra, Z.E.; Baker, M.L.; Hathaway, J.; Lopez, A.M.; Trujillo, J.; Sharp, A.; Miller, R.D. Comparative Genomic Analysis and Evolution of the T Cell Receptor Loci in the Opossum Monodelphis domestica. BMC Genom. 2008, 9. [Google Scholar] [CrossRef]
- Wang, X.; Parra, Z.E.; Miller, R.D. Platypus TCR μ Provides Insight into the Origins and Evolution of a Uniquely Mammalian TCR Locus. J. Immunol. 2011, 187, 5246–5254. [Google Scholar] [CrossRef]
- Flajnik, M.F. Re-Evaluation of the Immunological Big Bang. Curr. Biol. 2014, 24, R1060–R1065. [Google Scholar] [CrossRef]
- Haegert, D.G. T cell antigen receptors. In Anticancer Research; Paul, W.E., Ed.; Wolters Kluwer Health/Lippincott Williams & Wilkins: Philadelphia, PA, USA, 1982; Volume 2, pp. 219–226. ISBN 9781451117837. [Google Scholar]
- Vantourout, P.; Hayday, A. Six-of-the-Best: Unique Contributions of Γδ T Cells to Immunology. Nat. Rev. Immunol. 2013, 13, 88–100. [Google Scholar] [CrossRef]
- Hayday, A.C. Γδ T Cells and the Lymphoid Stress-Surveillance Response. Immunity 2009, 31, 184–196. [Google Scholar] [CrossRef]
- Kalyan, S.; Kabelitz, D. Defining the Nature of Human Γδ T Cells: A Biographical Sketch of the Highly Empathetic. Cell. Mol. Immunol. 2013, 10, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hayday, A.C. Γδ T Cell Update: Adaptate Orchestrators of Immune Surveillance. J. Immunol. 2019, 203, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.D.; Witherden, D.A.; Havran, W.L. The Role of Tissue-Resident Γδ T Cells in Stress Surveillance and Tissue Maintenance. Cells 2020, 9, 686. [Google Scholar] [CrossRef] [PubMed]
- Uribe, C.; Folch, H.; Enriquez, R.; Moran, G. Innate and Adaptive Immunity in Teleost Fish: A Review. Vet. Med. (Praha) 2011, 56, 486–503. [Google Scholar] [CrossRef]
- Bowden, T.J.; Cook, P.; Rombout, J.H.W.M. Development and Function of the Thymus in Teleosts. Fish Shellfish Immunol. 2005, 19, 413–427. [Google Scholar] [CrossRef]
- Abelli, L.; Romano, N.; Mastrolia, L. Histology of developing thymus in sea bass Dicentrarchus labrax (L.). In Contributions to Animal Biology; Argano, R., Ed.; Halocynthia Association: Palermo, Italy, 1994; pp. 9–15. [Google Scholar]
- Abelli, L.; Picchietti, S.; Romano, N.; Mastrolia, L.; Scapigliati, G. Immunocytochemical Detection of Thymocyte Antigenic Determinants in Developing Lymphoid Organs of Sea Bass Dicentrarchus labrax (L.). Fish Shellfish Immunol. 1996, 6, 493–505. [Google Scholar] [CrossRef]
- Lam, S.H.; Chua, H.L.; Gong, Z.; Lam, T.J.; Sin, Y.M. Development and Maturation of the Immune System in Zebrafish, Danio rerio: A Gene Expression Profiling, in Situ Hybridization and Immunological Study. Dev. Comp. Immunol. 2004, 28, 9–28. [Google Scholar] [CrossRef]
- Picchietti, S.; Guerra, L.; Buonocore, F.; Randelli, E.; Fausto, A.M.; Abelli, L. Lymphocyte Differentiation in Sea Bass Thymus: CD4 and CD8-α Gene Expression Studies. Fish Shellfish Immunol. 2009, 27, 50–56. [Google Scholar] [CrossRef]
- Picchietti, S.; Guerra, L.; Selleri, L.; Buonocore, F.; Abelli, L.; Scapigliati, G.; Mazzini, M.; Fausto, A.M. Compartmentalisation of T Cells Expressing CD8α and TCRβ in Developing Thymus of Sea Bass Dicentrarchus labrax (L.). Dev. Comp. Immunol. 2008, 32, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Parker, M.E.; Ciofani, M. Regulation of Γδ T Cell Effector Diversification in the Thymus. Front. Immunol. 2020, 11. [Google Scholar] [CrossRef] [PubMed]
- Parra, Z.E.; Baker, M.L.; Lopez, A.M.; Trujillo, J.; Volpe, J.M.; Miller, R.D. TCRμ Recombination and Transcription Relative to the Conventional TCR during Postnatal Development in Opossums. J. Immunol. 2009, 182, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Ruiz, M.; Sumaria, N.; Pennington, D.J.; Silva-Santos, B. Thymic Determinants of Γδ T Cell Differentiation. Trends Immunol. 2017, 38, 336–344. [Google Scholar] [CrossRef]
