
Investigation of the Interaction of Human Origin Recognition Complex Subunit 1 with G-Quadruplex DNAs of Human c-myc Promoter and Telomere Regions

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
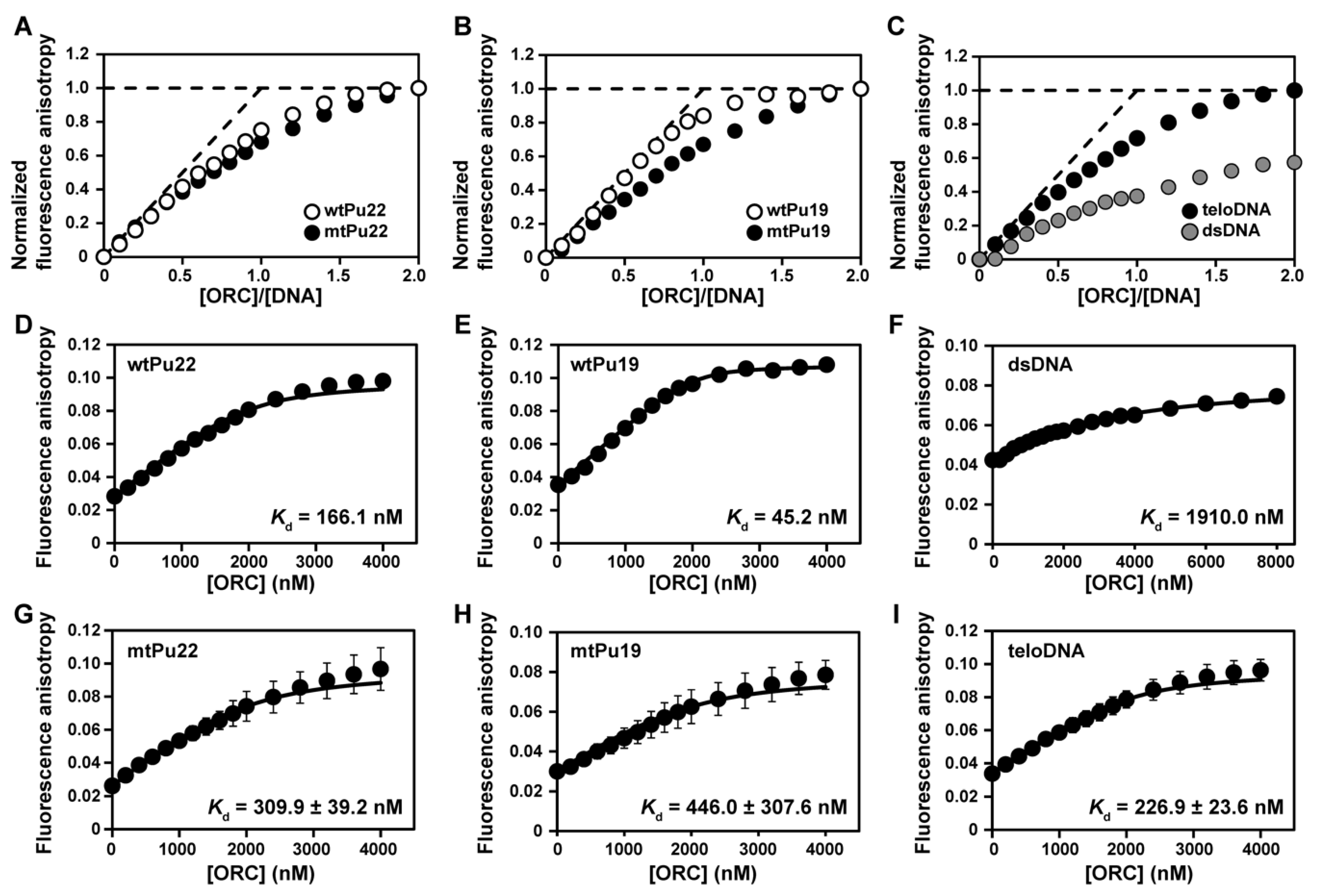
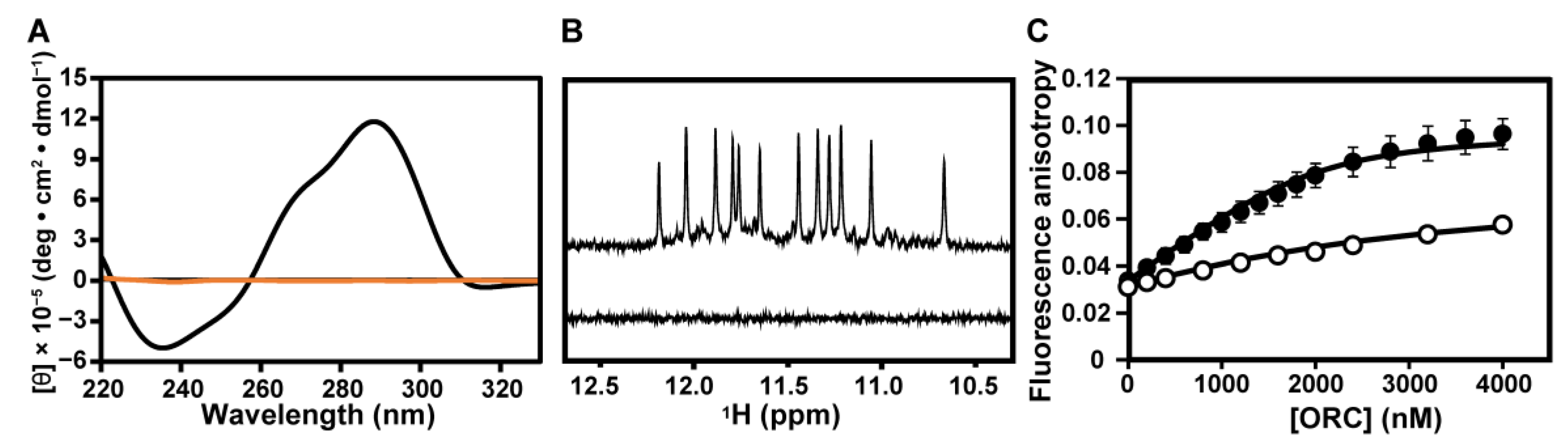
2.1. Binding of hORC1413–511 to DNAs Having a G4 Structure
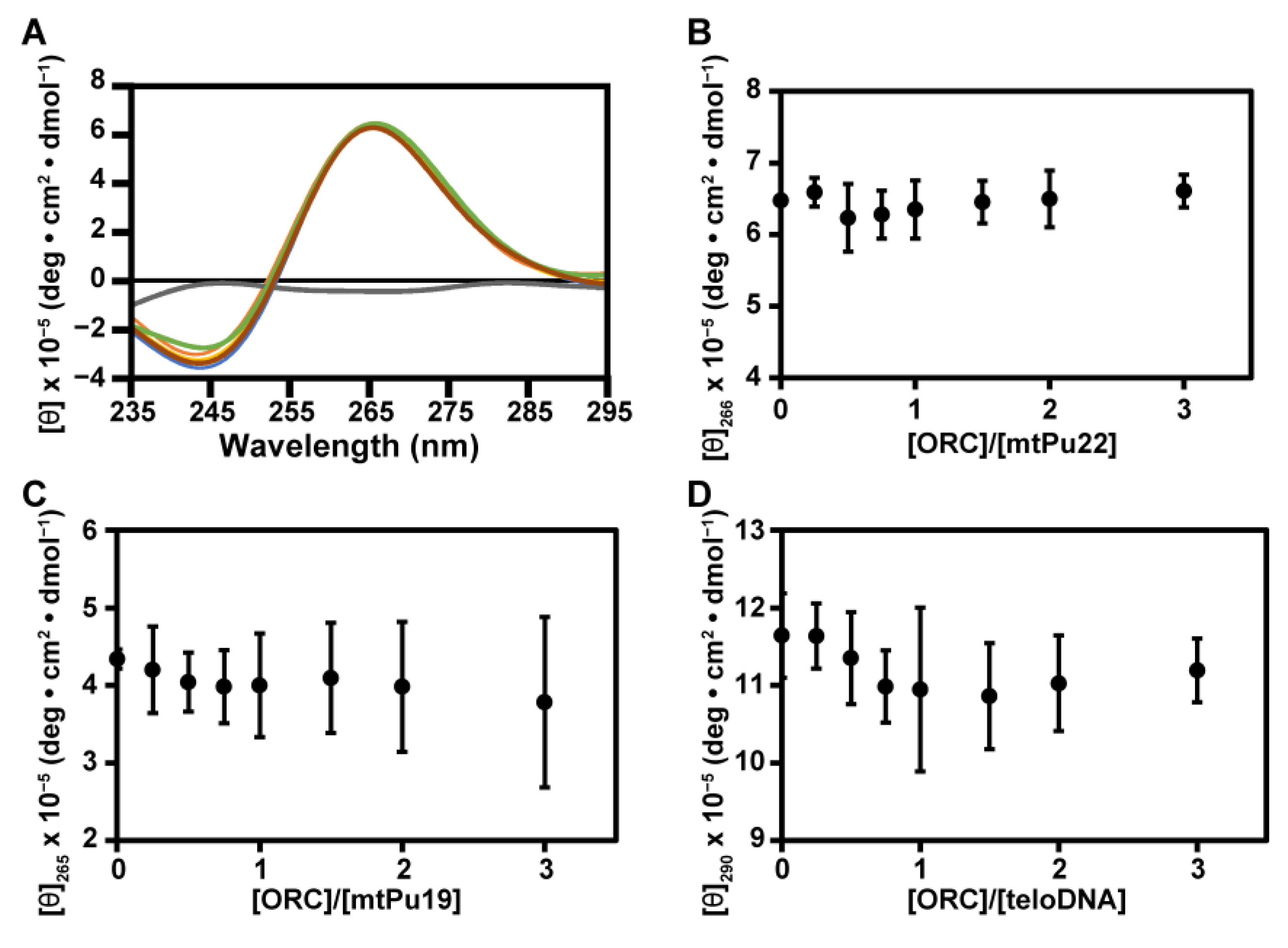
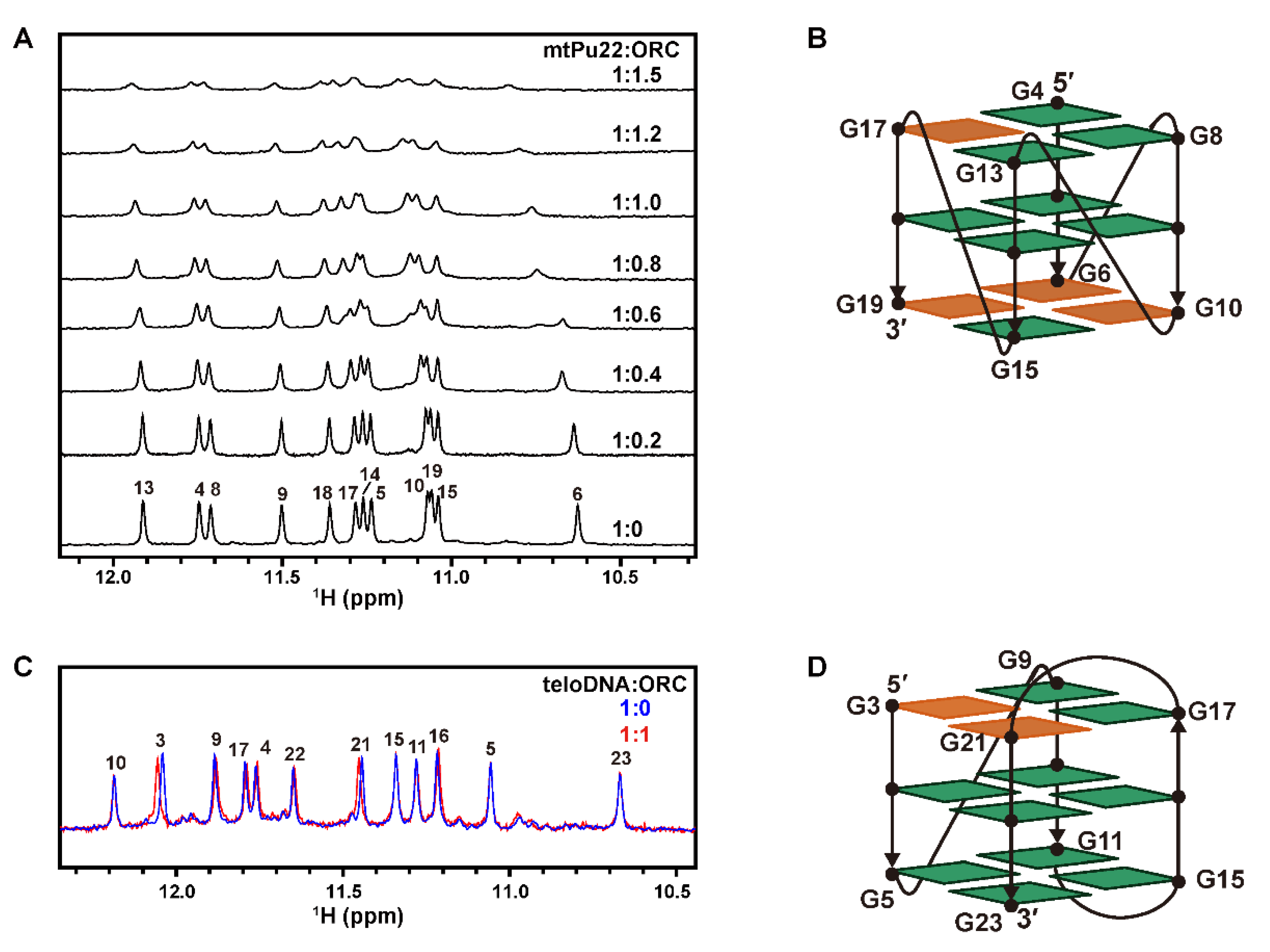
2.2. G4 Structures Are Retained in G-Rich DNAs in Complexes with hORC1413–511
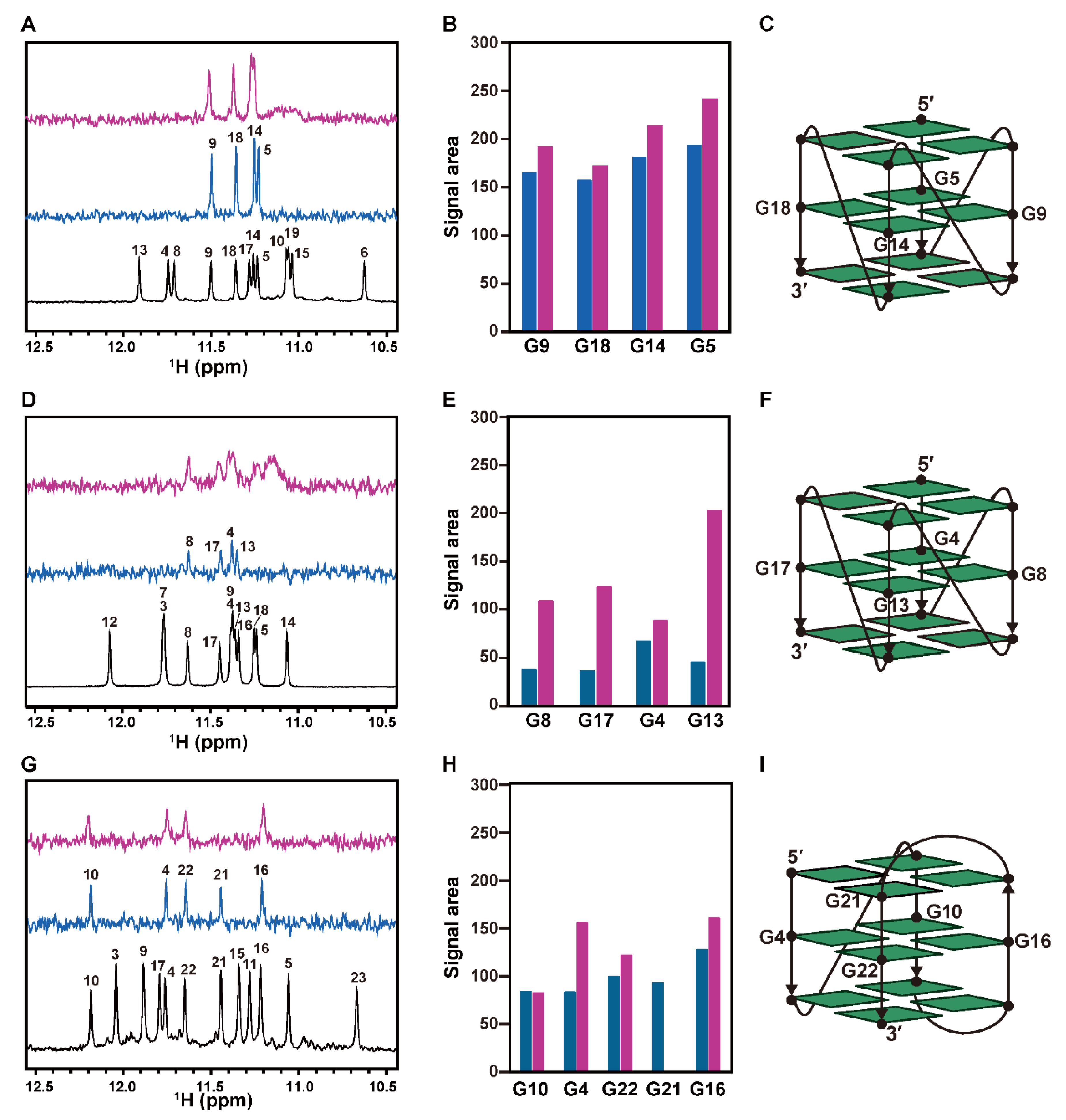
2.3. Interaction of hORC1413–511 with External G-Tetrad Planes of the G4 Structure
3. Discussion
4. Materials and Methods
4.1. Protein Expression and Purification
4.2. Preparation of DNAs
4.3. Fluorescence Anisotropy (FA) Measurement
4.4. CD Spectroscopy
4.5. NMR Spectroscopy
4.6. NMR-Detected Hydrogen–Deuterium Exchange (H/D Exchange) Measurement
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yuan, Z.; Li, H. Molecular mechanisms of eukaryotic origin initiation, replication fork progression, and chromatin maintenance. Biochem. J. 2020, 477, 3499–3525. [Google Scholar] [CrossRef] [PubMed]
- Bell, S.P.; Stillman, B. ATP-dependent recognition of eukaryotic origins of DNA replication by a multiprotein complex. Nature 1992, 357, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Rao, H.; Stillman, B. The origin recognition complex interacts with a bipartite DNA binding site within yeast replicators. Proc. Natl. Acad. Sci. USA 1995, 92, 