
Vitamin D Deficiency Induces Chronic Pain and Microglial Phenotypic Changes in Mice

 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Vitamin D Deficiency Promotes Primary Microglia Cells Activation and Proliferation
2.2. Vitamin D Deficiency Induces ROS Generation in Primary Microglia Cells
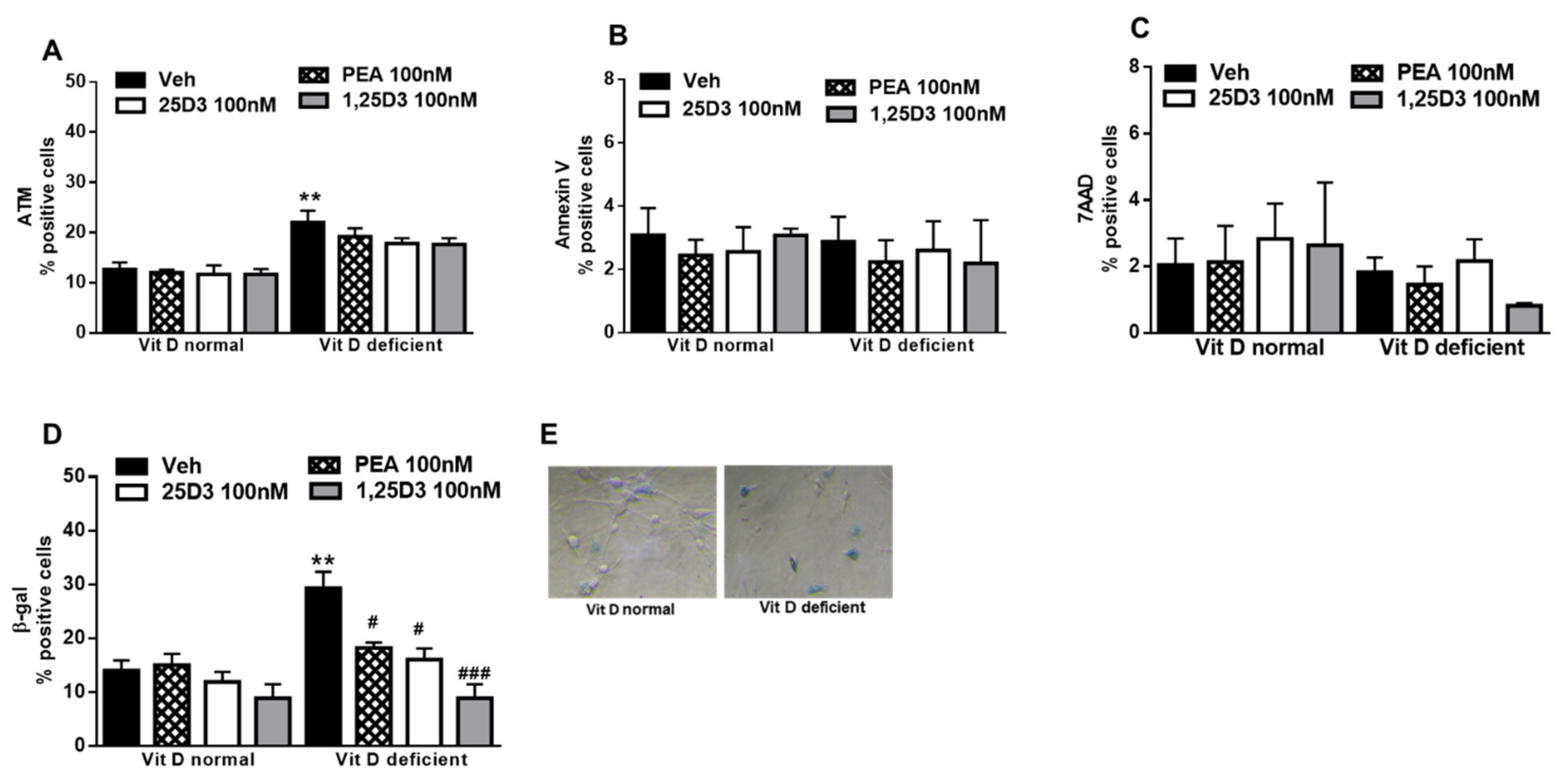
2.3. Vitamin D Deficiency Increases β-Gal Activity in Primary Microglia Cells
2.4. Vitamin D Deficiency Induces Pain Behavior in Adult Male and Female Mice
2.5. Vitamin D Deficiency Induces Microglial Morphological Changes in Adult Mice
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Primary Microglial Cultures
4.3. Immunocytochemistry
4.4. Measurement of Primary Microglial Cells Proliferation
4.5. Cell Cycle Analysis of Primary Microglial Cultures
4.6. Nexin V Assay in Primary Microglial Cultures
4.7. DCF-DA Assay in Primary Microglial Cultures
4.8. D-ROM Test
4.9. In Situ Senescence-Associated Beta-Galactosidase Assay in Primary Microglial Cultures
4.10. Serum Vitamin D Measurements
4.11. Behavior in Adult Mice
4.12. Ex Vivo Immunohistochemistry in Adult Mice
5. Statistical Analysis
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Eyles, D.W.; Smith, S.; Kinobe, R.; Hewison, M.; McGrath, J.J. Distribution of the Vitamin D receptor and 1α-hydroxylase in human brain. J. Chem. Neuroanat. 2005, 29, 21–30. [Google Scholar] [CrossRef] [PubMed]
- Langub, M.; Herman, J.; Malluche, H.; Koszewski, N. Evidence of functional vitamin D receptors in rat hippocampus. Neuroscience 2001, 104, 49–56. [Google Scholar] [CrossRef]
- Malcangio, M. Translational value of preclinical models for rheumatoid arthritis pain. Pain 2020, 161, 1399–1400. [Google Scholar] [CrossRef] [PubMed]
- Boontanrart, M.; Hall, S.D.; Spanier, J.A.; Hayes, C.E.; Olson, J.K. Vitamin D3 alters microglia immune activation by an IL-10 dependent SOCS3 mechanism. J. Neuroimmunol. 2016, 292, 126–136. [Google Scholar] [CrossRef]
- Landel, V.; Stephan, D.; Cui, X.; Eyles, D.; Feron, F. Differential expression of vitamin D-associated enzymes and receptors in brain cell subtypes. J. Steroid Biochem. Mol. Biol. 2018, 177, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Akyuz, G.; Sanal-Toprak, C.; Yagci, I.; Giray, E.; Kuru-Bektasoglu, P. The effect of vitamin D supplementation on pain, quality of life, and nerve conduction studies in women with chronic widespread pain. Int. J. Rehabil. Res. 2017, 40, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Zadro, J.R.; Shirley, D.; Ferreira, M.; Silva, A.P.C.; Lamb, S.E.; Cooper, C.; Ferreira, P.H. Is Vitamin D Supplementation Effective for Low Back Pain? A Systematic Review and Meta-Analysis. Pain Physician 2018, 21, 121–145. [Google Scholar] [CrossRef]