- Chien, Y.H.; Zeng, X.; Prinz, I. The Natural and the Inducible: Interleukin (IL)-17-Producing Γδ T Cells. Trends Immunol. 2013, 34, 151–154. [Google Scholar] [CrossRef]
- Ciofani, M.; Zúñiga-Pflücker, J.C. Determining γ δ versus α β T Cell Development. Nat. Rev. Immunol. 2010, 10, 657–663. [Google Scholar] [CrossRef]
- Silva-Santos, B.; Pennington, D.J.; Hayday, A.C. Lymphotoxin-Mediated Regulation of Γδ Cell Differentiation by Aβ T Cell Progenitors. Science 2005, 307, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Shibata, K.; Yamada, H.; Nakamura, R.; Sun, X.; Itsumi, M.; Yoshikai, Y. Identification of CD25 + Γδ T Cells As Fetal Thymus-Derived Naturally Occurring IL-17 Producers. J. Immunol. 2008, 181, 5940–5947. [Google Scholar] [CrossRef] [PubMed]
- Muro, R.; Takayanagi, H.; Nitta, T. T Cell Receptor Signaling for ΓδT Cell Development. Inflamm. Regen. 2019, 39, 6. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Abelli, L.; Guerra, L.; Randelli, E.; Proietti Serafini, F.; Belardinelli, M.C.; Buonocore, F.; Bernini, C.; Fausto, A.M.; Scapigliati, G. MHC II-β Chain Gene Expression Studies Define the Regional Organization of the Thymus in the Developing Bony Fish Dicentrarchus labrax (L.). Fish Shellfish Immunol. 2015, 42, 483–493. [Google Scholar] [CrossRef]
- Bonneville, M.; O’Brien, R.L.; Born, W.K. γ δ T Cell Effector Functions: A Blend of Innate Programming and Acquired Plasticity. Nat. Rev. Immunol. 2010, 10, 467–478. [Google Scholar] [CrossRef]
- Jensen, K.D.C.; Su, X.; Shin, S.; Li, L.; Youssef, S.; Yamasaki, S.; Steinman, L.; Saito, T.; Locksley, R.M.; Davis, M.M.; et al. Thymic Selection Determines Γδ T Cell Effector Fate: Antigen-Naive Cells Make Interleukin-17 and Antigen-Experienced Cells Make Interferon γ. Immunity 2008, 29, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Khairallah, C.; Chu, T.H.; Sheridan, B.S. Tissue Adaptations of Memory and Tissue-Resident Gamma Delta T Cells. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Abelli, L.; Picchietti, S.; Romano, N.; Mastrolia, L.; Scapigliati, G. Immunohistochemistry of Gut-Associated Lymphoid Tissue of the Sea Bass Dicentrarchus labrax (L.). Fish Shellfish Immunol. 1997, 7, 235–245. [Google Scholar] [CrossRef]
- Picchietti, S.; Terribili, F.R.; Mastrolia, L.; Scapigliati, G.; Abelli, L. Expression of Lymphocyte Antigenic Determinants in Developing Gut-Associated Lymphoid Tissue of the Sea Bass Dicentrarchus labrax (L.). Anat. Embryol. 1997, 196, 457–463. [Google Scholar] [CrossRef]
- Picchietti, S.; Guerra, L.; Bertoni, F.; Randelli, E.; Belardinelli, M.C.; Buonocore, F.; Fausto, A.M.; Rombout, J.H.; Scapigliati, G.; Abelli, L. Intestinal T Cells of Dicentrarchus labrax (L.): Gene Expression and Functional Studies. Fish Shellfish Immunol. 2011, 30, 609–617. [Google Scholar] [CrossRef]
- Asigbetse, K.E.; Eigenmann, P.A.; Frossard, C.P. Intestinal Lamina Propria TcRγδ+ Lymphocytes Selectively Express IL-10 and IL-17. J. Investig. Allergol. Clin. Immunol. 2010, 20, 391–401. [Google Scholar] [PubMed]
- Bakke-McKellep, A.M.; Frøystad, M.K.; Lilleeng, E.; Dapra, F.; Refstie, S.; Krogdahl, Å.; Landsverk, T. Response to Soy: T-Cell-like Reactivity in the Intestine of Atlantic Salmon, Salmo Salar L. J. Fish Dis. 2007, 30, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Fausto, A.M.; Randelli, E.; Carnevali, O.; Taddei, A.R.; Buonocore, F.; Scapigliati, G.; Abelli, L. Early Treatment with Lactobacillus delbrueckii Strain Induces an Increase in Intestinal T-Cells and Granulocytes and Modulates Immune-Related Genes of Larval Dicentrarchus labrax (L.). Fish Shellfish Immunol. 2009, 26, 368–376. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.S.; Bukowski, J.F.; Milner, J. Bioactive Food Components That Enhance Γδ T Cell Function May Play a Role in Cancer Prevention. J. Nutr. 2008, 138, 1–4. [Google Scholar] [CrossRef]
