
The Antihypertensive Effects and Potential Molecular Mechanism of Microalgal Angiotensin I-Converting Enzyme Inhibitor-Like Peptides: A Mini Review

 ,
,
Abstract
:1. Introduction
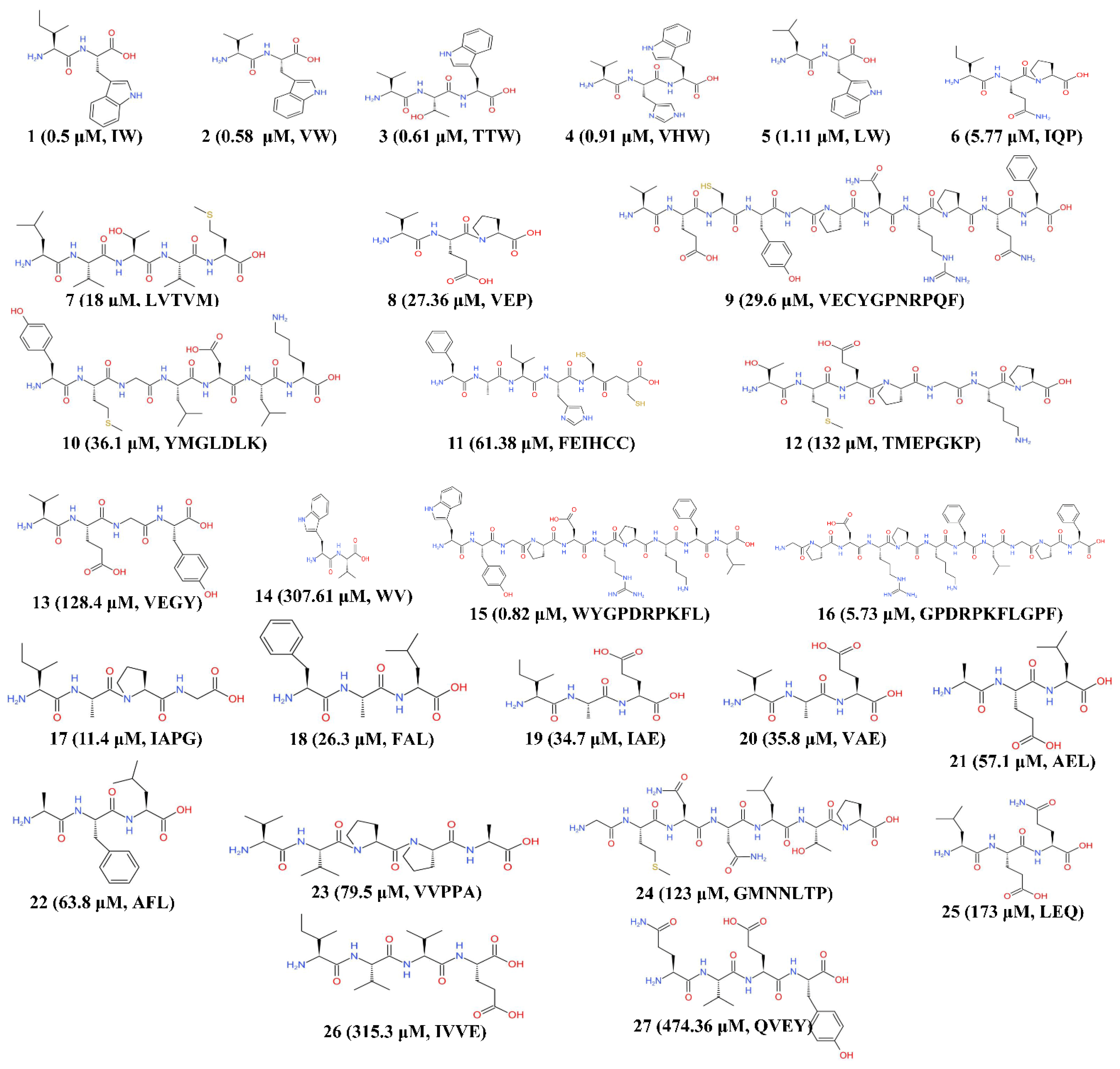
2. The Relationship between the Primary Structure of Microalgal Peptides and the ACE Inhibitory Effect
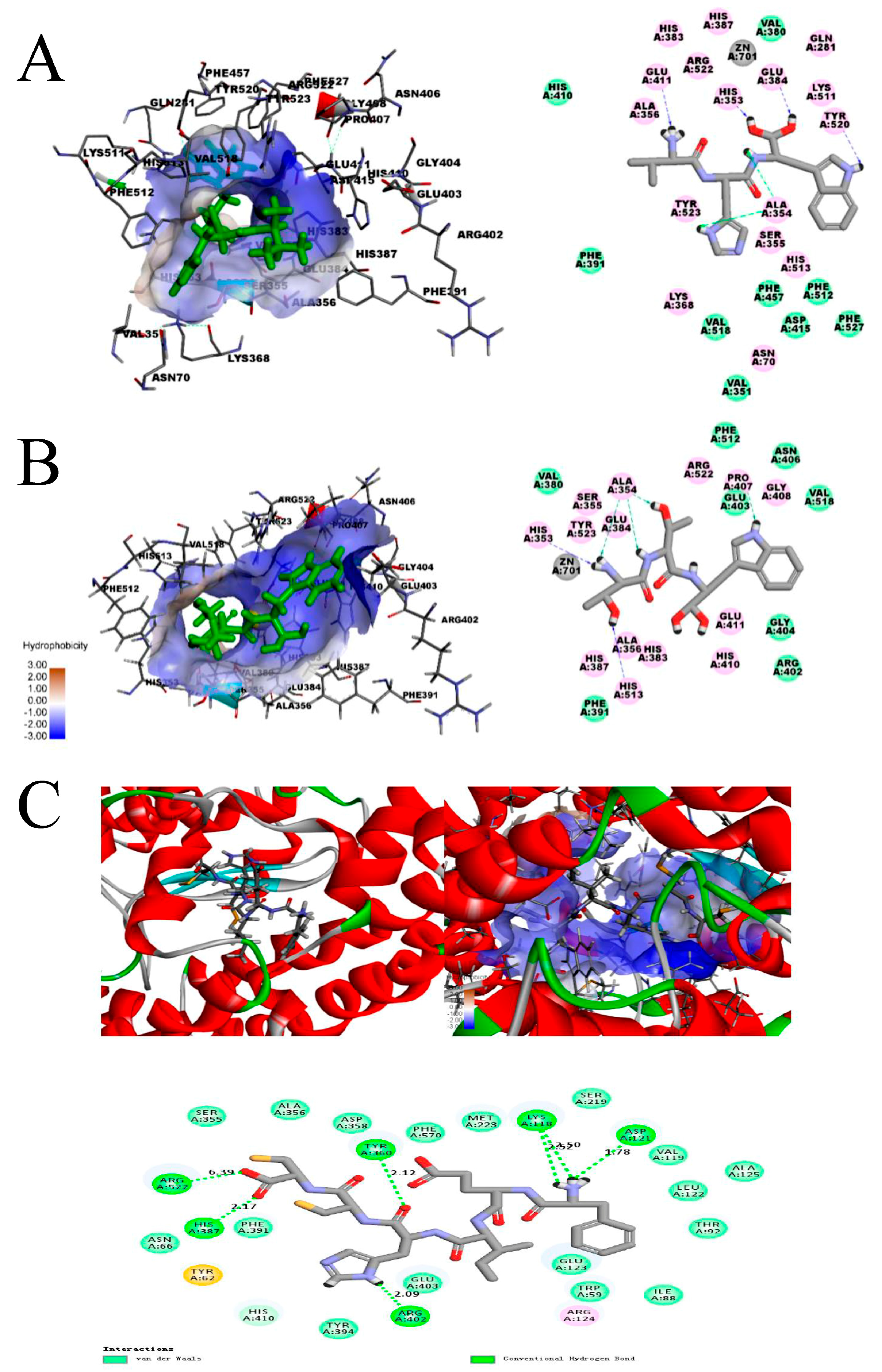
3. Molecular Docking of Microalgal Peptides to ACE
4. The Antioxidant and Anti-Inflammatory Properties of Microalgal ACE Inhibitory Peptides against Hypertension
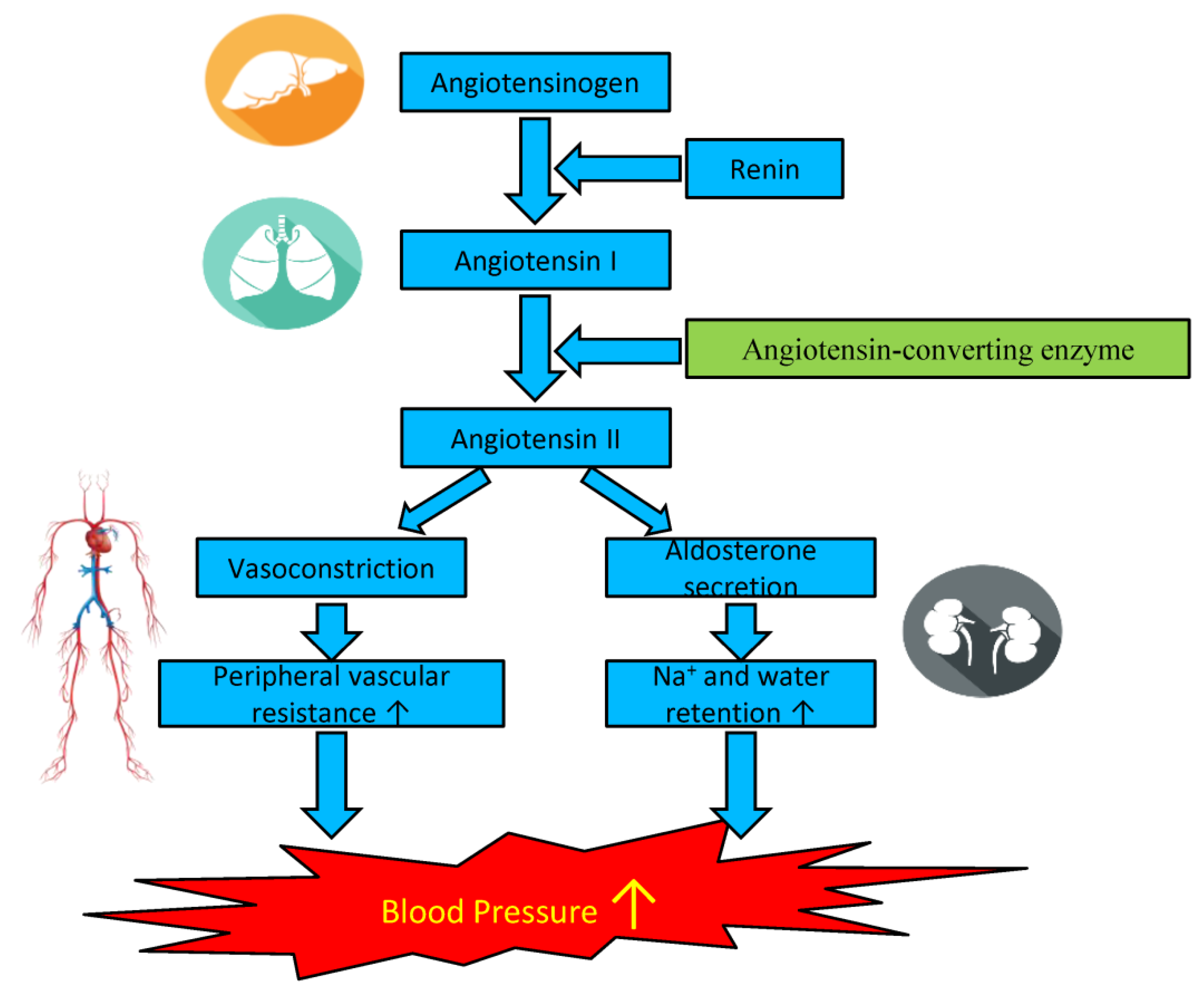
5. Antihypertensive Effects of Microalgal Biopeptides
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Morais, W.G.; Gorgich, M.; Correa, P.S.; Martins, A.A.; Mata, T.M.; Caetano, N.S. Microalgae for biotechnological applications: Cultivation, harvesting and biomass processing. Aquaculture 2020, 528. [Google Scholar] [CrossRef]
- Lafarga, T.; Fernandez-Sevilla, J.M.; Gonzalez-Lopez, C.; Acien-Fernandez, F.G. Spirulina for the food and functional food industries. Food Res. Int. 2020, 137. [Google Scholar] [CrossRef]
- Verma, S.; Kuila, A. Involvement of green technology in microalgal biodiesel production. Rev. Environ. Health 2020, 35, 173–188. [Google Scholar] [CrossRef] [PubMed]
- Markou, G.; Vandamme, D.; Muylaert, K. Microalgal and cyanobacterial cultivation: The supply of nutrients. Water Res. 2014, 65, 186–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matos, J.; Cardoso, C.; Bandarra, N.M.; Afonso, C. Microalgae as healthy ingredients for functional food: A review. Food Funct. 2017, 8, 2672–2685. [Google Scholar] [CrossRef]
- da Maia, J.L.; Cardoso, J.S.; Mastrantonio, D.J.D.; Bierhals, C.K.; Moreira, J.B.; Costa, J.A.V.; de Morais, M.G. Microalgae starch: A promising raw material for the bioethanol production. Int. J. Biol. Macromol. 2020, 165, 2739–2749. [Google Scholar] [CrossRef] [PubMed]
- Bekirogullari, M.; Figueroa-Torres, G.M.; Pittman, J.K.; Theodoropoulos, C. Models of microalgal cultivation for added-value products—A review. Biotechnol. Adv. 2020, 44. [Google Scholar] [CrossRef] [PubMed]
- Chew, K.W.; Yap, J.Y.; Show, P.L.; Suan, N.H.; Juan, J.C.; Ling, T.C.; Lee, D.J.; Chang, J.S. Microalgae biorefinery: High value products perspectives. Bioresour. Technol. 2017, 229, 53–62. [Google Scholar] [CrossRef]
- Amorim, M.L.; Soares, J.; Coimbra, J.S.D.; Leite, M.D.; Albino, L.F.T.; Martins, M.A. Microalgae proteins: Production, separation, isolation, quantification, and application in food and feed. Crit. Rev. Food Sci. 2020. [Google Scholar] [CrossRef]
- Canavate, J.P. Advancing assessment of marine phytoplankton community structure and nutritional value from fatty acid profiles of cultured microalgae. Rev. Aquacult. 2019, 11, 527–549. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, A.; Sabae, S.A.; Helal, A.M. Culture and Biorefinary of Two Freshwater Microalgae; Spirulina platensis and Chlorella vulgaris As Vitamins Sources. Biosci. Res. 2018, 15, 4584–4589. [Google Scholar]
- Panahi, Y.; Darvishi, B.; Jowzi, N.; Beiraghdar, F.; Sahebkar, A. Chlorella vulgaris: A Multifunctional Dietary Supplement with Diverse Medicinal Properties. Curr. Pharm. Des. 2016, 22, 164–173. [Google Scholar] [CrossRef]
- Ugya, A.Y.; Imam, T.S.; Li, A.F.; Ma, J.C.; Hua, X.Y. Antioxidant response mechanism of freshwater microalgae species to reactive oxygen species production: A mini review. Chem. Ecol. 2020, 36, 174–193. [Google Scholar] [CrossRef]