- Helde-Frankling, M.; Björkhem-Bergman, L. Vitamin D in Pain Management. Int. J. Mol. Sci. 2017, 18, 2170. [Google Scholar] [CrossRef] [Green Version]
- Guida, F.; Boccella, S.; Belardo, C.; Iannotta, M.; Piscitelli, F.; De Filippis, F.; Paino, S.; Ricciardi, F.; Siniscalco, D.; Marabese, I.; et al. Altered gut microbiota and endocannabinoid system tone in vitamin D deficiency-mediated chronic pain. Brain Behav. Immun. 2020, 85, 128–141. [Google Scholar] [CrossRef]
- Samuel, S.; Sitrin, M.D. Vitamin D’s role in cell proliferation and differentiation. Nutr. Rev. 2008, 66 (Suppl. 2), S116–S124. [Google Scholar] [CrossRef]
- Umar, M.; Sastry, K.S.; Chouchane, A.I. Role of Vitamin D Beyond the Skeletal Function: A Review of the Molecular and Clinical Studies. Int. J. Mol. Sci. 2018, 19, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Catale, C.; Gironda, S.; Iacono, L.L.; Carola, V. Microglial Function in the Effects of Early-Life Stress on Brain and Behavioral Development. J. Clin. Med. 2020, 9, 468. [Google Scholar] [CrossRef] [Green Version]
- Bae, Y.S.; Lee, J.H.; Choi, S.H.; Kim, S.; Almazan, F.; Witztum, J.L.; Miller, Y.I. Macrophages generate reactive oxygen species in response to minimally oxidized low-density lipoprotein: Toll-like receptor 4–and spleen tyrosine kinase–dependent activation of NADPH oxidase 2. Circ. Res. 2009, 104, 210–218. [Google Scholar] [CrossRef] [Green Version]
- Djukic, M.; Sostmann, N.; Bertsch, T.; Mecke, M.; Nessler, S.; Manig, A.; Hanisch, U.-K.; Triebel, J.; Bollheimer, L.C.; Sieber, C.; et al. Vitamin D deficiency decreases survival of bacterial meningoencephalitis in mice. J. Neuroinflamm. 2015, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Halicka, H.D.; Zhao, H.; Li, J.; Traganos, F.; Studzinski, G.P.; Darzynkiewicz, Z. Attenuation of constitutive DNA damage signaling by 1,25-dihydroxyvitamin D3. Aging 2012, 4, 270–278. [Google Scholar] [CrossRef] [Green Version]
- Wood, M.A.; Cavender, J.F. Research Article: Beta-galactosidase staining as a marker of cells enduring stress. BIOS 2004, 75, 139–146. [Google Scholar] [CrossRef]
- Boccella, S.; Cristiano, C.; Romano, R.; Iannotta, M.; Belardo, C.; Farina, A.; Guida, F.; Piscitelli, F.; Palazzo, E.; Mazzitelli, M.; et al. Ultra-micronized palmitoylethanolamide rescues the cognitive decline-associated loss of neural plasticity in the neuropathic mouse entorhinal cortex-dentate gyrus pathway. Neurobiol. Dis. 2019, 121, 106–119. [Google Scholar] [CrossRef]
