
Gene Expression Changes of Humans with Primary Mitral Regurgitation and Reduced Left Ventricular Ejection Fraction

Abstract
:1. Introduction
2. Results
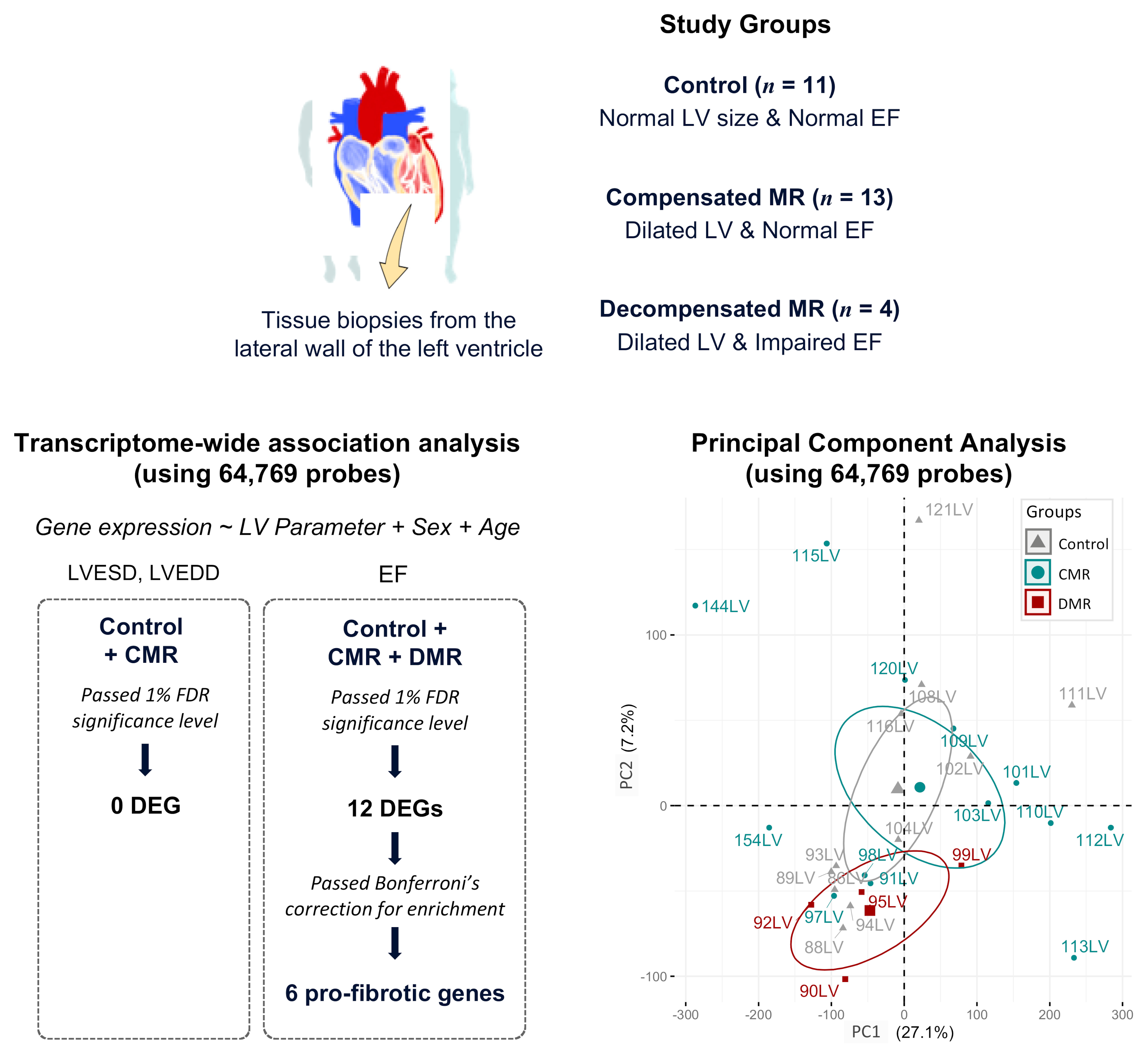
2.1. Study Participants
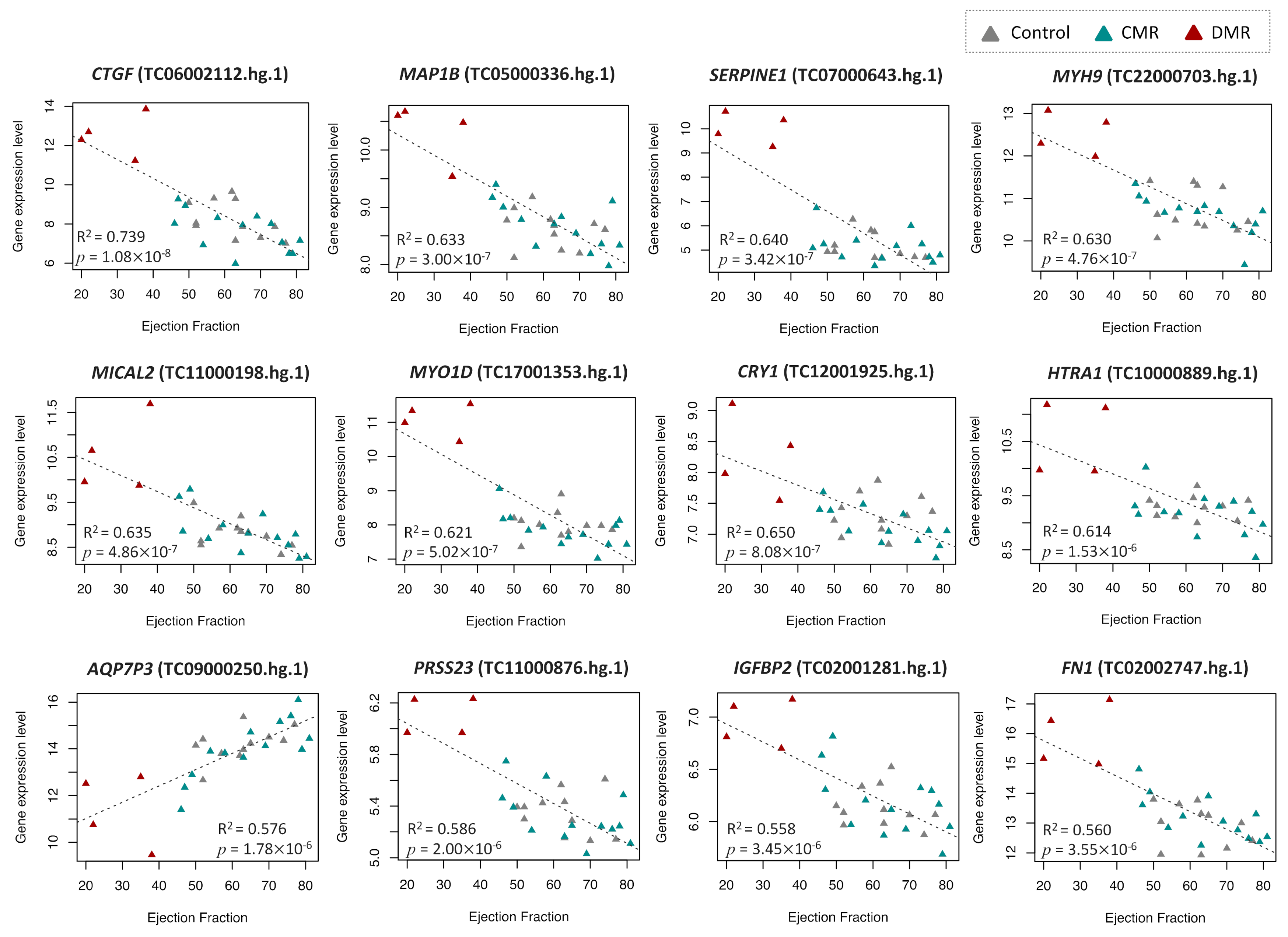
2.2. Identification of DEGs Associated with LV Dilatation and LV Dysfunction
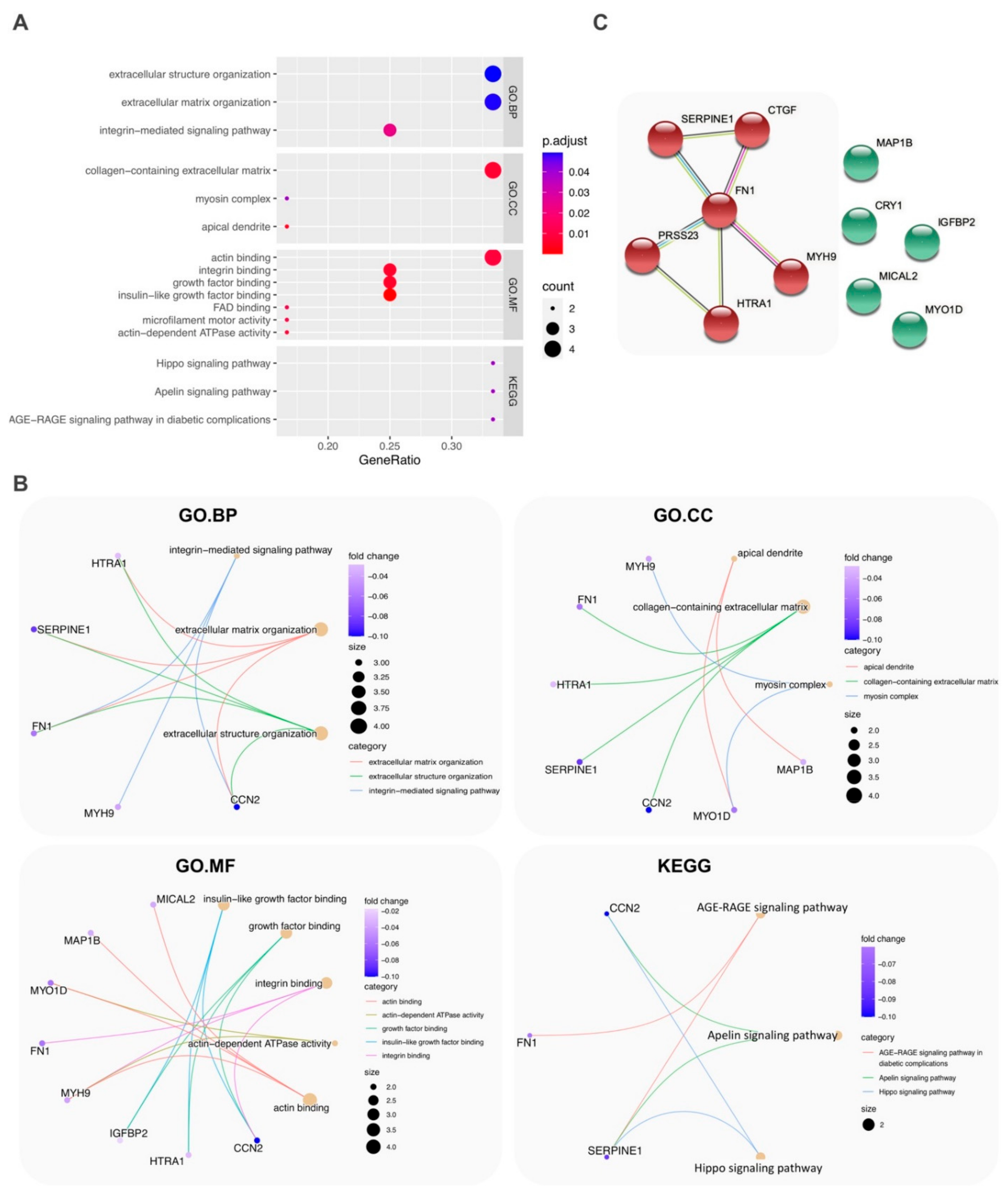
2.3. Functional and Pathway Enrichment Analysis
3. Discussion
4. Materials and Methods
4.1. Data Collection
4.2. RNA Isolation and Microarrays
4.3. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| FDR | False discovery rate |
| LV | Left ventricle |
| MR | Mitral regurgitation |
| ECM | Extracellular matrix |
| LVEDD | Left ventricular end-diastolic diameter |
| LVESD | Left ventricular end-systolic diameter |
| SF | Shortening fraction |
| EF | Ejection fraction |
References
- El Sabbagh, A.; Reddy, Y.N.V.; Nishimura, R.A. Mitral Valve Regurgitation in the Contemporary Era: Insights Into Diagnosis, Management, and Future Directions. JACC Cardiovasc. Imaging 2018, 11, 628–643. [Google Scholar] [CrossRef]
- McCutcheon, K.; Dickens, C.; van Pelt, J.; Dix-Peek, T.; Grinter, S.; McCutcheon, L.; Patel, A.; Hale, M.; Tsabedze, N.; Vachiat, A.; et al. Dynamic Changes in the Molecular Signature of Adverse Left Ventricular Remodeling in Patients With Compensated and Decompensated Chronic Primary Mitral Regurgitation. Circ. Heart Fail. 2019, 12, e005974. [Google Scholar] [CrossRef]
- Sweeney, M.; Corden, B.; Cook, S.A. Targeting cardiac fibrosis in heart failure with preserved ejection fraction: Mirage or miracle? EMBO Mol. Med. 2020, 12, e10865. [Google Scholar] [CrossRef]
