
Lithium Enhances the GABAergic Synaptic Activities on the Hypothalamic Preoptic Area (hPOA) Neurons


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
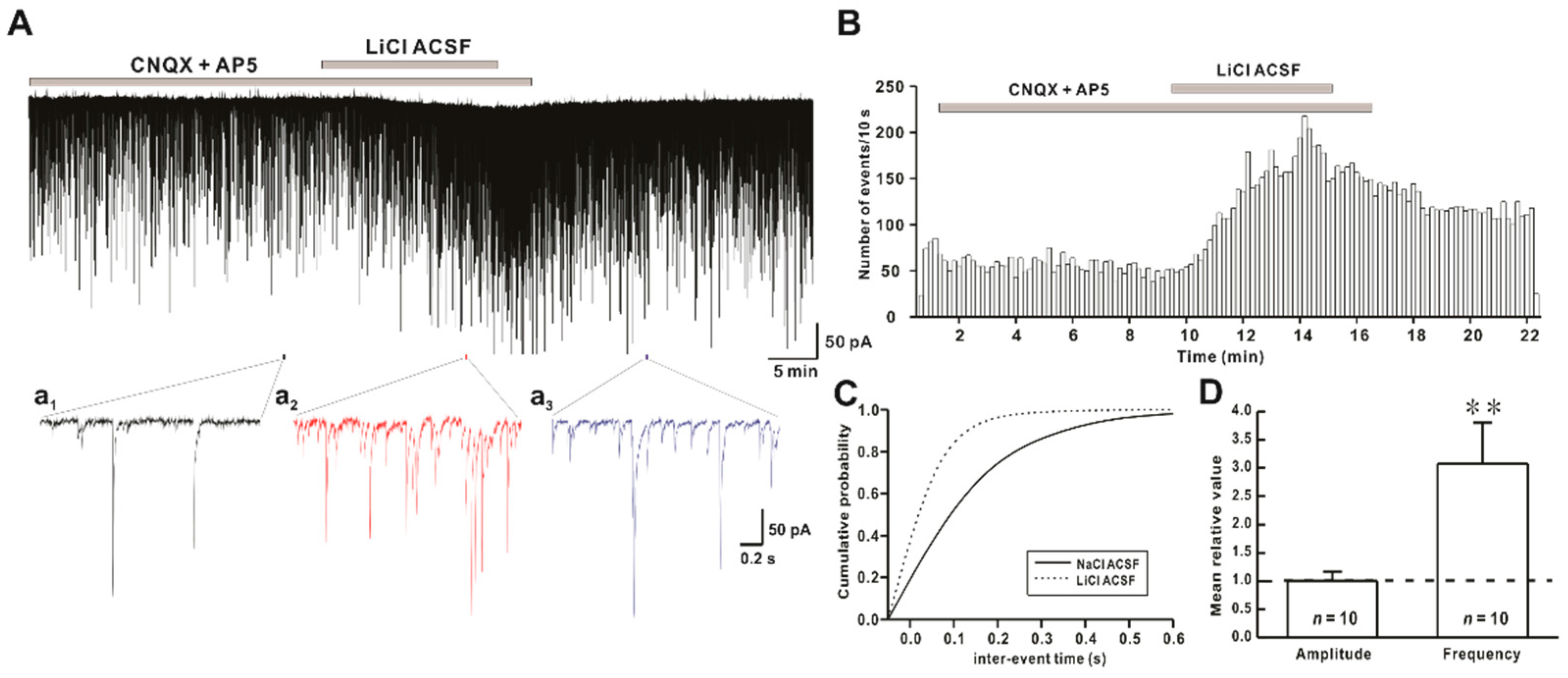
2.1. Li+ Enhances the Frequency of Spontaneous Inhibitory Postsynaptic Currents (sIPSCs)
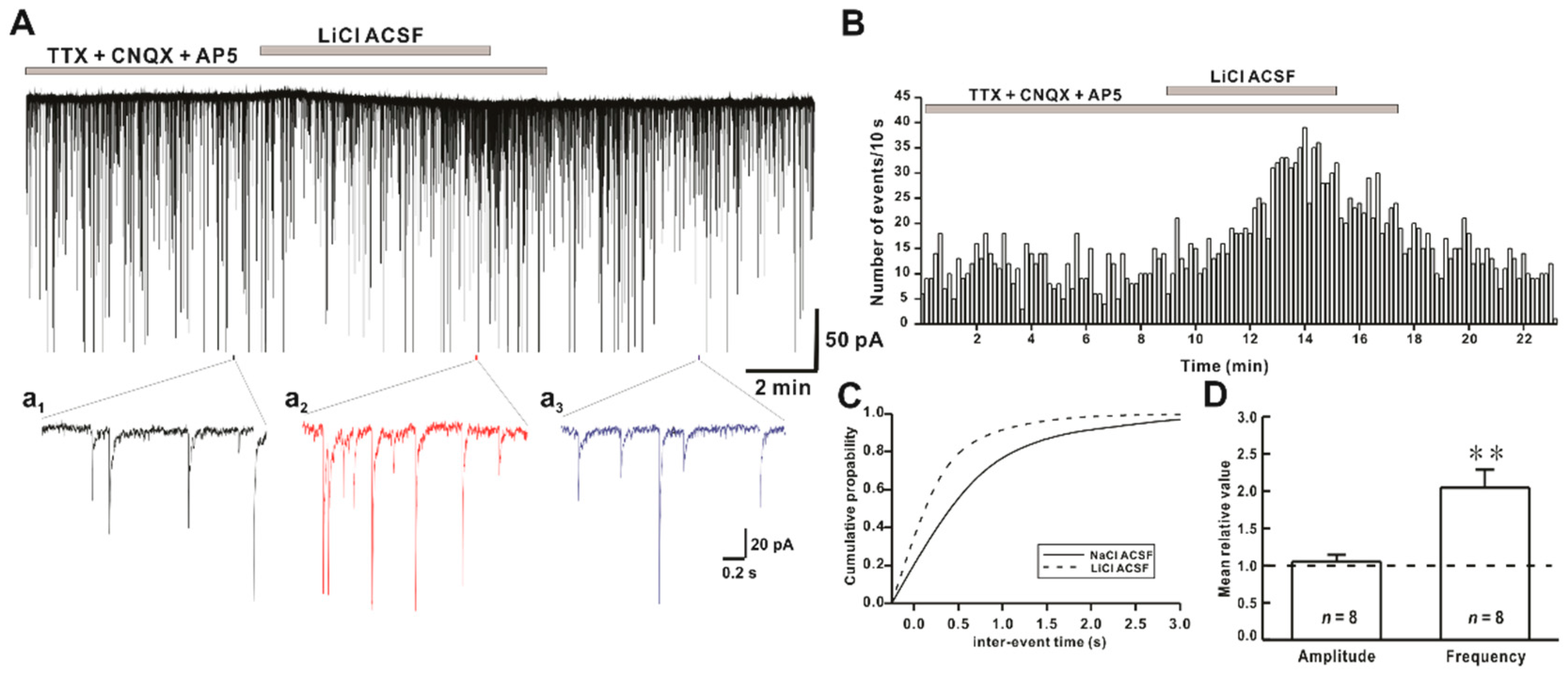
2.2. Li+ Enhances the Frequency of Miniature Inhibitory Postsynaptic Currents (mIPSCs)
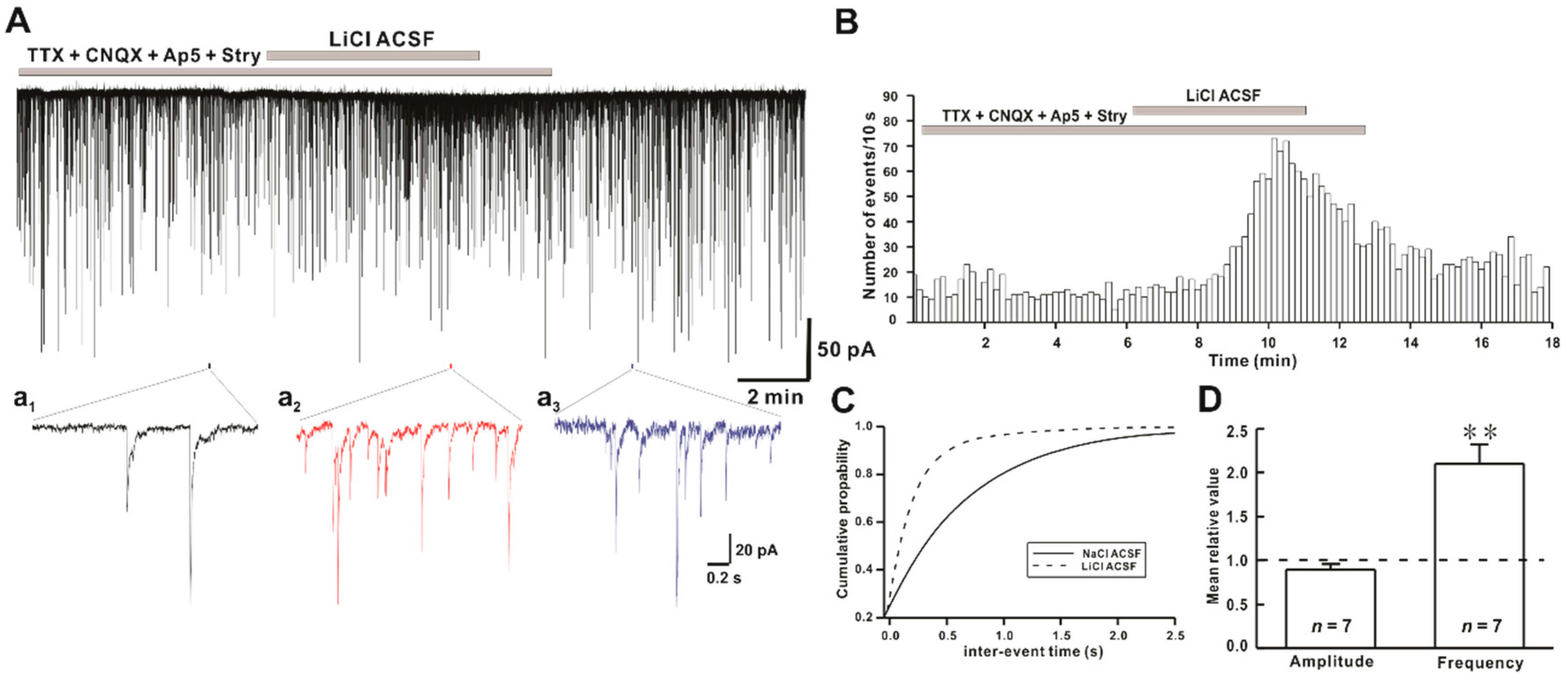
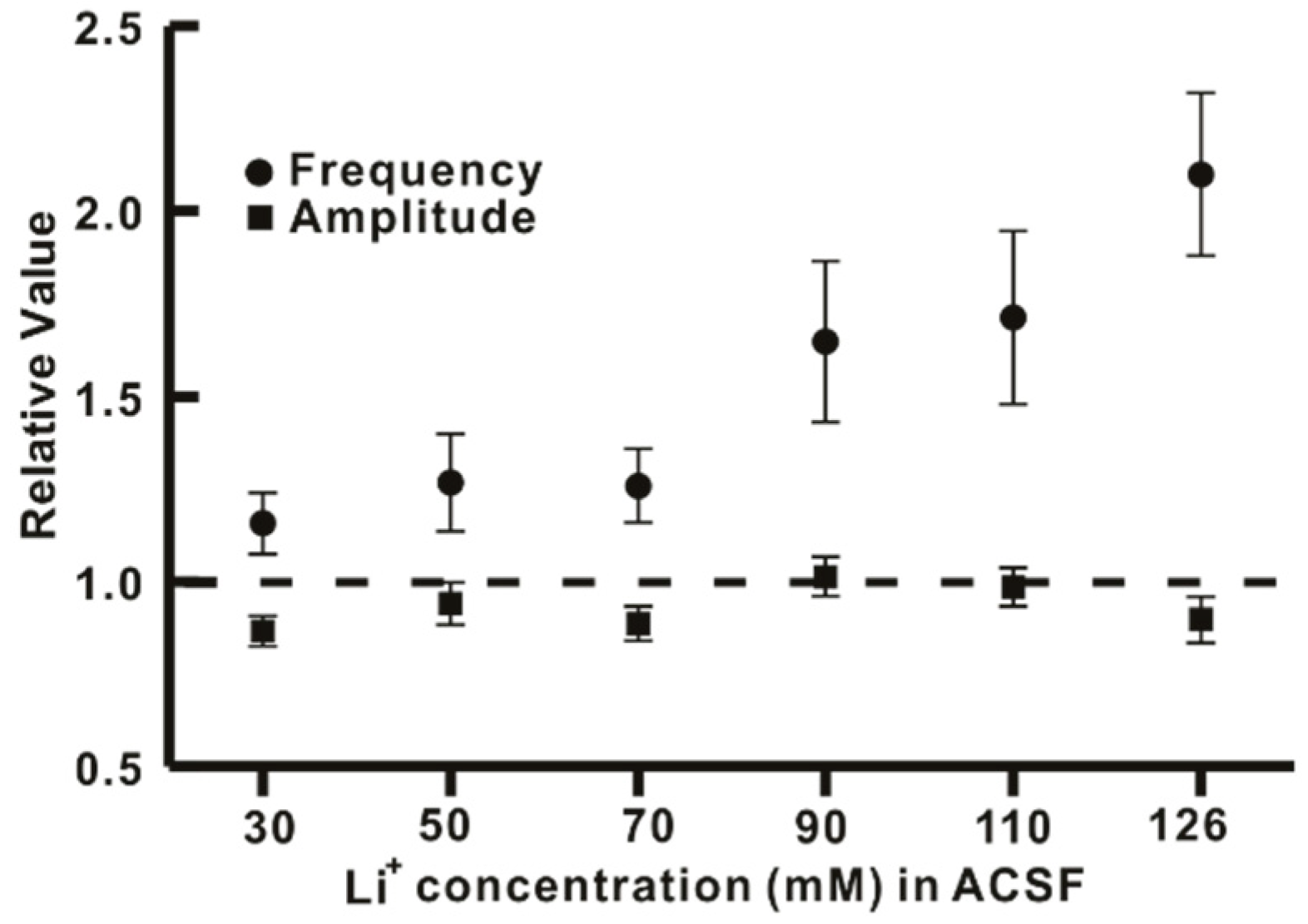
2.3. Li+ Increases the Frequency of GABAergic mIPSCs on hPOA Neurons
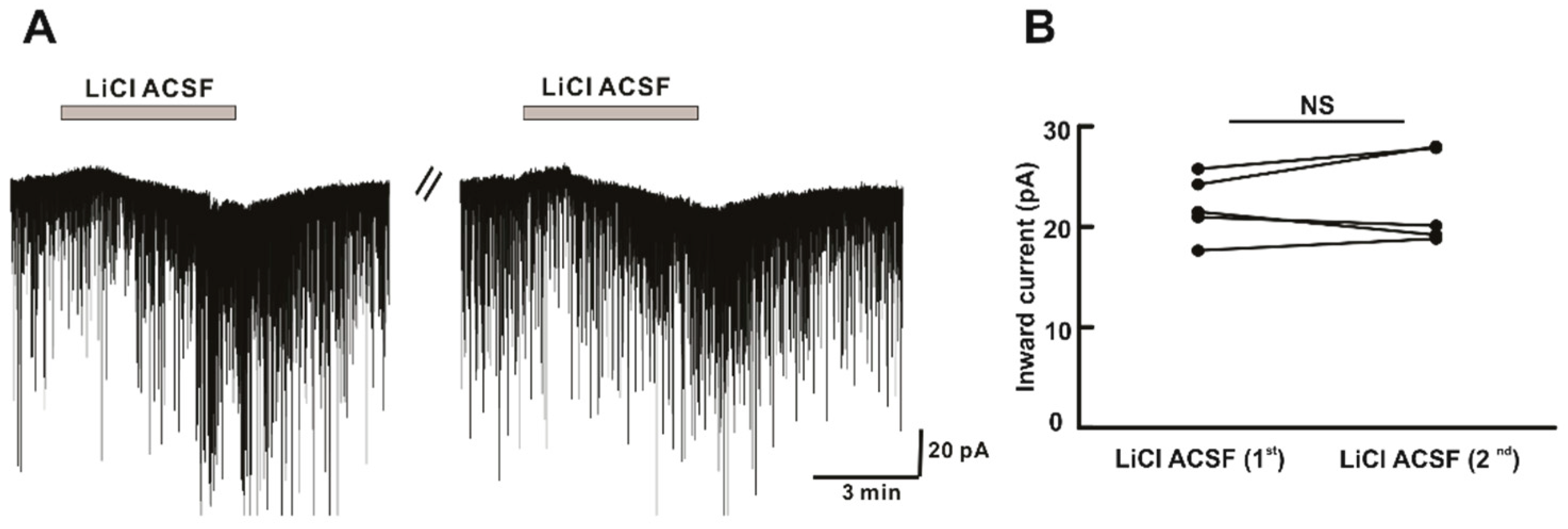
2.4. Li+ Perfusion Induces Repeatable Inward Currents on hPOA Neurons
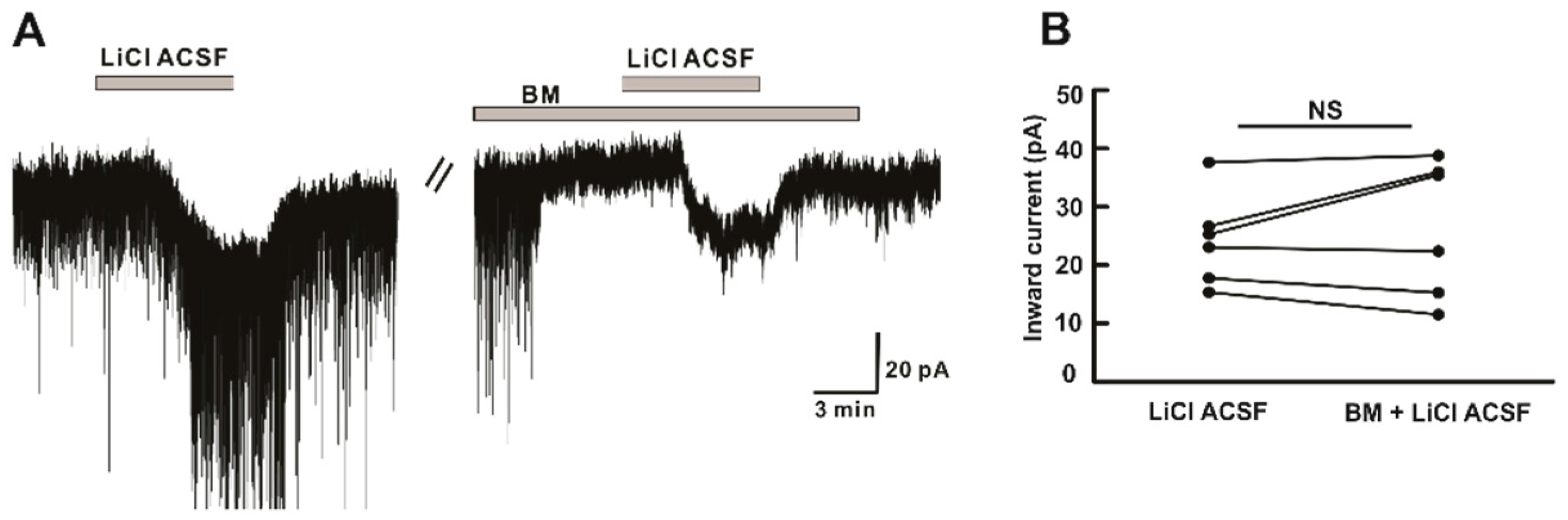
2.5. Li+ Directly Acts on hPOA Neurons and Its Action Is Independent of Amino-Acid Receptors
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Brain Slice Preparation
4.3. Whole-Cell Patch-Clamp Recording and Data Analysis
4.4. Chemicals
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Watanabe, M.; Maemura, K.; Kanbara, K.; Tamayama, T.; Hayasaki, H. GABA and GABA receptors in the central nervous system and other organs. Int. Rev. Cytol. 2002, 231, 1–47. [Google Scholar]
- Kirmse, K.; Holthoff, K. Functions of GABAergic transmission in the immature brain. e-Neuroforum 2017, 23, 27–33. [Google Scholar] [CrossRef]
- Ramamoorthi, K.; Lin, Y. The contribution of GABAergic dysfunction to neurodevelopmental disorders. Trends Mol. Med. 2011, 17, 452–462. [Google Scholar] [CrossRef]
- Deidda, G.; Bozarth, I.F.; Cancedda, L. Modulation of GABAergic transmission in development and neurodevelopmental disorders: Investigating physiology and pathology to gain therapeutic perspectives. Front. Cell Neurosci. 2014, 8, 119. [Google Scholar] [CrossRef] [Green Version]