- Guida, F.; Luongo, L.; Boccella, S.; Giordano, M.E.; Romano, R.; Bellini, G.; Manzo, I.; Furiano, A.; Rizzo, A.; Imperatore, R.; et al. Palmitoylethanolamide induces microglia changes associated with increased migration and phagocytic activity: Involvement of the CB2 receptor. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Guida, F.; Luongo, L.; Marmo, F.; Romano, R.; Iannotta, M.; Napolitano, F.; Belardo, C.; Marabese, I.; Aniello, A.D.; De Gregorio, D.; et al. Palmitoylethanolamide reduces pain-related behaviors and restores glutamatergic synapses homeostasis in the medial prefrontal cortex of neuropathic mice. Mol. Brain 2015, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Luongo, L.; Guida, F.; Boccella, S.; Bellini, G.; Gatta, L.; Rossi, F.; De Novellis, V.; Maione, S. Palmitoylethanolamide reduces formalin-induced neuropathic-like behaviour through spinal glial/microglial phenotypical changes in mice. CNS Neurol. Disord. Drug Targets 2013, 12, 45–54. [Google Scholar] [CrossRef]
- Sertznig, P.; Reichrath, J. Peroxisome proliferator-activated receptors (PPARs) in dermatology: Challenge and promise. Derm. Endocrinol. 2011, 3, 130–135. [Google Scholar] [CrossRef]
- Barcelon, E.E.; Cho, W.-H.; Jun, S.B.; Lee, S.J. Brain Microglial Activation in Chronic Pain-Associated Affective Disorder. Front. Neurosci. 2019, 13, 213. [Google Scholar] [CrossRef] [Green Version]
- Streit, W.J.; Braak, H.; Xue, Q.-S.; Bechmann, I. Dystrophic (senescent) rather than activated microglial cells are associated with tau pathology and likely precede neurodegeneration in Alzheimer’s disease. Acta Neuropathol. 2009, 118, 475–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Streit, W.J.; Khoshbouei, H.; Bechmann, I. Dystrophic microglia in late-onset Alzheimer’s disease. Glia 2020, 68, 845–854. [Google Scholar] [CrossRef]
- Day, C.; Rees, K.; Weich, S.; Clarke, A.; Stranges, S. OP35 Vitamin D to reduce depressive symptoms: A systematic review. J. Epidemiol. Community Health 2011, 65 (Suppl II), A16. [Google Scholar] [CrossRef] [Green Version]
- Coraggio, V.; Guida, F.; Boccella, S.; Scafuro, M.; Paino, S.; Romano, D.; Maione, S.; Luongo, L. Neuroimmune-Driven Neuropathic Pain Establishment: A Focus on Gender Differences. Int. J. Mol. Sci. 2018, 19, 281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolders, J.; Schuurman, K.G.; Van Strien, M.E.; Melief, J.; Hendrickx, D.; Hol, E.M.; Van Eden, C.; Luchetti, S.; Huitinga, I. Expression of Vitamin D Receptor and Metabolizing Enzymes in Multiple Sclerosis—Affected Brain Tissue. J. Neuropathol. Exp. Neurol. 2013, 72, 91–105. [Google Scholar] [CrossRef] [Green Version]
- Rinaldi, B.; Guida, F.; Furiano, A.; Donniacuo, M.; Luongo, L.; Gritti, G.; Urbanek, K.; Messina, G.; Maione, S.; Rossi, F.; et al. Effect of Prolonged Moderate Exercise on the Changes of Nonneuronal Cells in Early Myocardial Infarction. Neural Plast. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Yin, Z.; Raj, D.D.; Schaafsma, W.; Van Der Heijden, R.A.; Kooistra, S.M.; Reijne, A.C.; Zhang, X.; Moser, J.; Brouwer, N.; Heeringa, P.; et al. Low-Fat Diet with Caloric Restriction Reduces White Matter Microglia Activation During Aging. Front. Mol. Neurosci. 2018, 11, 65. [Google Scholar] [CrossRef]