- Zhou, S.G.; Ma, H.J.; Guo, Z.Y.; Zhang, W.; Yang, X. FHL2 participates in renal interstitial fibrosis by altering the phenotype of renal tubular epithelial cells via regulating the beta-catenin pathway. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2734–2741. [Google Scholar] [PubMed]
- Tamura, N.; Ogawa, Y.; Chusho, H.; Nakamura, K.; Nakao, K.; Suda, M.; Kasahara, M.; Hashimoto, R.; Katsuura, G.; Mukoyama, M.; et al. Cardiac fibrosis in mice lacking brain natriuretic peptide. Proc. Natl. Acad. Sci. USA 2000, 97, 4239–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagler, M.A.; Hadley, T.M.; Zhang, H.; Mehra, K.; Roos, C.M.; Schaff, H.V.; Suri, R.M.; Miller, J.D. TGF-beta signalling and reactive oxygen species drive fibrosis and matrix remodelling in myxomatous mitral valves. Cardiovasc. Res. 2013, 99, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Iwaki, T.; Urano, T.; Umemura, K. PAI-1, progress in understanding the clinical problem and its aetiology. Br. J. Haematol. 2012, 157, 291–298. [Google Scholar] [CrossRef]
- Valiente-Alandi, I.; Potter, S.J.; Salvador, A.M.; Schafer, A.E.; Schips, T.; Carrillo-Salinas, F.; Gibson, A.M.; Nieman, M.L.; Perkins, C.; Sargent, M.A.; et al. Inhibiting Fibronectin Attenuates Fibrosis and Improves Cardiac Function in a Model of Heart Failure. Circulation 2018, 138, 1236–1252. [Google Scholar] [CrossRef] [PubMed]
- Muratoglu, S.C.; Belgrave, S.; Hampton, B.; Migliorini, M.; Coksaygan, T.; Chen, L.; Mikhailenko, I.; Strickland, D.K. LRP1 protects the vasculature by regulating levels of connective tissue growth factor and HtrA1. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2137–2146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayoumi, A.S.; Teoh, J.P.; Aonuma, T.; Yuan, Z.; Ruan, X.; Tang, Y.; Su, H.; Weintraub, N.L.; Kim, I.M. MicroRNA-532 protects the heart in acute myocardial infarction, and represses prss23, a positive regulator of endothelial-to-mesenchymal transition. Cardiovasc. Res. 2017, 113, 1603–1614. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhu, M.; Chen, X.; Jiang, X. MYH9 Inhibition Suppresses TGF-beta1-Stimulated Lung Fibroblast-to-Myofibroblast Differentiation. Front. Pharmacol. 2020, 11, 573524. [Google Scholar] [CrossRef] [PubMed]
- Frangogiannis, N.G. The Extracellular Matrix in Ischemic and Nonischemic Heart Failure. Circ. Res. 2019, 125, 117–146. [Google Scholar] [CrossRef] [PubMed]
- Tsai, F.C.; Chang, G.J.; Lai, Y.J.; Chang, S.H.; Chen, W.J.; Yeh, Y.H. Ubiquitin Pathway Is Associated with Worsening Left Ventricle Function after Mitral Valve Repair: A Global Gene Expression Study. Int. J. Mol. Sci. 2020, 21, 5073. [Google Scholar] [CrossRef]
- Vainio, L.E.; Szabo, Z.; Lin, R.; Ulvila, J.; Yrjola, R.; Alakoski, T.; Piuhola, J.; Koch, W.J.; Ruskoaho, H.; Fouse, S.D.; et al. Connective Tissue Growth Factor Inhibition Enhances Cardiac Repair and Limits Fibrosis After Myocardial Infarction. JACC Basic Transl. Sci. 2019, 4, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Montenegro-Venegas, C.; Tortosa, E.; Rosso, S.; Peretti, D.; Bollati, F.; Bisbal, M.; Jausoro, I.; Avila, J.; Caceres, A.; Gonzalez-Billault, C. MAP1B regulates axonal development by modulating Rho-GTPase Rac1 activity. Mol. Biol. Cell 2010, 21, 3518–3528. [Google Scholar] [CrossRef] [Green Version]
- Giarratana, N.; Conti, F.; La Rovere, R.; Gijsbers, R.; Carai, P.; Duelen, R.; Vervliet, T.; Bultynck, G.; Ronzoni, F.; Piciotti, R.; et al. MICAL2 is essential for myogenic lineage commitment. Cell Death Dis. 2020, 11, 654. [Google Scholar] [CrossRef] [PubMed]