- Belousov, A.B.; O’Hara, B.F.; Denisova, J.V. Acetylcholine becomes the major excitatory neurotransmitter in the hypothalamus in vitro in the absence of glutamate excitation. J. Neurosci. 2001, 21, 2015–2027. [Google Scholar] [CrossRef] [Green Version]
- Vincent, S.R.; Hökfelt, T.; Wu, J.Y. GABA neuron systems in hypothalamus and the pituitary gland. Neuroendocrinology 1982, 34, 117–125. [Google Scholar] [CrossRef]
- Rampon, C.; Luppi, P.H.; Fort, P.; Peyron, C.; Jouvet, M. Distribution of glycine-immunoreactive cell bodies and fibers in the rat brain. Neuroscience 1996, 75, 737–755. [Google Scholar] [CrossRef]
- Zeilhofer, H.U.; Studler, B.; Arabadzisz, D.; Schweizer, C.; Ahmadi, S.; Layh, B.; Bösl, M.R.; Fritschy, J.M. Glycinergic neurons expressing enhanced green fluorescent protein in bacterial artificial chromosome transgenic mice. J. Comp. Neurol. 2005, 482, 123–141. [Google Scholar] [CrossRef]
- Hoffman, N.W.; Wuarin, J.P.; Dudek, F.E. Whole-cell recordings of spontaneous synaptic currents in medial preoptic neurons from rat hypothalamic slices: Mediation by amino acid neurotransmitters. Brain Res. 1994, 660, 349–352. [Google Scholar] [CrossRef]
- McCann, S.M.; Vijayan, E.; Negro-Vilar, A.; Mizunuma, H.; Mangat, H. Gamma aminobutyric acid (GABA), a modulator of anterior pituitary hormone secretion by hypothalamic and pituitary action. Psychoneuroendocrinology 1984, 9, 97–106. [Google Scholar] [CrossRef]
- Yu, S.; François, M.; Huesing, C.; Münzberg, H. The hypothalamic preoptic area and body weight control. Neuroendocrinology 2017, 106, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikosha, A.S.; Kovzun, O.I.; Tronko, M.D. Biological effects of lithium–fundamental and medical aspects. Ukr. Biochem. J. 2017, 89, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Won, E.; Kim, Y.K. An oldie but goodie: Lithium in the treatment of bipolar disorder through neuroprotective and neurotrophic mechanisms. Int. J. Mol. Sci. 2017, 18, 2679. [Google Scholar] [CrossRef] [Green Version]
