
Transcriptomic Profiles of CD47 in Breast Tumors Predict Outcome and Are Associated with Immune Activation

 ,
,  , ,
, ,  and
and 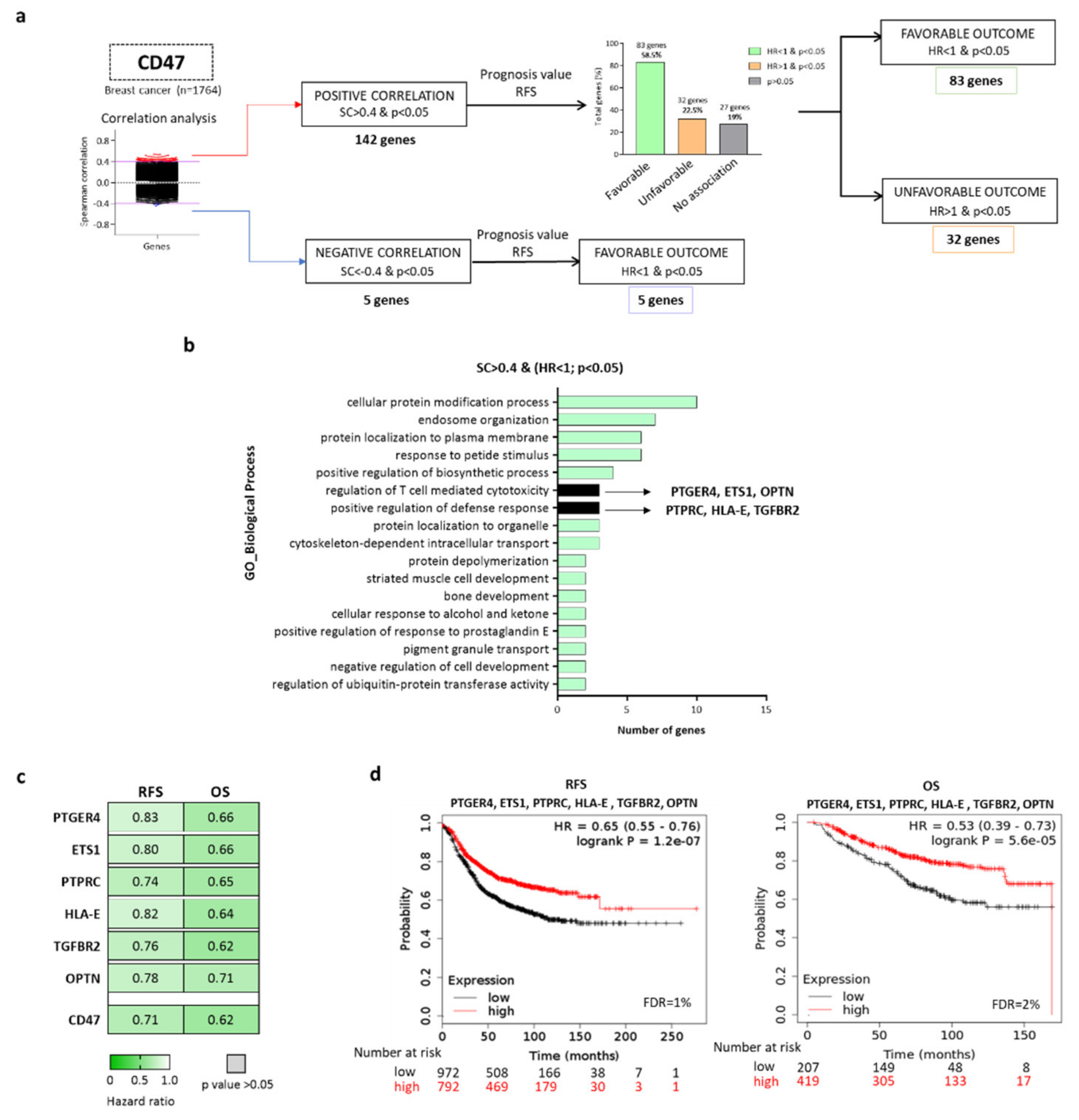{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Identification of Genes Expressed in Breast Tumors with High Expression of CD47
2.2. CD47-Immune Signature is Associated with Favorable Prognosis in Breast Cancer, Especially for the BASAL-Like and HER2+ Subtypes
2.3. CD47-Immune Signature Correlated with the Presence of Immune Infiltrates in Basal-Like and HER2+ Breast Tumors
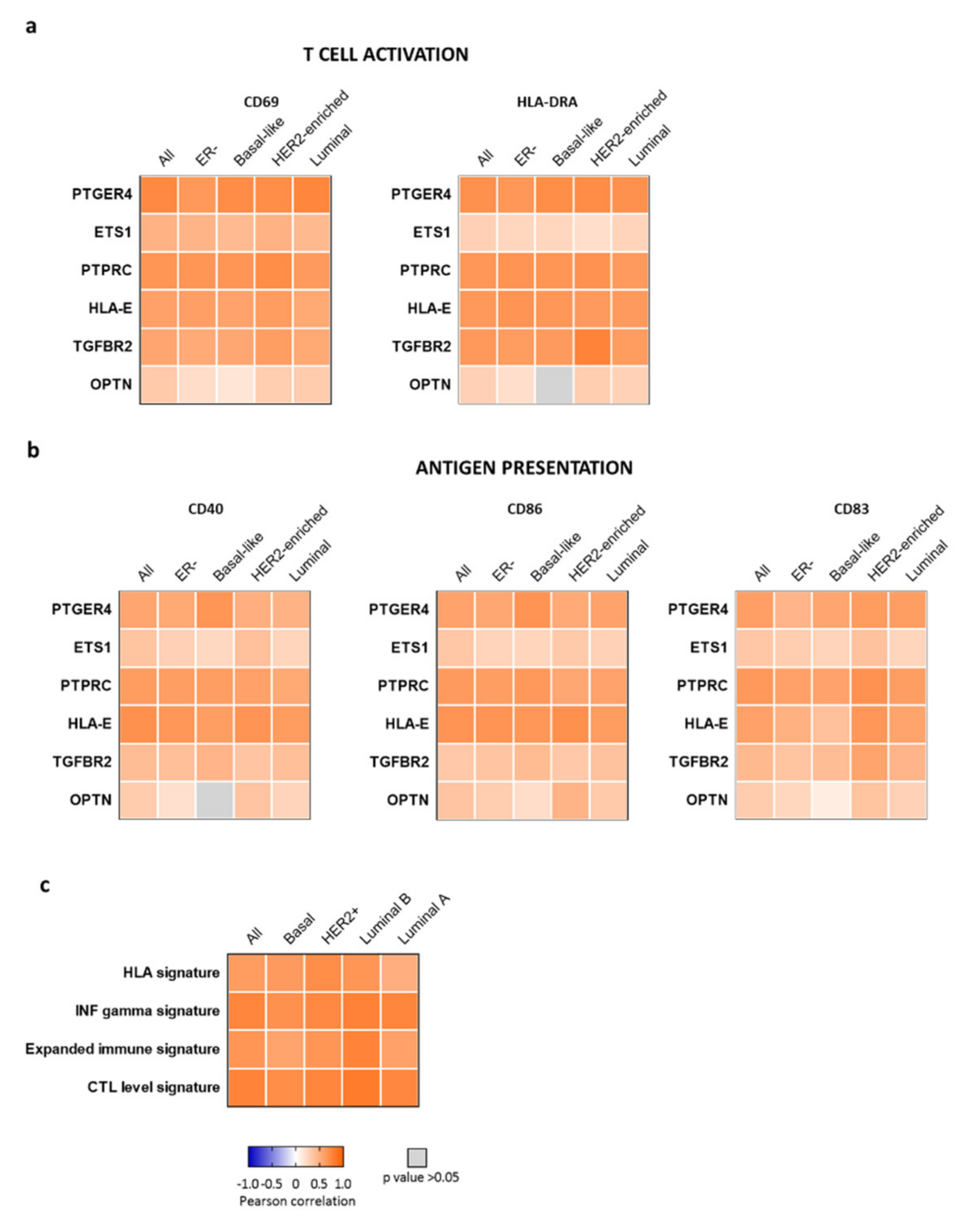
2.4. CD47-Immune Signature is Associated with Markers of T Cell Activation and Antigen Presentation
2.5. CD47-Immune Signature Correlated with Gene Signatures of T Cell Activation
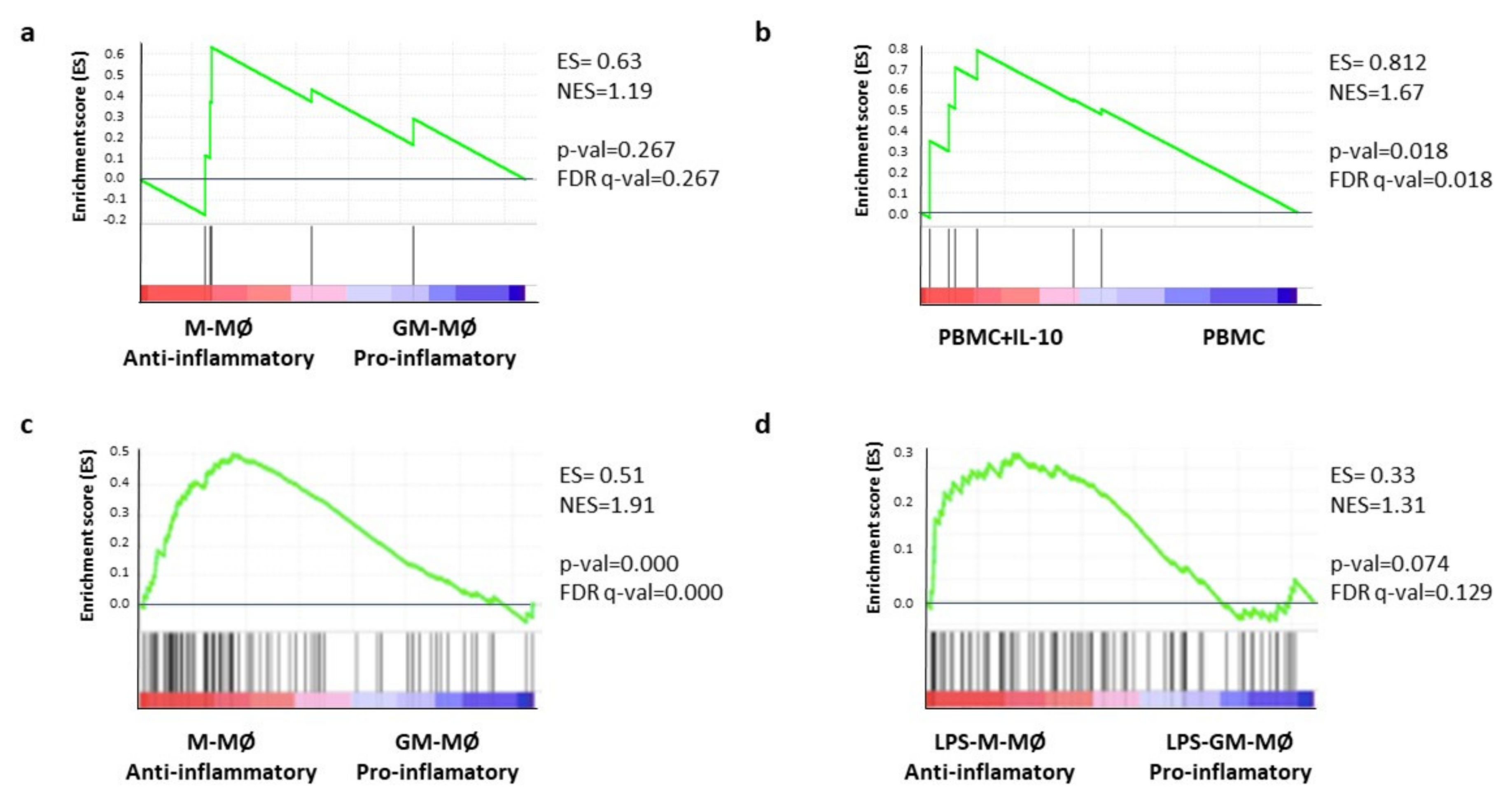
2.6. Gene-Set Enrichment Analysis (GSEA) Confirm the Association of the CD47-Immune Signature with Pro-Tumoral Macrophages
3. Discussion
4. Materials and Methods
4.1. Exploratory Cohort
4.2. Validation Cohort
4.3. Gene Function Analysis
4.4. Protein Functional Analyses
4.5. Association between Tumor Immune Infiltrates and Gene Expression
4.6. Correlation between Gene Expression and T Cell Activation and Antigen Presentation
4.7. Correlation between Gene Expression and Macrophage Signatures
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hargadon, K.M.; Johnson, C.E.; Williams, C.J. Immune checkpoint blockade therapy for cancer: An overview of FDA-approved immune checkpoint inhibitors. Int. Immunopharmacol. 2018, 62, 29–39. [Google Scholar] [CrossRef]
- Luke, J.J.; Flaherty, K.T.; Ribas, A.; Long, G.V. Targeted agents and immunotherapies: Optimizing outcomes in melanoma. Nat. Rev. Clin. Oncol. 2017, 14, 463–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldman, A.D.; Fritz, J.M.; Lenardo, M.J. A guide to cancer immunotherapy: From T cell basic science to clinical practice. Nat. Rev. Immunol. 2020, 20, 651–668. [Google Scholar] [CrossRef]