- Novoveska, L.; Ross, M.E.; Stanley, M.S.; Pradelles, R.; Wasiolek, V.; Sassi, J.F. Microalgal Carotenoids: A Review of Production, Current Markets, Regulations, and Future Direction. Mar. Drugs 2019, 17, 640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondal, A.; Bose, S.; Banerjee, S.; Patra, J.K.; Malik, J.; Mandal, S.K.; Kilpatrick, K.L.; Das, G.; Kerry, R.G.; Fimognari, C.; et al. Marine Cyanobacteria and Microalgae Metabolites—A Rich Source of Potential Anticancer Drugs. Mar. Drugs 2020, 18, 476. [Google Scholar] [CrossRef] [PubMed]
- Hussein, H.A.; Abdullah, M.A. Anticancer Compounds Derived from Marine Diatoms. Mar. Drugs 2020, 18, 356. [Google Scholar] [CrossRef]
- Tabarzad, M.; Atabaki, V.; Hosseinabadi, T. Anti-inflammatory Activity of Bioactive Compounds from Microalgae and Cyanobacteria by Focusing on the Mechanisms of Action. Mol. Biol. Rep. 2020, 47, 6193–6205. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, S.; Nailwal, N.; Kumar, M.; Barve, K. Recent Patents and Discovery of Anti-Inflammatory Agents from Marine Source. Recent Pat. Infla Allergy Drug Discov. 2019, 13, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Leng, L.J.; Wei, L.; Xiong, Q.; Xu, S.Y.; Li, W.T.; Lv, S.; Lu, Q.; Wan, L.P.; Wen, Z.Y.; Zhou, W.G. Use of microalgae based technology for the removal of antibiotics from wastewater: A review. Chemosphere 2020, 238. [Google Scholar] [CrossRef] [PubMed]
- Falaise, C.; Francois, C.; Travers, M.A.; Morga, B.; Haure, J.; Tremblay, R.; Turcotte, F.; Pasetto, P.; Gastineau, R.; Hardivillier, Y.; et al. Antimicrobial Compounds from Eukaryotic Microalgae against Human Pathogens and Diseases in Aquaculture. Mar. Drugs 2016, 14, 159. [Google Scholar] [CrossRef] [Green Version]
- Yarkent, C.; Gurlek, C.; Oncel, S.S. Potential of microalgal compounds in trending natural cosmetics: A review. Sustain. Chem. Pharm. 2020, 17. [Google Scholar] [CrossRef]
- Raposo, M.F.D.; de Morais, A.M.M.B.; de Morais, R.M.S.C. Carotenoids from Marine Microalgae: A Valuable Natural Source for the Prevention of Chronic Diseases. Mar. Drugs 2015, 13, 5128–5155. [Google Scholar] [CrossRef] [PubMed]
- Amato, A.; Terzo, S.; Mule, F. Natural Compounds as Beneficial Antioxidant Agents in Neurodegenerative Disorders: A Focus on Alzheimer’s Disease. Antioxidants 2019, 8, 608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Ianora, A. Marine Organisms with Anti-Diabetes Properties. Mar. Drugs 2016, 14, 220. [Google Scholar] [CrossRef] [PubMed]
- Haldar, R. Global Brief on Hypertension: Silent Killer, Global Public Health Crisis. Indian J. Phys. Med. Rehabil. 2013, 24, 2. [Google Scholar] [CrossRef]
- Mendis, S.; Davis, S.; Norrving, B. Organizational Update The World Health Organization Global Status Report on Noncommunicable Diseases 2014; One More Landmark Step in the Combat Against Stroke and Vascular Disease. Stroke 2015, 46, 121–122. [Google Scholar] [CrossRef]
- Valensi, P.; Lorgis, L.; Cottin, Y. Prevalence, incidence, predictive factors and prognosis or silent myocardial infarction: A review of the literature. Arch. Cardiovasc. Dis. 2011, 104, 178–188. [Google Scholar] [CrossRef] [Green Version]
- Spence, J.D.; Hammond, R. Hypertension and Stroke. In Hypertension and the Brain as an End-Organ Target; Girouard, H., Ed.; Springer International Publishing: Cham, Switzerland, 2016; pp. 39–54. [Google Scholar]