- Malhi, G.S.; Tanious, M.; Das, P.; Coulston, C.M.; Berk, M. Potential mechanisms of action of lithium in bipolar disorder. CNS Drugs 2013, 27, 135–153. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Sohn, J.W.; Ahn, S.C.; Park, W.S.; Ho, W.K. Li+ enhances GABAergic inputs to granule cells in the rat hippocampal dentate gyrus. Neuropharmacology 2004, 46, 638–646. [Google Scholar] [CrossRef]
- Smythe, G.A.; Brandstater, J.F.; Lazarus, L. Acute effects of lithium on central dopamine and serotonin activity reflected by inhibition of prolactin and growth hormone secretion in the rat. Aust. J. Biol. Sci. 1979, 32, 329–334. [Google Scholar] [CrossRef] [Green Version]
- Bschor, T.; Ritter, D.; Winkelmann, P.; Erbe, S.; Uhr, M.; Ising, M.; Lewitzka, U. Lithium monotherapy increases ACTH and cortisol response in the DEX/CRH test in unipolar depressed subjects. A study with 30 treatment-naive patients. PLoS ONE 2011, 6, e27613. [Google Scholar] [CrossRef] [Green Version]
- Anai, H.; Ueta, Y.; Serino, R.; Nomura, M.; Kabashima, N.; Shibuya, I.; Takasugi, M.; Nakashima, Y.; Yamashita, H. Upregulation of the expression of vasopressin gene in the paraventricular and supraoptic nuclei of the lithium-induced diabetes insipidus rat. Brain Res. 1997, 772, 161–166. [Google Scholar] [CrossRef]
- Burns, G.; Herz, A.; Nikolarakis, K. Stimulation of hypothalamic opioid peptide release by lithium is mediated by opioid autoreceptors: Evidence from a combined in vitro, ex vivo study. Neuroscience 1990, 36, 691–697. [Google Scholar] [CrossRef]
- Sheikha, S.H.; LeGate, L.S.; Banerji, T.K. Lithium suppresses ovariectomy-induced surges in plasma gonadotropins in rats. Life Sci. 1989, 44, 1363–1369. [Google Scholar] [CrossRef]