- Dong, Y.; Sun, Q.; Zhang, X. PD-1 and its ligands are important immune checkpoints in cancer. Oncotarget 2017, 8, 2171–2186. [Google Scholar] [CrossRef] [Green Version]
- Ribas, A.; Wolchok, J.D. Cancer immunotherapy using checkpoint blockade. Science 2018, 359, 1350–1355. [Google Scholar] [CrossRef] [Green Version]
- Gettinger, S.N.; Horn, L.; Gandhi, L.; Spigel, D.R.; Antonia, S.J.; Rizvi, N.A.; Powderly, J.D.; Heist, R.S.; Carvajal, R.D.; Jackman, D.M.; et al. Overall Survival and Long-Term Safety of Nivolumab (Anti–Programmed Death 1 Antibody, BMS-936558, ONO-4538) in Patients with Previously Treated Advanced Non–Small-Cell Lung Cancer. J. Clin. Oncol. 2015, 33, 2004–2012. [Google Scholar] [CrossRef]
- Zappasodi, R.; Merghoub, T.; Wolchok, J.D. Emerging Concepts for Immune Checkpoint Blockade-Based Combination Therapies. Cancer Cell 2018, 33, 581–598. [Google Scholar] [CrossRef] [Green Version]
- Noblejas-López, M.d.M.; Nieto-Jiménez, C.; Morcillo García, S.; Pérez-Peña, J.; Nuncia-Cantarero, M.; Andrés-Pretel, F. Expression of MHC class I, HLA-A and HLA-B identifies immune-activated breast tumors with favorable outcome. Oncoimmunology 2019, 8, e1629780. [Google Scholar] [CrossRef]
- Jiang, P.; Gu, S.; Pan, D.; Fu, J.; Sahu, A.; Hu, X. Signatures of T cell dysfunction and exclusion predict cancer immunotherapy response. Nat. Med. 2018, 24, 1550–1558. [Google Scholar] [CrossRef]
- Ayers, M.; Lunceford, J.; Nebozhyn, M.; Murphy, E.; Loboda, A.; Kaufman, D.R.; Andrew, A.; Jonathan, D.; Cheng, S.; Peter, K.; et al. IFN-γ–related mRNA profile predicts clinical response to PD-1 blockade. J. Clin. Investig. 2017, 127, 2930–2940. [Google Scholar] [CrossRef]