- Schiffrin, E.L.; Lipman, M.L.; Mann, J.F.E. Chronic kidney disease—Effects on the cardiovascular system. Circulation 2007, 116, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Pullalarevu, R.; Akbar, G.; Teehan, G. Secondary Hypertension, Issues in Diagnosis and Treatment. Prim. Care 2014, 41, 749–764. [Google Scholar] [CrossRef] [PubMed]
- Tuttolomondo, A.; Di Raimondo, D.; Pecoraro, R.; Arnao, V.; Pinto, A.; Licata, G. Atherosclerosis as an Inflammatory Disease. Curr. Pharm. Design 2012, 18, 4266–4288. [Google Scholar] [CrossRef]
- Nava, E.; Llorens, S. The Local Regulation of Vascular Function: From an Inside-Outside to an Outside-Inside Model. Front. Physiol. 2019, 10. [Google Scholar] [CrossRef]
- Aluko, R.E. Food protein-derived renin-inhibitory peptides: In vitro and in vivo properties. J. Food Biochem. 2019, 43. [Google Scholar] [CrossRef] [Green Version]
- Erdmann, K.; Cheung, B.W.Y.; Schroder, H. The possible roles of food-derived bioactive peptides in reducing the risk of cardiovascular disease. J. Nutr. Biochem. 2008, 19, 643–654. [Google Scholar] [CrossRef]
- Chirumamilla, R.R.; Marchant, R.; Nigam, P. Captopril and its synthesis from chiral intermediates. J. Chem. Technol. Biotechnol. 2001, 76, 123–127. [Google Scholar] [CrossRef]
- Todd, P.A.; Goa, K.L. Enalapril. Drugs 1992, 43, 346–381. [Google Scholar] [CrossRef] [PubMed]
- Burrello, J.; Erhardt, E.M.; Saint-Hilary, G.; Veglio, F.; Rabbia, F.; Mulatero, P.; Monticone, S.; D’Ascenzo, F. Pharmacological Treatment of Arterial Hypertension in Children and Adolescents: A Network Meta-Analysis. Hypertension 2018, 72, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Barrios, V.; Escobar, C. Antihypertensive and organ-protective effects of benazepril. Expert Rev. Cardiovasc. Ther. 2010, 8, 1653–1671. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Musoles, R.; Salom, J.B.; Martinez-Maqueda, D.; Lopez-Diez, J.J.; Recio, I.; Manzanares, P. Antihypertensive effects of lactoferrin hydrolyzates: Inhibition of angiotensin and endothelin-converting enzymes. Food Chem. 2013, 139, 994–1000. [Google Scholar] [CrossRef] [PubMed]
- Ruzicka, M.; Coletta, E.; White, R.; Davies, R.; Haddad, H.; Leenen, F.H.H. Effects of ACE Inhibitors on Cardiac Angiotensin II and Aldosterone in Humans: “Relevance of Lipophilicity and Affinity for ACE”. Am. J. Hypertens. 2010, 23, 1179–1182. [Google Scholar] [CrossRef] [Green Version]
- Gomes, C.; Ferreira, D.; Carvalho, J.P.F.; Barreto, C.A.V.; Fernandes, J.; Gouveia, M.; Ribeiro, F.; Duque, A.S.; Vieira, S.I. Current genetic engineering strategies for the production of antihypertensive ACEI peptides. Biotechnol. Bioeng. 2020, 117, 2610–2628. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Fitzgerald, R.J. Enhancing bioactive peptide release and identification using targeted enzymatic hydrolysis of milk proteins. Anal. Bioanal. Chem. 2018, 410, 3407–3423. [Google Scholar] [CrossRef]
- Li-Chan, E.C.Y. Bioactive peptides and protein hydrolysates: Research trends and challenges for application as nutraceuticals and functional food ingredients. Curr. Opin. Food Sci. 2015, 1, 28–37. [Google Scholar] [CrossRef] [Green Version]
- Servaes, K.; Maesen, M.; Prandi, B.; Sforza, S.; Elst, K. Polar Lipid Profile of Nannochloropsis oculata Determined Using a Variety of Lipid Extraction Procedures. J. Agric. Food Chem. 2015, 63, 3931–3941. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.C.; Lammi, C.; Boschin, G.; Arnoldi, A.; Aiello, G. Recent Advances in Microalgae Peptides: Cardiovascular Health Benefits and Analysis. J. Agric. Food Chem. 2019, 67, 11825–11838. [Google Scholar] [CrossRef] [PubMed]