- Allagui, M.; Hfaiedh, N.; Croute, F.; Guermazi, F.; Vincent, C.; Soleilhavoup, J.; El, A.F. Side effects of low serum lithium concentrations on renal, thyroid, and sexual functions in male and female rats. Comptes Rendus Biol. 2005, 328, 900–911. [Google Scholar] [CrossRef]
- Bschor, T.; Adli, M.; Baethge, C.; Eichmann, U.; Ising, M.; Uhr, M.; Modell, S.; Künzel, H.; Müller-Oerlinghausen, B.; Bauer, M. Lithium augmentation increases the ACTH and cortisol response in the combined DEX/CRH test in unipolar major depression. Neuropsychopharmacology 2002, 27, 470–478. [Google Scholar] [CrossRef] [Green Version]
- Sugawara, M.; Hashimoto, K.; Hattori, T.; Takao, T.; Suemaru, S.; Ota, Z. Effects of lithium on the hypothalamo-pituitary-adrenal axis. Endocrinol. Jpn. 1988, 35, 655–663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lombardi, G.; Panza, N.; Biondi, B.; Di Lorenzo, L.; Lupoli, G.; Muscettola, G.; Carella, C.; Bellastella, A. Effects of lithium treatment on hypothalamic-pituitary-thyroid axis: A longitudinal study. J. Endocrinol. Investig. 1993, 16, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Kusalic, M.; Engelsmann, F. Effect of lithium maintenance treatment on hypothalamic pituitary gonadal axis in bipolar men. J. Psychiatry Neurosci. 1996, 21, 181–186. [Google Scholar] [PubMed]
- Tappaz, M.L.; Brownstein, M.J.; Kopin, I.J. Glutamate decarboxylase (GAD) and γ-aminobutyric acid (GABA) in discrete nuclei of hypothalamus and substantia nigra. Brain Res. 1977, 125, 109–121. [Google Scholar] [CrossRef]
- Bhattarai, J.P.; Park, S.J.; Chun, S.W.; Cho, D.H.; Han, S.K. Activation of synaptic and extrasynaptic glycine receptors by taurine in preoptic hypothalamic neurons. Neurosci. Lett. 2015, 608, 51–56. [Google Scholar] [CrossRef]