2224–2228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dellino, G.I.; Cittaro, D.; Piccioni, R.; Luzi, L.; Banfi, S.; Segalla, S.; Cesaroni, M.; Mendoza-Maldonado, R.; Giacca, M.; Pelicci, P.G. Genome-wide mapping of human DNA-replication origins: Levels of transcription at ORC1 sites regulate origin selection and replication timing. Genome Res. 2013, 1, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Miotto, B.; Ji, Z.; Struhl, K. Selectivity of ORC binding sites and the relation to replication timing, fragile sites, and deletions in cancers. Proc. Natl Acad. Sci. USA 2016, 113, E4810–E4819. [Google Scholar] [CrossRef] [Green Version]
- Méchali, M. Eukaryotic DNA replication origins: Many choices for appropriate answers. Nat. Rev. Mol. Cell Biol. 2010, 11, 728–738. [Google Scholar] [CrossRef] [PubMed]
- Ganier, O.; Prorok, P.; Akerman, I.; Méchali, M. Metazoan DNA replication origins. Curr. Opin. Cell Biol. 2019, 58, 134–141. [Google Scholar] [CrossRef]
- Sun, J.; Kawakami, H.; Zech, J.; Speck, C.; Stillman, B.; Li, H. Cdc6-induced conformational changes in ORC bound to origin DNA revealed by cryo-electron microscopy. Structure 2012, 20, 534–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, Z.; Riera, A.; Bai, L.; Sun, J.; Nandi, S.; Spanos, C.; Chen, A.Z.; Barbon, M.; Rappsilber, J.; Stillman, B.; et al. Structural basis of Mcm2–7 replicative helicase loading by ORC–Cdc6 and Cdt1. Nat. Struct. Mol. Biol. 2017, 3, 316–324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, N.; Lam, W.H.; Zhai, Y.; Cheng, J.; Cheng, E.; Zhao, Y.; Gao, N.; Tye, B.K. Structure of the origin recognition complex bound to DNA replication origin. Nature 2018, 559, 217–222. [Google Scholar] [CrossRef]