- Brunskill, E.W.; Georgas, K.; Rumballe, B.; Little, M.H.; Potter, S.S. Defining the molecular character of the developing and adult kidney podocyte. PLoS ONE 2011, 6, e24640. [Google Scholar] [CrossRef] [Green Version]
- Rabinovich-Nikitin, I.; Lieberman, B.; Martino, T.A.; Kirshenbaum, L.A. Circadian-Regulated Cell Death in Cardiovascular Diseases. Circulation 2019, 139, 965–980. [Google Scholar] [CrossRef]
- Verkerk, A.O.; Lodder, E.M.; Wilders, R. Aquaporin Channels in the Heart-Physiology and Pathophysiology. Int. J. Mol. Sci. 2019, 20, 2039. [Google Scholar] [CrossRef] [Green Version]
- Barutaut, M.; Fournier, P.; Peacock, W.F.; Evaristi, M.F.; Caubere, C.; Turkieh, A.; Desmoulin, F.; Eurlings, L.W.M.; van Wijk, S.; Rocca, H.B.; et al. Insulin-like Growth Factor Binding Protein 2 predicts mortality risk in heart failure. Int. J. Cardiol. 2020, 300, 245–251. [Google Scholar] [CrossRef]
- Storey, J.D.; Taylor, J.E.; Siegmund, D. Strong control, conservative point estimation and simultaneous conservative consistency of false discovery rates: A unified approach. J. R. Stat. Soc. Ser. B Stat. Methodol. 2004, 66, 187–205. [Google Scholar] [CrossRef]
- Yu, G.; Wang, L.G.; Han, Y.; He, Q.Y. clusterProfiler: An R package for comparing biological themes among gene clusters. OMICS J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Clinical Variable | Control n = 11 | CMR n = 13 | DMR n = 4 | p Value (CMR vs. Control) | p Value (DMR vs. Control) | p Value (DMR vs. CMR) |
|---|---|---|---|---|---|---|
| Age, years | 62.45 ± 11.34 | 59.46 ± 12.87 | 59.75 ± 13.96 | 0.551 | 0.555 | 0.970 |
| Sex (n, %) | ||||||
| Female | 7 (64) | 3 (23) | 2 (50) | 0.0953 | 0.999 | 0.5378 |
| Male | 4 (36) | 10 (77) | 2 (50) | |||
| Parameters for LV diameter | ||||||
| LVEDD, mm | 47.27 ± 4.03 | 57.85 ± 8.07 | 61.25 ± 14.93 | 5.86 × 10−4 | 0.157 | 0.554 |