- Rijal, S.; Cho, D.H.; Park, S.-A.; Jang, S.H.; Ábrahám, I.M.; Han, S.K. Melatonin Suppresses the Kainate Receptor-Mediated Excitation on Gonadotropin-Releasing Hormone Neurons in Female and Male Prepubertal Mice. Int. J. Mol. Sci. 2020, 21, 5991. [Google Scholar] [CrossRef]
- Butler-Munro, C.; Coddington, E.J.; Shirley, C.H.; Heyward, P.M. Lithium modulates cortical excitability in vitro. Brain Res. 2010, 1352, 50–60. [Google Scholar] [CrossRef]
- Wu, M.; Zaborszky, L.; Hajszan, T.; Van Den Pol, A.N.; Alreja, M. Hypocretin/orexin innervation and excitation of identified septohippocampal cholinergic neurons. J. Neurosci. 2004, 24, 3527–3536. [Google Scholar] [CrossRef] [Green Version]
- Moffitt, J.R.; Bambah-Mukku, D.; Eichhorn, S.W.; Vaughn, E.; Shekhar, K.; Perez, J.D.; Rubinstein, N.D.; Hao, J.; Regev, A.; Dulac, C. Molecular, spatial, and functional single-cell profiling of the hypothalamic preoptic region. Science 2018, 362, 641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Decavel, C.; Van Den Pol, A.N. GABA: A dominant neurotransmitter in the hypothalamus. J. Comp. Neurol. 1990, 302, 1019–1037. [Google Scholar] [CrossRef] [PubMed]
- Van Den Pol, A.N.; Trombley, P.Q. Glutamate neurons in hypothalamus regulate excitatory transmission. J. Neurosci. 1993, 13, 2829–2836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacaille, J.C.; Cloutier, S.; Reader, T.A. Lithium reduced synaptic transmisson and increased neuronal excitability without altering endogenous serotonin, norepinephrine and dopamine in rat hippocampal slices in vitro. Prog. Neuropsychopharmacol. Biol. Psychiatry 1992, 3, 397–412. [Google Scholar] [CrossRef]
- Higashitani, Y.; Kudo, Y.; Ogura, A.; Kato, H. Acute effects of lithium on synaptic transmission in rat hippocampus studied in vitro. Biol. Psychiatry 1990, 27, 174–182. [Google Scholar] [CrossRef]
- Valentín, A.; García-Seoane, J.J.; Colino, A. Lithium enhances synaptic transmission in neonatal rat hippocampus. Neuroscience 1997, 78, 385–391. [Google Scholar] [CrossRef]