- Bleichert, F.; Botchan, M.R.; Berger, J.M. Crystal structure of the eukaryotic origin recognition complex. Nature 2015, 519, 321–326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, J.M.; Bleichert, F. Structural mechanism for replication origin binding and remodeling by a metazoan origin recognition complex and its co-loader Cdc6. Nat. Commun. 2020, 11, 4263. [Google Scholar] [CrossRef] [PubMed]
- Tocilj, A.; On, K.F.; Yuan, Z.; Jingchuan Sun, J.; Elkayam, E.; Li, H.; Stillman, B.; Joshua-Tor, L. Structure of the active form of human origin recognition complex and its ATPase motor module. eLife 2017, 6, e20818. [Google Scholar] [CrossRef]
- Jaremko, M.J.; On, K.F.; Thomas, D.R.; Stillman, B.; Joshua-Tor, L. The dynamic nature of the human origin recognition complex revealed through five cryoEM structures. eLife 2020, 9, e58622. [Google Scholar] [CrossRef]
- Xu, N.; You, Y.; Liu, C.; Balasov, M.; Lun, L.T.; Geng, Y.; Fung, C.P.; Miao, H.; Tian, H.; Choy, T.T.; et al. Structural basis of DNA replication origin recognition by human Orc6 protein binding with DNA. Nucleic Acids Res. 2020, 48, 11146–11161. [Google Scholar] [CrossRef]
- Cheng, J.; Li, N.; Wang, X.; Hu, J.; Zhai, Y.; Gao, N. Structural insight into the assembly and conformational activation of human origin recognition complex. Cell Discov. 2020, 6, 88. [Google Scholar] [CrossRef]