- Murakami, T.; Okamoto, K.; Ogaki, M.; Iizuka, Y. Effect of Chlorella on blood pressure, cerebral stroke lesions, hypertensive vascular changes and life-span in spontaneously hypertensive rats. Nippon Eiyo Shokuryo Gakkaishi. 1987, 40, 351–359. [Google Scholar] [CrossRef]
- Miyakoshi, M. Study of Chlorella producted from the Chikugo area. Clin. Rep. 1980, 14, 3931–3941. [Google Scholar]
- Inoue, K.; Mukaiyama, Y.; Oka, H.; Sansawa, H. Clinical Effects of Chlorella in Hypertensive Men. Nippon Eiyo Shokuryo Gakkaishi. 1995, 48, 485–489. [Google Scholar] [CrossRef] [Green Version]
- Suetsuna, K.; Chen, J.R. Identification of antihypertensive peptides from peptic digest of two microalgae, Chlorella vulgaris and Spirulina platensis. Mar. Biotechnol. 2001, 3, 305–309. [Google Scholar] [CrossRef]
- Samarakoon, K.; Jeon, Y.J. Bio-functionalities of proteins derived from marine algae—A review. Food Res. Int. 2012, 48, 948–960. [Google Scholar] [CrossRef]
- Chen, J.L.; Tan, L.; Li, C.Y.; Zhou, C.X.; Hong, P.Z.; Sun, S.L.; Qian, Z.J. Mechanism Analysis of a Novel Angiotensin-I-Converting Enzyme Inhibitory Peptide from Isochrysis zhanjiangensis Microalgae for Suppressing Vascular Injury in Human Umbilical Vein Endothelial Cells. J. Agric. Food Chem. 2020, 68, 4411–4423. [Google Scholar] [CrossRef]
- Heo, S.Y.; Ko, S.C.; Kim, C.S.; Oh, G.W.; Ryu, B.; Qian, Z.J.; Kim, G.; Park, W.S.; Choi, I.W.; Phan, T.T.V.; et al. A heptameric peptide purified from Spirulina sp. gastrointestinal hydrolysate inhibits angiotensin I-converting enzyme- and angiotensin II-induced vascular dysfunction in human endothelial cells. Int. J. Mol. Med. 2017, 39, 1072–1082. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, K.W.; O-Nam, K.; Ko, J.Y.; Lee, J.H.; Kang, M.C.; Kim, D.; Lee, J.B.; Lee, J.S.; Jeon, Y.J. Purification and identification of novel angiotensin-I converting enzyme (ACE) inhibitory peptides from cultured marine microalgae (Nannochloropsis oculata) protein hydrolysate. J. Appl. Phycol. 2013, 25, 1595–1606. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Xie, J.L.; Chen, X.J.; Wu, J.J.; Zhang, Y.Y.; Zhou, Y.; Zhang, L.J.; Tang, Y.J.; Wei, D.Z. Antihypertensive Effects, Molecular Docking Study, and Isothermal Titration Calorimetry Assay of Angiotensin I-Converting Enzyme Inhibitory Peptides from Chlorella vulgaris. J. Agric. Food Chem. 2018, 66, 1359–1368. [Google Scholar] [CrossRef] [PubMed]
- Carrizalez-Lopez, C.; Gonzalez-Ortega, O.; Ochoa-Mendez, C.E.; Galvan-Moreno, F.U.; Rosales-Mendoza, S.; Monreal-Escalante, E.; Banuelos-Hernandez, B.; Paz-Maldonado, L.M.T.; Castro-Moreno, P.; Ibarra-Barajas, M.; et al. Expression of multiple antihypertensive peptides as a fusion protein in the chloroplast of Chlamydomonas reinhardtii. J. Appl. Phycol. 2018, 30, 1701–1709. [Google Scholar] [CrossRef]
- Beaulieu, L. Insights into the Regulation of Algal Proteins and Bioactive Peptides Using Proteomic and Transcriptomic Approaches. Molecules 2019, 24, 1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, I.A.; de Magalhães, C.S.; Dardenne, L.E. Receptor–ligand molecular docking. Biophys. Rev. 2014, 6, 75–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, M.L.; Cheng, S.Z.; Lu, W.H.; Du, M. Advancement and prospects of bioinformatics analysis for studying bioactive peptides from food-derived protein: Sequence, structure, and functions. Trac-Trend Anal. Chem. 2018, 105, 7–17. [Google Scholar] [CrossRef]