- Maggi, A.; Enna, S. Regional alterations in rat brain neurotransmitter systems following chronic lithium treatment. J. Neurochem. 1980, 34, 888–892. [Google Scholar] [CrossRef]
- Tanimoto, K.; Maeda, K.; Terada, T. Inhibitory effect of lithium on neuroleptic and serotonin receptors in rat brain. Brain Res. 1983, 265, 148–151. [Google Scholar] [CrossRef]
- Antonelli, T.; Ferioli, V.; Lo Gallo, G.; Tomasini, M.C.; Fernandez, M.; O’Connor, W.T.; Glennon, J.C.; Tanganelli, S.; Ferraro, L. Differential effects of acute and short-term lithium administration on dialysate glutamate and GABA levels in the frontal cortex of the conscious rat. Synapse 2000, 38, 355–362. [Google Scholar] [CrossRef]
- Gottesfeld, Z.; Ebstein, B.S.; Samuel, D. Effect of lithium on concentrations of glutamate and GABA levels in amygdala and hypothalamus of rat. Nat. New Biol. 1971, 234, 124–125. [Google Scholar] [CrossRef]
- Akoev, G.; Sizaya, N. Influence of lithium ions on the electrical activity of nerve cells of the leech. Neurophysiology 1970, 2, 484–489. [Google Scholar] [CrossRef]
- Grafe, P.; Reddy, M.; Emmert, H.; Ten Bruggencate, G. Effects of lithium on electrical activity and potassium ion distribution in the vertebrate central nervous system. Brain Res. 1983, 279, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Leung, L.S. Sodium-activated potassium conductance participates in the depolarizing afterpotential following a single action potential in rat hippocampal CA1 pyramidal cells. Brain Res. 2004, 1023, 185–192. [Google Scholar] [CrossRef]
- Janka, Z.; Jones, D. Lithium entry into neural cells via sodium channels: A morphometric approach. Neuroscience 1982, 7, 2849–2857. [Google Scholar] [CrossRef]