- Cayrou, C.; Coulombe, P.; Vigneron, A.; Stanojcic, S.; Ganier, O.; Peiffer, I.; Rivals, E.; Puy, A.; Laurent-Chabalier, S.; Desprat, R.; et al. Genome-scale analysis of metazoan replication origins reveals their organization in specific but flexible sites defined by conserved features. Genome Res. 2011, 21, 1438–1449. [Google Scholar] [CrossRef] [Green Version]
- Cayrou, C.; Coulombe, P.; Puy, A.; Rialle, S.; Kaplan, N.; Segal, E.; Méchali, M. New insights into replication origin characteristics in metazoans. Cell Cycle 2012, 11, 658–667. [Google Scholar] [CrossRef] [Green Version]
- Besnard, E.; Babled, A.; Lapasset, L.; Milhavet, O.; Parrinello, H.; Dantec, C.; Marin, J.M.; Lemaitre, J.M. Unraveling cell type-specific and reprogrammable human replication origin signatures associated with G-quadruplex consensus motifs. Nat. Struct. Mol. Biol. 2012, 8, 837–844. [Google Scholar] [CrossRef]
- Sugimoto, N.; Maehara, K.; Yoshida, K.; Ohkawa, Y.; Fujita, M. Genome-wide analysis of the spatiotemporal regulation of firing and dormant replication origins in human cells. Nucleic Acids Res. 2018, 46, 6683–6696. [Google Scholar] [CrossRef] [PubMed]
- Valton, A.L.; Hassan-Zadeh, V.; Lema, I.; Boggetto, N.; Alberti, P.; Saintomé, C.; Riou, J.F.; Prioleau, M.N. G4 motifs affect origin positioning and efficiency in two vertebrate replicators. EMBO J. 2014, 33, 732–746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prorok, P.; Artufel, M.; Aze, A.; Coulombe, P.; Peiffer, I.; Lacroix, L.; Guédin, A.; Mergny, J.L.; Damaschke, J.; Schepers, A.; et al. Involvement of G-quadruplex regions in mammalian replication origin activity. Nat. Commun. 