- Liu, Z.; Han, C.; Fu, Y.-X. Targeting innate sensing in the tumor microenvironment to improve immunotherapy. Cell. Mol. Immunol. 2020, 17, 13–26. [Google Scholar] [CrossRef] [PubMed]
- Ponzoni, M.; Pastorino, F.; Di Paolo, D.; Perri, P.; Brignole, C. Targeting Macrophages as a Potential Therapeutic Intervention: Impact on Inflammatory Diseases and Cancer. Int. J. Mol. Sci. 2018, 19, 1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atri, C.; Guerfali, F.; Laouini, D. Role of Human Macrophage Polarization in Inflammation during Infectious Diseases. Int. J. Mol. Sci. 2018, 19, 1801. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Huang, Q.; Xiao, W.; Zhao, Y.; Pi, J.; Xu, H. Advances in Anti-Tumor Treatments Targeting the CD47/SIRPα Axis. Front. Immunol. 2020, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Jalil, A.R.; Andrechak, J.C.; Discher, D.E. Macrophage checkpoint blockade: Results from initial clinical trials, binding analyses, and CD47-SIRPα structure–function. Antib. Ther. 2020, 3, 80–94. [Google Scholar] [CrossRef] [Green Version]
- Brown, E. Integrin-associated protein (CD47) and its ligands. Trends Cell Biol. 2001, 11, 130–135. [Google Scholar] [CrossRef]
- Adams, S.; van der Laan, L.J.; Vernon-Wilson, E.; Renardel de Lavalette, C.; Döpp, E.A.; Dijkstra, C.D. Signal-regulatory protein is selectively expressed by myeloid and neuronal cells. J. Immunol. 1998, 161, 1853–1859. [Google Scholar]
- Chao, M.P.; Weissman, I.L.; Majeti, R. The CD47–SIRPα pathway in cancer immune evasion and potential therapeutic implications. Curr. Opin. Immunol. 2012, 24, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Kuleshov, M.V.; Jones, M.R.; Rouillard, A.D.; Fernandez, N.F.; Duan, Q.; Wang, Z. Enrichr: A comprehensive gene set enrichment analysis web server 2016 update. Nucleic Acids Res. 2016, 44, W90–W97. [Google Scholar] [CrossRef] [Green Version]