- Obara, S.; Grundfest, H. Effects of lithium on different membrane components of crayfish stretch receptor neurons. J. Gen. Physiol. 1968, 51, 635–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giacobini, E.; Hovmark, S.; Stepita-Klauco, M. Studies on the Mechanism of Action of Lithium Ions: II. Potassium Sensitive Influx of Lithium Ions into the Crayfish Stretch Receptor Neuron Determined by Microflamephotometry. Acta Physiol. Scand. 1970, 80, 528–532. [Google Scholar] [CrossRef]
- Leite, J.A.; Orellana, A.M.M.; Kinoshita, P.F.; de Mello, N.P.; Scavone, C.; Kawamoto, E.M. Neuroinflammation and Neurotransmission Mechanisms Involved in Neuropsychiatric Disorders. In Mechanisms of Neuroinflammation; IntechOpen: London, UK, 2017. [Google Scholar]
- Sheffler, Z.M.; Reddy, V.; Pillarisetty, L.S. Physiology, Neurotransmitters; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Forlenza, O.V.; De-Paula, V.J.R.; Diniz, B.S.O. Neuroprotective effects of lithium: Implications for the treatment of Alzheimer’s disease and related neurodegenerative disorders. ACS Chem. Neurosci. 2014, 5, 443–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maletzky, B.M.; Shore, J.H. Lithium treatment for psychiatric disorders. West. J. Med. 1978, 128, 488–498. [Google Scholar]
- Segal, J. Lithium-an update on the mechanisms of action. Part two: Neural effects and neuroanatomical substrate. Afr. J. Psychiatry 2004, 7, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Shibuya-Tayoshi, S.; Tayoshi, S.Y.; Sumitani, S.; Ueno, S.; Harada, M.; Ohmori, T. Lithium effects on brain glutamatergic and GABAergic systems of healthy volunteers as measured by proton magnetic resonance spectroscopy. Prog. Neuropsychopharmacol. Biol. Psychiatry 2008, 32, 249–256. [Google Scholar] [CrossRef]