2019, 10, 3274. [Google Scholar] [CrossRef] [Green Version]
- McWhinney, C.; Leffak, M. Autonomous replication of a DNA fragment containing the chromosomal replication origin of the human c-myc gene. Nucleic Acids Res. 1990, 18, 1233–1242. [Google Scholar] [CrossRef] [Green Version]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Hänsel-Hertsch, R.; Beraldi, D.; Lensing, S.V.; Marsico, G.; Zyner, K.; Parry, A.; Antonio, M.D.; Pike, J.; Kimura, H.; Narita, M.; et al. G-quadruplex structures mark human regulatory chromatin. Nat. Genet. 2016, 48, 1267–1272. [Google Scholar] [CrossRef] [Green Version]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Hoshina, S.; Yura, K.; Teranishi, H.; Kiyasu, N.; Tominaga, A.; Kadoma, H.; Nakatsuka, A.; Kunichika, T.; Obuse, C.; Waga, S. Human origin recognition complex binds preferentially to G-quadruplex-preferable RNA and single-stranded DNA. J. Biol. Chem. 2013, 288, 30161–30171. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, C.E.; Gokhale, V.; Yang, D.; Hurley, L.H. G-Quadruplex-Mediated Transcriptional Regulation of PDGFR-β. Top. Curr. Chem. 2013, 330, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Phan, A.T.; Modi, Y.S.; Patel, D.J. Propeller-type parallel-stranded G-quadruplexes in the human c-myc promoter. J. Am. Chem. Soc. 2004, 126, 8710–8716. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Chen, D.; Dai, J.; Jones, R.A.; Yang, D. Solution structure of the biologically relevant G-quadruplex element in the human c-MYC promoter. Implications for G-quadruplex stabilization. Biochemistry 2005, 44, 2048–2058. [Google Scholar] [CrossRef]
- Mathad, R.I.; Hatzakis, E.; Dai, J.; Yang, D. c-MYC promoter G-quadruplex formed at the 5′-end of NHE III1 element: Insights into biological relevance and parallel-stranded G-quadruplex stability. Nucleic Acids Res. 2011, 39, 9023–9033. [Google Scholar] [CrossRef]
- Luu, K.N.; Phan, A.T.; Kuryavyi, V.; Lacroix, L.; Patel, D.J. Structure of the human telomere in K+ solution: An intramolecular (3 + 1) G-quadruplex scaffold. J. Am. Chem. Soc. 2006, 128, 9963–9970. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharyya, D.; Mirihana Arachchilage, G.; Basu, S. Metal Cations in G-Quadruplex folding and stability. Front. Chem. 2016, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human telomeric sequence forms a hybrid-type intramolecular G-quadruplex structure with mixed parallel/antiparallel strands in potassium solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef] [Green Version]
- Adrian, M.; Heddi, B.; Phan, A.T. NMR spectroscopy of G-quadruplexes. Methods 2012, 57, 11–24. [Google Scholar] [CrossRef]
- Kondo, K.; Mashima, T.; Oyoshi, T.; Yagi, R.; Kurokawa, R.; Kobayashi, N.; Nagata, T.; Katahira, M. Plastic roles of phenylalanine and tyrosine residues of an RGG motif of TLS/FUS in binary and ternary complex formation with the G-quadruplex structures of telomeric DNA and TERRA. Sci. Rep. 2018, 8, 2864. [Google Scholar] [CrossRef]