- He, R.; Aluko, R.E.; Ju, X.R. Evaluating Molecular Mechanism of Hypotensive Peptides Interactions with Renin and Angiotensin Converting Enzyme. PLoS ONE 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Alashi, A.M.; Young, J.F.; Therkildsen, M.; Aluko, R.E. Enzyme inhibition kinetics and molecular interactions of patatin peptides with angiotensin I-converting enzyme and renin. Int. J. Biol. Macromol. 2017, 101, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Ni, H.; Li, L.; Liu, G.; Hu, S.Q. Inhibition Mechanism and Model of an Angiotensin I-Converting Enzyme (ACE)-Inhibitory Hexapeptide from Yeast (Saccharomyces cerevisiae). PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [Green Version]
- Hou, W.C.; Chen, H.; Lin, Y.H. Antioxidant peptides with angiotensin converting enzyme inhibitory activities and applications for angiotensin converting enzyme purification. J. Agric. Food Chem. 2003, 51, 1706–1709. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.L.; Ryu, B.; Zhang, Y.Y.; Liang, P.; Li, C.Y.; Zhou, C.X.; Yang, P.; Hong, P.Z.; Qian, Z.J. Comparison of an angiotensin-I-converting enzyme inhibitory peptide from tilapia (Oreochromis niloticus) with captopril: Inhibition kinetics, in vivo effect, simulated gastrointestinal digestion and a molecular docking study. J. Sci. Food Agric. 2020, 100, 315–324. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.Y.; Wang, C.C.; Lu, Y.L.; Wu, W.C.; Hou, W.C. Antioxidant, anti-semicarbazide-sensitive amine oxidase, and anti-hypertensive activities of geraniin isolated from Phyllanthus urinaria. Food Chem. Toxicol. 2008, 46, 2485–2492. [Google Scholar] [CrossRef]
- Pujiastuti, D.Y.; Amin, M.N.G.; Alamsjah, M.A.; Hsu, J.L. Marine Organisms as Potential Sources of Bioactive Peptides that Inhibit the Activity of Angiotensin I-Converting Enzyme: A Review. Molecules 2019, 24, 2541. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wu, Y.J.; Yang, C.F.; Liu, B.; Huang, Y.F. Hypotensive, hypoglycaemic and hypolipidaemic effects of bioactive compounds from microalgae and marine micro-organisms. Int. J. Food Sci. Technol. 2015, 50, 1705–1717. [Google Scholar] [CrossRef]
- Lin, Y.H.; Chen, G.W.; Yeh, C.H.; Song, H.; Tsai, J.S. Purification and Identification of Angiotensin I-Converting Enzyme Inhibitory Peptides and the Antihypertensive Effect of Chlorella sorokiniana Protein Hydrolysates. Nutrients 2018, 10, 1397. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Sawano, Y.; Miyakawa, T.; Xue, Y.L.; Cai, M.Y.; Egashira, Y.; Ren, D.F.; Tanokura, M. One-Week Antihypertensive Effect of Ile-Gln-Pro in Spontaneously Hypertensive Rats. J. Agric. Food Chem. 2011, 59, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Qian, Z.J.; Heo, S.J.; Oh, C.H.; Kang, D.H.; Jeong, S.H.; Park, W.S.; Choi, I.W.; Jeon, Y.J.; Jung, W.K. Angiotensin I-Converting Enzyme (ACE) Inhibitory Peptide Isolated from Biodiesel Byproducts of Marine Microalgae, Nannochloropsis Oculata. J. Biobased Mater. Bioenerg. 2013, 7, 135–142. [Google Scholar] [CrossRef]
- Lu, J.; Ren, D.F.; Xue, Y.L.; Sawano, Y.; Miyakawa, T.; Tanokura, M. Isolation of an Antihypertensive Peptide from Alcalase Digest of Spirulina platensis. J. Agric. Food Chem. 2010, 58, 7166–7171. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Xu, N.J.; Sun, X.; Yu, H.; Zhou, C.X. Hydrolysis and purification of ACE inhibitory peptides from the marine microalga Isochrysis galbana. J. Appl. Phycol. 2015, 27, 351–361. [Google Scholar] [CrossRef]