- Latimer, C.S.; Brewer, L.D.; Searcy, J.L.; Chen, K.-C.; Popović, J.; Kraner, S.D.; Thibault, O.; Blalock, E.M.; Landfield, P.W.; Porter, N.M. Vitamin D prevents cognitive decline and enhances hippocampal synaptic function in aging rats. Proc. Natl. Acad. Sci. USA 2014, 111, E4359–E4366. [Google Scholar] [CrossRef] [Green Version]
- Luongo, L.; Guida, F.; Imperatore, R.; Napolitano, F.; Gatta, L.; Cristino, L.; Giordano, C.; Siniscalco, D.; Di Marzo, V.; Bellini, G.; et al. The A1 adenosine receptor as a new player in microglia physiology. Glia 2013, 62, 122–132. [Google Scholar] [CrossRef]
- Alessio, N.; Capasso, S.; Ferone, A.; Di Bernardo, G.; Cipollaro, M.; Casale, F.; Peluso, G.; Giordano, A.; Galderisi, U. Misidentified Human Gene Functions with Mouse Models: The Case of the Retinoblastoma Gene Family in Senescence. Neoplasia 2017, 19, 781–790. [Google Scholar] [CrossRef] [PubMed]
- Chaplan, S.; Bach, F.; Pogrel, J.; Chung, J.; Yaksh, T. Quantitative assessment of tactile allodynia in the rat paw. J. Neurosci. Methods 1994, 53, 55–63. [Google Scholar] [CrossRef]
- Cristino, L.; Luongo, L.; Squillace, M.; Paolone, G.; Mango, D.; Piccinin, S.; Zianni, E.; Imperatore, R.; Iannotta, M.; Longo, F.; et al. d-Aspartate oxidase influences glutamatergic system homeostasis in mammalian brain. Neurobiol. Aging 2015, 36, 1890–1902. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alessio, N.; Belardo, C.; Trotta, M.C.; Paino, S.; Boccella, S.; Gargano, F.; Pieretti, G.; Ricciardi, F.; Marabese, I.; Luongo, L.; et al. Vitamin D Deficiency Induces Chronic Pain and Microglial Phenotypic Changes in Mice. Int. J. Mol. Sci. 2021, 22, 3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073604
Alessio N, Belardo C, Trotta MC, Paino S, Boccella S, Gargano F, Pieretti G, Ricciardi F, Marabese I, Luongo L, et al. Vitamin D Deficiency Induces Chronic Pain and Microglial Phenotypic Changes in Mice. International Journal of Molecular Sciences. 2021; 22(7):3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073604
Chicago/Turabian StyleAlessio, Nicola, Carmela Belardo, Maria Consiglia Trotta, Salvatore Paino, Serena Boccella, Francesca Gargano, Gorizio Pieretti, Flavia Ricciardi, Ida Marabese, Livio Luongo, and et al. 2021. "Vitamin D Deficiency Induces Chronic Pain and Microglial Phenotypic Changes in Mice" International Journal of Molecular Sciences 22, no. 7: 3604. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22073604