- Enokizono, Y.; Konishi, Y.; Nagata, K.; Ouhashi, K.; Uesugi, S.; Ishikawa, F.; Katahira, M. Structure of hnRNP D complexed with single-stranded telomere DNA and unfolding of the quadruplex by heterogeneous nuclear ribonucleoprotein D. J. Biol. Chem. 2005, 280, 18862–18870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brázda, V.; Hároníková, L.; Liao, J.C.C.; Fojta, M. DNA and RNA quadruplex-binding proteins. Int. J. Mol. Sci. 2014, 15, 17493–17517. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, M.W.; Bell, M.; Mir, M.; Kao, J.A.; Darzacq, X.; Botchan, M.R.; Berger, J.M. A new class of disordered elements controls DNA replication through initiator self-assembly. eLife 2019, 8, e48562. [Google Scholar] [CrossRef]
- Kuipers, B.J.H.; Gruppen, H. Prediction of molar extinction coefficients of proteins and peptides using UV absorption of the constituent amino acids at 214 nm to enable quantitative reverse phase high-performance liquid chromatography-mass spectrometry analysis. J. Agric. Food Chem. 2007, 55, 5445–5451. [Google Scholar] [CrossRef] [PubMed]
- Favicchio, R.; Dragan, A.I.; Kneale, G.G.; Read, C.M. Fluorescence spectroscopy and anisotropy in the analysis of DNA-protein interactions. Methods Mol. Biol. 2009, 543, 589–611. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence | Structure |
|---|---|---|