- Ko, S.C.; Kang, N.; Kim, E.A.; Kang, M.C.; Lee, S.H.; Kang, S.M.; Lee, J.B.; Jeon, B.T.; Kim, S.K.; Park, S.J.; et al. A novel angiotensin I-converting enzyme (ACE) inhibitory peptide from a marine Chlorella ellipsoidea and its antihypertensive effect in spontaneously hypertensive rats. Process Biochem. 2012, 47, 2005–2011. [Google Scholar] [CrossRef]
- Montone, C.M.; Capriotti, A.L.; Cavaliere, C.; La Barbera, G.; Piovesana, S.; Chiozzi, R.Z.; Lagana, A. Peptidomic strategy for purification and identification of potential ACE-inhibitory and antioxidant peptides in Tetradesmus obliquus microalgae. Anal. Bioanal. Chem. 2018, 410, 3573–3586. [Google Scholar] [CrossRef]
- Cao, D.Q.; Lv, X.J.; Xu, X.T.; Yu, H.; Sun, X.; Xu, N.J. Purification and identification of a novel ACE inhibitory peptide from marine alga Gracilariopsis lemaneiformis protein hydrolysate. Eur. Food Res. Technol. 2017, 243, 1829–1837. [Google Scholar] [CrossRef]
- Daskaya-Dikmen, C.; Yucetepe, A.; Karbancioglu-Guler, F.; Daskaya, H.; Ozcelik, B. Angiotensin-I-Converting Enzyme (ACE)-Inhibitory Peptides from Plants. Nutrients 2017, 9, 316. [Google Scholar] [CrossRef] [PubMed]
- de Castro, R.J.S.; Sato, H.H. Biologically active peptides: Processes for their generation, purification and identification and applications as natural additives in the food and pharmaceutical industries. Food Res. Int. 2015, 74, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.X.; An, Q.; Le, L.Q.; Geng, F.; Jiang, L.Z.; Yan, J.; Xiang, D.B.; Peng, L.X.; Zou, L.; Zhao, G.; et al. Prospects of cereal protein-derived bioactive peptides: Sources, bioactivities diversity, and production. Crit. Rev. Food Sci. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q. Preparation of Functional Peanut Oligopeptide and Its Biological Activity. In Peanut Processing Characteristics and Quality Evaluation; Wang, Q., Ed.; Springer: Singapore, 2018; pp. 461–537. [Google Scholar]
- Steiner, T. Structural Evidence for the Aromatic–(i+1) Amine Hydrogen Bond in Peptides: L-Tyr-L-Tyr-L-Leu Monohydrate. Acta Crystallogr. Sect. D 1998, 54, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Huang, A.Z.; Zhou, L.; Zhang, D.W.; Yao, J.L.; Zhang, Y. Molecular design and validation of halogen bonding orthogonal to hydrogen bonding in breast cancer MDM2-peptide complex. J. Mol. Graph. Model. 2016, 70, 40–44. [Google Scholar] [CrossRef]
- Fan, H.B.; Liao, W.; Wu, J.P. Molecular interactions, bioavailability, and cellular mechanisms of angiotensin-converting enzyme inhibitory peptides. J. Food Biochem. 2019, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caballero, J. Considerations for Docking of Selective Angiotensin-Converting Enzyme Inhibitors. Molecules 2020, 25, 295. [Google Scholar] [CrossRef] [Green Version]
- Wu, H.X.; Liu, Y.L.; Guo, M.R.; Xie, J.L.; Jiang, X.M. A Virtual Screening Method for Inhibitory Peptides of Angiotensin I-Converting Enzyme. J. Food Sci. 2014, 79, C1635–C1642. [Google Scholar] [CrossRef]
- Wang, X.M.; Chen, H.X.; Fu, X.G.; Li, S.Q.; Wei, J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. LWT-Food Sci. Technol. 2017, 75, 93–99. [Google Scholar] [CrossRef]
- Rohit, A.C.; Sathisha, K.; Aparna, H.S. A variant peptide of buffalo colostrum beta-lactoglobulin inhibits angiotensin I-converting enzyme activity. Eur. J. Med. Chem. 2012, 53, 211–219. [Google Scholar] [CrossRef] [PubMed]