- Chuang, Y.; Hung, M.E.; Cangelose, B.K.; Leonard, J.N. Regulation of the IL-10-driven macrophage phenotype under incoherent stimuli. Innate Immun. 2016, 22, 647–657. [Google Scholar] [CrossRef] [Green Version]
- Sierra-Filardi, E.; Puig-Kröger, A.; Blanco, F.J.; Nieto, C.; Bragado, R.; Palomero, M.I. Activin A skews macrophage polarization by promoting a proinflammatory phenotype and inhibiting the acquisition of anti-inflammatory macrophage markers. Blood 2011, 117, 5092–5101. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Tang, Z.; Gao, S.; Li, C.; Feng, Y.; Zhou, X. Tumor-Associated Macrophages: Recent Insights and Therapies. Front. Oncol. 2020, 10, 188. [Google Scholar] [CrossRef]
- Gonzalez-Dominguez, E.; Dominguez-Soto, A.; Nieto, C.; Flores-Sevilla, J.L.; Pacheco-Blanco, M.; Campos-Pena, V. Atypical Activin A and IL-10 Production Impairs Human CD16+ Monocyte Differentiation into Anti-Inflammatory Macrophages. J. Immunol. 2016, 196, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riera-Borrull, M.; Cuevas, V.D.; Alonso, B.; Vega, M.A.; Joven, J.; Izquierdo, E. Palmitate Conditions Macrophages for Enhanced Responses toward Inflammatory Stimuli via JNK Activation. J. Immunol. 2017, 199, 3858–3869. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Tanaka, I.; Kotani, M.; Miyaoka, F.; Sando, T.; Muro, S. Gene expression of the human prostaglandin E receptor EP4 subtype: Differential regulation in monocytoid and lymphoid lineage cells by phorbol ester. J. Mol. Med. 1996, 74, 333–336. [Google Scholar] [CrossRef]
- Bhat, N.K.; Thompson, C.B.; Lindsten, T.; June, C.H.; Fujiwara, S.; Koizumi, S. Reciprocal expression of human ETS1 and ETS2 genes during T-cell activation: Regulatory role for the protooncogene ETS1. Proc. Natl. Acad. Sci. USA 1990, 87, 3723–3727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felberg, J.; Lefebvre, D.C.; Lam, M.; Wang, Y.; Ng, D.H.W.; Birkenhead, D. Subdomain X of the Kinase Domain of Lck Binds CD45 and Facilitates Dephosphorylation. J. Biol. Chem. 2004, 279, 3455–3462. [Google Scholar] [CrossRef] [Green Version]