- Nonaka, S.; Hough, C.J.; Chuang, D.M. Chronic lithium treatment robustly protects neurons in the central nervous system against excitotoxicity by inhibiting N-methyl-D-aspartate receptor-mediated calcium influx. Proc. Natl. Acad. Sci. USA 1998, 95, 2642–2647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Gray, N.A.; Falke, C.A.; Chen, W.; Yuan, P.; Szabo, S.T.; Einat, H.; Manji, H.K. Modulation of synaptic plasticity by antimanic agents: The role of AMPA glutamate receptor subunit 1 synaptic expression. J. Neurosci. 2004, 24, 6578–6589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giusti, C.F.; Amorim, S.R.; Guerra, R.A.; Portes, E.S. Endocrine disturbances related to the use of lithium. Arq. Bras. Endocrinol. Metabol. 2012, 56, 153–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Filippa, V.P.; Mohamed, F.H. Lithium Therapy Effects on the Reproductive System. In Psychiatry and Neuroscience Update; Springer: Cham, Switzerland, 2019; pp. 187–200. [Google Scholar]
- Pfeifer, W.D.; Davis, L.; Van der Velde, C.D. Lithium accumulation in some endocrine tissues. Acta Biol. Med. Ger. 1976, 35, 1519–1523. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rijal, S.; Jang, S.H.; Park, S.J.; Han, S.K. Lithium Enhances the GABAergic Synaptic Activities on the Hypothalamic Preoptic Area (hPOA) Neurons. Int. J. Mol. Sci. 2021, 22, 3908. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083908
Rijal S, Jang SH, Park SJ, Han SK. Lithium Enhances the GABAergic Synaptic Activities on the Hypothalamic Preoptic Area (hPOA) Neurons. International Journal of Molecular Sciences. 2021; 22(8):3908. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083908
Chicago/Turabian StyleRijal, Santosh, Seon Hui Jang, Soo Joung Park, and Seong Kyu Han. 2021. "Lithium Enhances the GABAergic Synaptic Activities on the Hypothalamic Preoptic Area (hPOA) Neurons" International Journal of Molecular Sciences 22, no. 8: 3908. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22083908