- Jao, C.L.; Huang, S.L.; Hsu, K.C. Angiotensin I-converting enzyme inhibitory peptides: Inhibition mode, bioavailability, and antihypertensive effects. BioMedicine 2012, 2, 130–136. [Google Scholar] [CrossRef]
- Tu, M.L.; Wang, C.; Chen, C.; Zhang, R.Y.; Liu, H.X.; Lu, W.H.; Jiang, L.Z.; Du, M. Identification of a novel ACE-inhibitory peptide from casein and evaluation of the inhibitory mechanisms. Food Chem. 2018, 256, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Sun, L.P.; Zhuang, Y.L. Preparation and identification of novel inhibitory angiotensin-I-converting enzyme peptides from tilapia skin gelatin hydrolysates: Inhibition kinetics and molecular docking. Food Funct. 2018, 9, 5251–5259. [Google Scholar] [CrossRef]
- Briones, A.M.; Touyz, R.M. Oxidative Stress and Hypertension: Current Concepts. Curr. Hypertens. Rep. 2010, 12, 135–142. [Google Scholar] [CrossRef]
- Sheih, I.C.; Wu, T.K.; Fang, T.J. Antioxidant properties of a new antioxidative peptide from algae protein waste hydrolysate in different oxidation systems. Bioresour. Technol. 2009, 100, 3419–3425. [Google Scholar] [CrossRef]
- Majumder, K.; Wu, J.P. Molecular Targets of Antihypertensive Peptides: Understanding the Mechanisms of Action Based on the Pathophysiology of Hypertension. Int. J. Mol. Sci. 2015, 16, 256–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkia, I.; Al-Haj, L.; Hamid, A.A.; Zakaria, M.; Saari, N.; Zadjali, F. Indigenous marine diatoms as novel sources of bioactive peptides with antihypertensive and antioxidant properties. Int. J. Food Sci. Technol. 2019, 54, 1514–1522. [Google Scholar] [CrossRef]
- Mahdieh, G.; Fazilati, M.; Izadi, M.; Pilehvarian, A.; Nazem, H. Investigation of ACE Inhibitory Effect and Antioxidant Activity of Peptide Extracted from Spirulina Platensis. Chem. Methodol. 2020, 4, 172–180. [Google Scholar] [CrossRef] [Green Version]
- Barrows, I.R.; Ramezani, A.; Raj, D.S. Inflammation, Immunity, and Oxidative Stress in Hypertension-Partners in Crime? Adv. Chronic Kidney Dis. 2019, 26, 122–130. [Google Scholar] [CrossRef]
- Guzik, T.J.; Hoch, N.E.; Brown, K.A.; McCann, L.A.; Rahman, A.; Dikalov, S.; Goronzy, J.; Weyand, C.; Harrison, D.G. Role of the T cell in the genesis of angiotensin II-induced hypertension and vascular dysfunction. J. Exp. Med. 2007, 204, 2449–2460. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2. [Google Scholar] [CrossRef] [Green Version]
- Xu, W.; Liu, P.; Mu, Y.P. Research progress on signaling pathways in cirrhotic portal hypertension. World J. Clin. Cases 2018, 6, 335–343. [Google Scholar] [CrossRef]
- Li, M.; Liu, X.; He, Y.P.; Zheng, Q.Y.; Wang, M.; Wu, Y.; Zhang, Y.P.; Wang, C.Y. Celastrol attenuates angiotensin II mediated human umbilical vein endothelial cells damage through activation of Nrf2/ERK1/2/Nox2 signal pathway. Eur. J. Pharmacol. 2017, 797, 124–133. [Google Scholar] [CrossRef]
- Zhou, X.L.; Liang, L.W.; Zhao, Y.; Zhang, H. Epigallocatechin-3-Gallate Ameliorates Angiotensin II-Induced Oxidative Stress and Apoptosis in Human Umbilical Vein Endothelial Cells through the Activation of Nrf2/Caspase-3 Signaling. J. Vasc. Res. 2017, 54, 299–308. [Google Scholar] [CrossRef] [PubMed]
- Ochoa-Méndez, C.E.; Lara-Hernández, I.; González, L.M.; Aguirre-Bañuelos, P.; Ibarra-Barajas, M.; Castro-Moreno, P.; González-Ortega, O.; Soria-Guerra, R.E. Bioactivity of an antihypertensive peptide expressed in Chlamydomonas reinhardtii. J. Biotechnol. 2016, 240, 76–84. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Inhibitory Pattern | Species | No. | Peptides Sequence | IC50 (μM) | Study Type | Reference |