| LVESD, mm | 31.18 ± 4.94 | 37.69 ± 9.27 | 54.00 ± 17.38 | 0.049 | 0.077 | 0.024 |
| Parameters for LV function | ||||||
| SF, % | 0.34 ± 0.07 | 0.35 ± 0.10 | 0.13 ± 0.10 | 0.767 | 3.30 × 10−4 | 1.20 × 10−3 |
| EF, % | 62.27 ± 9.01 | 64.46 ± 12.69 | 28.75 ± 9.07 | 0.637 | 2.49 × 10−5 | 1.12 × 10−4 |
| Probe Set ID | CHR | Start | End | β Coef. | S.E. | p Value | Gene Symbol | Description |
|---|---|---|---|---|---|---|---|---|
| LVEDD | ||||||||
| TC03001196.hg.1 | 3 | 14389951 | 14394068 | 0.025 | 0.004 | 1.70 × 10−5 | LINC01267 | long intergenic non-protein coding RNA 1267 |
| TC01002205.hg.1 | 1 | 11917521 | 11918992 | 0.298 | 0.065 | 1.68 × 10−4 | NPPB | natriuretic peptide precursor B |
| LVESD | ||||||||
| TC02002167.hg.1 | 2 | 105974169 | 106055230 | −0.037 | 0.006 | 6.67 × 10−6 | FHL2 | four and a half LIM domains 2 |
| TC03001196.hg.1 | 3 | 14389951 | 14394068 | 0.025 | 0.004 | 1.01 × 10−5 | LINC01267 | long intergenic non-protein coding RNA 1267 |
| EF | ||||||||
| TC06002112.hg.1 | 6 | 132269316 | 132272518 | −0.100 | 0.012 | 1.80 × 10−8 | CTGF | connective tissue growth factor |
| TC05000336.hg.1 | 5 | 71403061 | 71505397 | −0.036 | 0.005 | 3.00 × 10−7 | MAP1B | microtubule associated protein 1B |
| TC07000643.hg.1 | 7 | 100770370 | 100782547 | −0.092 | 0.013 | 3.43 × 10−7 | SERPINE1 | serpin peptidase inhibitor, clade E (nexin, plasminogen activator inhibitor type 1), member 1 |
| TC22000703.hg.1 | 22 | 36677323 | 36784063 | −0.040 | 0.006 | 4.76 × 10−7 | MYH9 | myosin, heavy chain 9, non-muscle |
| TC11000198.hg.1 | 11 | 12115543 | 12285332 | −0.038 | 0.006 | 4.86 × 10−7 | MICAL2 | microtubule associated monooxygenase, calponin and LIM domain containing 2 |