- Calduch-Giner, J.A.; Sitjà-Bobadilla, A.; Pérez-Sánchez, J. Gene Expression Profiling Reveals Functional Specialization along the Intestinal Tract of a Carnivorous Teleostean Fish (Dicentrarchus labrax). Front. Physiol. 2016, 7. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi-Miyashita, M.; Shimada, S.I.; Kurosu, H.; Kato-Nagaoka, N.; Matsuoka, Y.; Ohwaki, M.; Ishikawa, H.; Nanno, M. An Accessory Role of TCRγδ+ Cells in the Exacerbation of Inflammatory Bowel Disease in TCRα Mutant Mice. Eur. J. Immunol. 2001, 31, 980–988. [Google Scholar] [CrossRef]
- Falk, M.C.; Ng, G.; Zhang, G.Y.; Fanning, G.C.; Kamath, K.R.; Knight, J.F. Predominance of T Cell Receptor Vδ3 in Small Bowel Biopsies from Coeliac Disease Patients. Clin. Exp. Immunol. 1994, 98, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, 2002–2007. [Google Scholar] [CrossRef] [PubMed]
- Buonocore, F.; Gerdol, M.; Pallavicini, A.; Stocchi, V.; Randelli, E.; Belardinelli, M.C.; Miccoli, A.; Saraceni, P.R.; Secombes, C.J.; Scapigliati, G.; et al. Identification, Molecular Characterization and Functional Analysis of Interleukin (IL)-2 and IL-2like (IL-2L) Cytokines in Sea Bass (Dicentrarchus labrax L.). Cytokine 2020, 126. [Google Scholar] [CrossRef] [PubMed]
- Picchietti, S.; Nuñez-Ortiz, N.; Stocchi, V.; Randelli, E.; Buonocore, F.; Guerra, L.; Scapigliati, G. Evolution of Lymphocytes. Immunoglobulin T of the Teleost Sea Bass (Dicentrarchus labrax): Quantitation of Gene Expressing and Immunoreactive Cells. Fish Shellfish Immunol. 2017, 63, 40–52. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primers Sequence 5′-3′ (Forward, FW, and Reverse, RV) | Accession Number |
|---|---|---|
| β-actin | ATGTACGTTGCCATCC (FW, RT) GAGATGCCACGCTCTC (RV, RT) | AJ493428 |
| 18S rRNA | CCAACGAGCTGCTGACC (FW, q) CCGTTACCCGTGGTCC (RV, q) | AY831388 |
| TRγ | CTGCTGTGTGTGGCCTCAGAC (FW, RT) GTGCTGGACGGAGCAGTGGTA (RV, RT) CTGCTGTGTGTGGCCTCAGAC (FW, q) AGCAAGAGAGTCCACAGCAGT (RV, q) | FR745889 |
| TRβ | AGATTACCGGACCATCAGTGAAAG (FW, RT) TCAGTAGTTCTGCTTTCCCTTTGA (RV, RT) | AJ493441 |
| CD3ε | CCTTACCACTGTAAATATGAGGACG (FW, RT) CAGGTTGACTCCGGGCTGCTG (RV, RT) | KX231274 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miccoli, A.; Guerra, L.; Pianese, V.; Saraceni, P.R.; Buonocore, F.; Taddei, A.R.; Couto, A.; De Wolf, T.; Fausto, A.M.; Scapigliati, G.; et al. Molecular, Cellular and Functional Analysis of TRγ Chain along the European Sea Bass Dicentrarchus labrax Development. Int. J. Mol. Sci. 2021, 22, 3376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073376
Miccoli A, Guerra L, Pianese V, Saraceni PR, Buonocore F, Taddei AR, Couto A, De Wolf T, Fausto AM, Scapigliati G, et al. Molecular, Cellular and Functional Analysis of TRγ Chain along the European Sea Bass Dicentrarchus labrax Development. International Journal of Molecular Sciences. 2021; 22(7):3376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073376
Chicago/Turabian StyleMiccoli, Andrea, Laura Guerra, Valeria Pianese, Paolo Roberto Saraceni, Francesco Buonocore, Anna Rita Taddei, Ana Couto, Tania De Wolf, Anna Maria Fausto, Giuseppe Scapigliati, and et al. 2021. "Molecular, Cellular and Functional Analysis of TRγ Chain along the European Sea Bass Dicentrarchus labrax Development" International Journal of Molecular Sciences 22, no. 7: 3376. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073376