|---|---|---|---|---|---|---|
| Noncompetitive | Chlorella sorokiniana | 1 | IW | 0.5 | In vitro (rabbit lung) | [68] |
| 2 | VW | 0.58 | In vitro (rabbit lung) | |||
| Chlorella vulgaris | 3 | TTW | 0.61 | In vivo (rats) | [55] | |
| 4 | VHW | 0.91 | In vivo (rats) | |||
| Chlorella sorokiniana | 5 | LW | 1.11 | In vitro (rabbit lung) | [68] | |
| Spirulina platensis | 6 | IQP | 5.77 | In vivo (rats) | [69] | |
| Nannochloropsis oculata | 7 | LVTVM | 18 | In vitro | [70] | |
| Spirulina platensis | 8 | VEP | 27.36 | In vivo (rats) | [71] | |
| Chlorella vulgaris | 9 | VECYGPNRPQF | 29.6 | In vitro | [54] | |
| Isochrysis galbana | 10 | YMGLDLK | 36.1 | In vitro | [72] | |
| Isochrysis zhanjiangensis | 11 | FEIHCC | 61.38 | In vitro | [51] | |
| Spirulina sp. | 12 | TMEPGKP | 132 | In vitro | [52] | |
| Competitive | Chlorella ellipsoidea | 13 | VEGY | 128.4 | In vivo (rats) | [73] |
| Tetradesmus obliquus | 14 | WV | 307.61 | In vitro (rabbit lung) | [68] | |
| Not available | Tetradesmus obliquus | 15 | WYGPDRPKFL | 0.82 | In vitro | [74] |
| 16 | GPDRPKFLGPF | 5.73 | In vitro | |||
| Spirulina platensis | 17 | IAPG | 11.4 | In vivo (rats) | [49] | |
| Chlorella vulgaris | 18 | FAL | 26.3 | In vivo (rats) | ||
| Spirulina platensis | 19 | IAE | 34.7 | In vivo (rats) | ||
| 20 | VAE | 35.8 | In vivo (rats) | |||
| Chlorella vulgaris | 21 | AEL | 57.1 | In vivo (rats) | ||
| 22 | AFL | 63.8 | In vivo (rats) | |||
| 23 | VVPPA | 79.5 | In vivo (rats) | |||
| Nannochloropsis oculata | 24 | GMNNLTP | 123 | In vitro | [53] | |
| 25 | LEQ | 173 | In vitro | |||
| Chlorella vulgaris | 26 | IVVE | 315.3 | In vivo (rats) | [49] | |
| Gracilariopsis lemaneiformis | 27 | QVEY | 474.36 | In vitro | [75] |
| Peptides | Hydrogen Bond | Distance (Å) | Active Sites Pockets |
|---|---|---|---|
| TTW | His353 | 2.17 | S2 |
| Ala354 | 2.08 | S1 | |
| His513 | 2.48 | S2 | |
| Ala354 | 2.02 | S1 | |
| Ala354 | 2.46 | S1 | |
| Pro407 | 1.89 | - | |
| VHW | Glu411 | 2.08 | - |
| Ala354 | 2.41 | S1 | |
| Ala354 | 1.96 | S1 | |
| His353 | 2.45 | S2 | |
| Tyr520 | 2.23 | S2 | |
| Glu384 | 1.93 | S1 | |
| FEHICC | Arg522 | 6.39 | - |
| His387 | 2.17 | - | |
| Arg402 | 2.09 | - | |
| Tyr360 | 2.12 | - | |
| Lys118 | 2.52 | - | |
| 2.50 | |||
| Asp121 | 1.78 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Q.; Chen, Q.; Zhang, T.; Liu, M.; Duan, S.; Sun, X. The Antihypertensive Effects and Potential Molecular Mechanism of Microalgal Angiotensin I-Converting Enzyme Inhibitor-Like Peptides: A Mini Review. Int. J. Mol. Sci. 2021, 22, 4068. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084068
Jiang Q, Chen Q, Zhang T, Liu M, Duan S, Sun X. The Antihypertensive Effects and Potential Molecular Mechanism of Microalgal Angiotensin I-Converting Enzyme Inhibitor-Like Peptides: A Mini Review. International Journal of Molecular Sciences. 2021; 22(8):4068. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084068
Chicago/Turabian StyleJiang, Qichen, Qi Chen, Tongqing Zhang, Meng Liu, Shunshan Duan, and Xian Sun. 2021. "The Antihypertensive Effects and Potential Molecular Mechanism of Microalgal Angiotensin I-Converting Enzyme Inhibitor-Like Peptides: A Mini Review" International Journal of Molecular Sciences 22, no. 8: 4068. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22084068