| TC17001353.hg.1 | 17 | 30819627 | 31203902 | −0.061 | 0.009 | 5.20 × 10−7 | MYO1D | myosin ID |
| TC12001925.hg.1 | 12 | 107385142 | 107487607 | −0.025 | 0.004 | 8.08 × 10−7 | CRY1 | cryptochrome circadian clock 1 |
| TC10000889.hg.1 | 10 | 124221041 | 124274424 | −0.029 | 0.005 | 1.53 × 10−6 | HTRA1 | HtrA serine peptidase 1 |
| TC09000250.hg.1 | 9 | 42858152 | 42893137 | 0.072 | 0.011 | 1.78 × 10−6 | AQP7P3 | aquaporin 7 pseudogene 3 |
| TC11000876.hg.1 | 11 | 86511282 | 86663886 | −0.016 | 0.003 | 2.00 × 10−6 | PRSS23 | protease, serine, 23 |
| TC02001281.hg.1 | 2 | 217497551 | 217529159 | −0.018 | 0.003 | 3.54 × 10−6 | IGFBP2 | insulin like growth factor binding protein 2 |
| TC02002747.hg.1 | 2 | 216225163 | 216300895 | −0.060 | 0.010 | 3.55 × 10−6 | FN1 | fibronectin 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, F.-C.; Chen, Y.-L.; Yen, K.-C.; Chiu, C.-H.; Chen, J.-H.; Yeh, Y.-H.; Tsai, P.-C. Gene Expression Changes of Humans with Primary Mitral Regurgitation and Reduced Left Ventricular Ejection Fraction. Int. J. Mol. Sci. 2021, 22, 3454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073454
Tsai F-C, Chen Y-L, Yen K-C, Chiu C-H, Chen J-H, Yeh Y-H, Tsai P-C. Gene Expression Changes of Humans with Primary Mitral Regurgitation and Reduced Left Ventricular Ejection Fraction. International Journal of Molecular Sciences. 2021; 22(7):3454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073454
Chicago/Turabian StyleTsai, Feng-Chun, Yu-Lin Chen, Kun-Chi Yen, Cheng-Hsun Chiu, Jui-Hsuan Chen, Yung-Hsin Yeh, and Pei-Chien Tsai. 2021. "Gene Expression Changes of Humans with Primary Mitral Regurgitation and Reduced Left Ventricular Ejection Fraction" International Journal of Molecular Sciences 22, no. 7: 3454. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073454