| mtPu22 | 5′-TGAGGGTGGGTAGGGTGGGTAA-3′ | Parallel G4 |
| mtPu19 | 5′-TAGGGAGGGTAGGGAGGGT-3′ | Parallel G4 |
| teloDNA | 5′-TTGGGTTAGGGTTAGGGTTAGGGA-3′ | (3+1)-type G4 |
| wtPu22 | 5′-TGAGGGTGGGGAGGGTGGGGAA-3′ | G4 |
| wtPu19 | 5′-TGGGGAGGGTGGGGAGGGT-3′ | G4 |
| dsDNA_1 | 5′-AAATTTAAAAAAAAAAATAATT-3′ | Duplex |
| dsDNA_2 | 5′-AATTATTTTTTTTTTTAAATTT-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eladl, A.; Yamaoki, Y.; Hoshina, S.; Horinouchi, H.; Kondo, K.; Waga, S.; Nagata, T.; Katahira, M. Investigation of the Interaction of Human Origin Recognition Complex Subunit 1 with G-Quadruplex DNAs of Human c-myc Promoter and Telomere Regions. Int. J. Mol. Sci. 2021, 22, 3481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073481
Eladl A, Yamaoki Y, Hoshina S, Horinouchi H, Kondo K, Waga S, Nagata T, Katahira M. Investigation of the Interaction of Human Origin Recognition Complex Subunit 1 with G-Quadruplex DNAs of Human c-myc Promoter and Telomere Regions. International Journal of Molecular Sciences. 2021; 22(7):3481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073481
Chicago/Turabian StyleEladl, Afaf, Yudai Yamaoki, Shoko Hoshina, Haruka Horinouchi, Keiko Kondo, Shou Waga, Takashi Nagata, and Masato Katahira. 2021. "Investigation of the Interaction of Human Origin Recognition Complex Subunit 1 with G-Quadruplex DNAs of Human c-myc Promoter and Telomere Regions" International Journal of Molecular Sciences 22, no. 7: 3481. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073481