- Llano, M.; Lee, N.; Navarro, F.; García, P.; Albar, J.P.; Geraghty, D.E. HLA-E-bound peptides influence recognition by inhibitory and triggering CD94/NKG2 receptors: Preferential response to an HLA-G-derived nonamer. Eur. J. Immunol. 1998, 28, 2854–2863. [Google Scholar] [CrossRef]
- Tzachanis, D.; Li, L.; Lafuente, E.M.; Berezovskaya, A.; Freeman, G.J.; Boussiotis, V.A. Twisted gastrulation (Tsg) is regulated by Tob and enhances TGF-β signaling in activated T lymphocytes. Blood 2007, 109, 2944–2952. [Google Scholar] [CrossRef] [Green Version]
- Sahlender, D.A.; Roberts, R.C.; Arden, S.D.; Spudich, G.; Taylor, M.J.; Luzio, J.P. Optineurin links myosin VI to the Golgi complex and is involved in Golgi organization and exocytosis. J. Cell Biol. 2005, 169, 285–295. [Google Scholar] [CrossRef]
- Rooney, M.S.; Shukla, S.A.; Wu, C.J.; Getz, G.; Hacohen, N. Molecular and Genetic Properties of Tumors Associated with Local Immune Cytolytic Activity. Cell 2015, 160, 48–61. [Google Scholar] [CrossRef] [Green Version]
- Györffy, B.; Lanczky, A.; Eklund, A.C.; Denkert, C.; Budczies, J.; Li, Q. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1809 patients. Breast Cancer Res. Treat. 2010, 123, 725–731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldhirsch, A.; Winer, E.P.; Coates, A.S.; Gelber, R.D.; Piccart-Gebhart, M.; Thürlimann, B. Personalizing the treatment of women with early breast cancer: Highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann. Oncol. 2013, 24, 2206–2223. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.Y.; Tan, C.M.; Kou, Y.; Duan, Q.; Wang, Z.; Meirelles, G. Enrichr: Interactive and collaborative HTML5 gene list enrichment analysis tool. BMC Bioinform. 2013, 14, 128. [Google Scholar] [CrossRef] [Green Version]
- Bateman, A. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar]
- Li, T.; Fan, J.; Wang, B.; Traugh, N.; Chen, Q.; Liu, J.S. TIMER: A Web Server for Comprehensive Analysis of Tumor-Infiltrating Immune Cells. Cancer Res. 2017, 77, e108–e110. [Google Scholar] [CrossRef] [Green Version]
- Cortazar, A.R.; Torrano, V.; Martín-Martín, N.; Caro-Maldonado, A.; Camacho, L.; Hermanova, I. CANCERTOOL: A Visualization and Representation Interface to Exploit Cancer Datasets. Cancer Res. 2018, 78, 6320–6328. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noblejas-López, M.d.M.; Baliu-Piqué, M.; Nieto-Jiménez, C.; Cimas, F.J.; Morafraile, E.C.; Pandiella, A.; Corbi, Á.L.; Győrffy, B.; Ocaña, A. Transcriptomic Profiles of CD47 in Breast Tumors Predict Outcome and Are Associated with Immune Activation. Int. J. Mol. Sci. 2021, 22, 3836. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083836
Noblejas-López MdM, Baliu-Piqué M, Nieto-Jiménez C, Cimas FJ, Morafraile EC, Pandiella A, Corbi ÁL, Győrffy B, Ocaña A. Transcriptomic Profiles of CD47 in Breast Tumors Predict Outcome and Are Associated with Immune Activation. International Journal of Molecular Sciences. 2021; 22(8):3836. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083836
Chicago/Turabian StyleNoblejas-López, María del Mar, Mariona Baliu-Piqué, Cristina Nieto-Jiménez, Francisco J. Cimas, Esther C. Morafraile, Atanasio Pandiella, Ángel L. Corbi, Balázs Győrffy, and Alberto Ocaña. 2021. "Transcriptomic Profiles of CD47 in Breast Tumors Predict Outcome and Are Associated with Immune Activation" International Journal of Molecular Sciences 22, no. 8: 3836. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083836