
A New Tool to Study Parkinsonism in the Context of Aging: MPTP Intoxication in a Natural Model of Multimorbidity

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
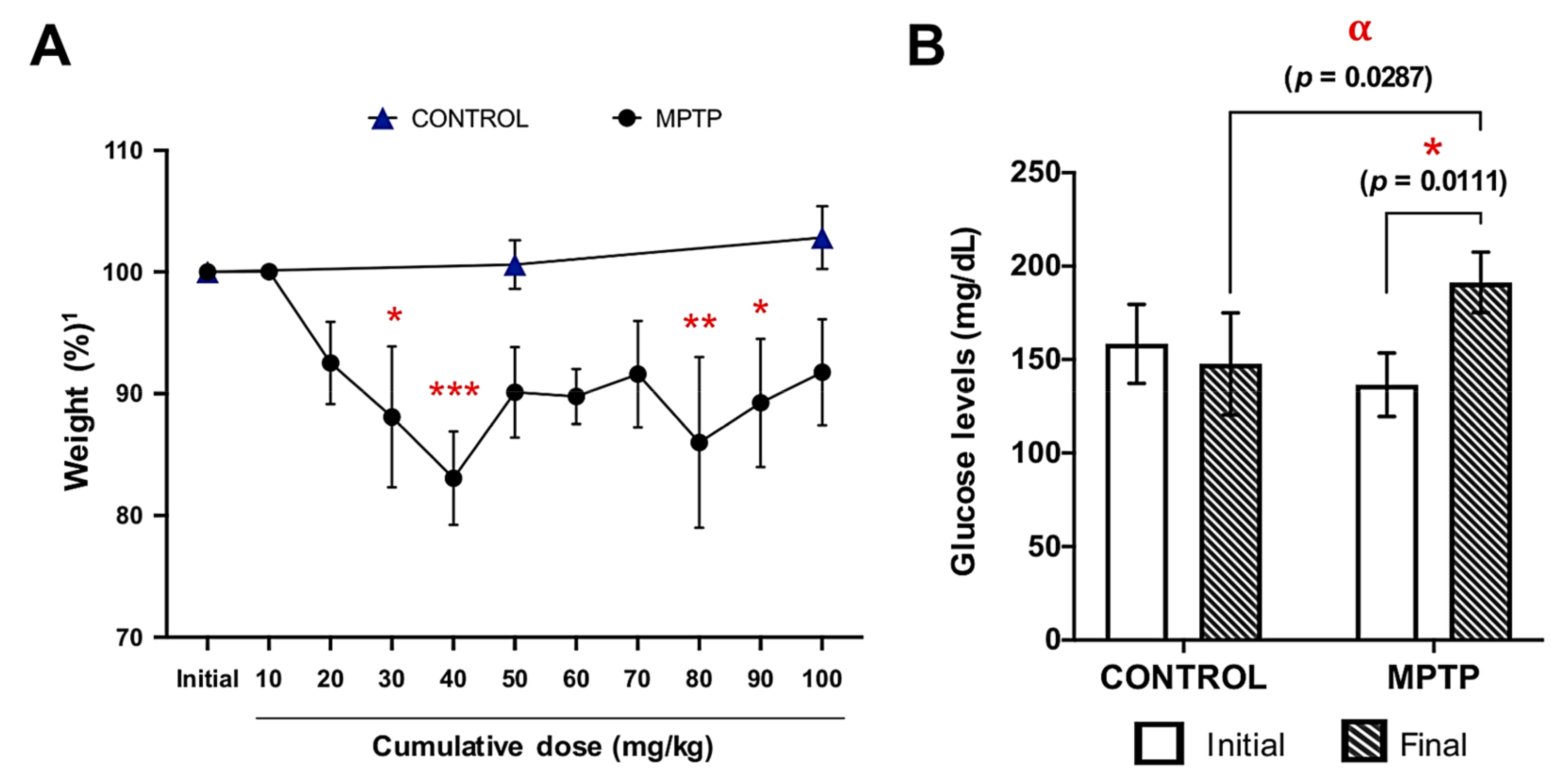
2.1. MPTP Intoxication Affects Body Weight
2.2. Increase in Glucose Levels Induced by MPTP Administration
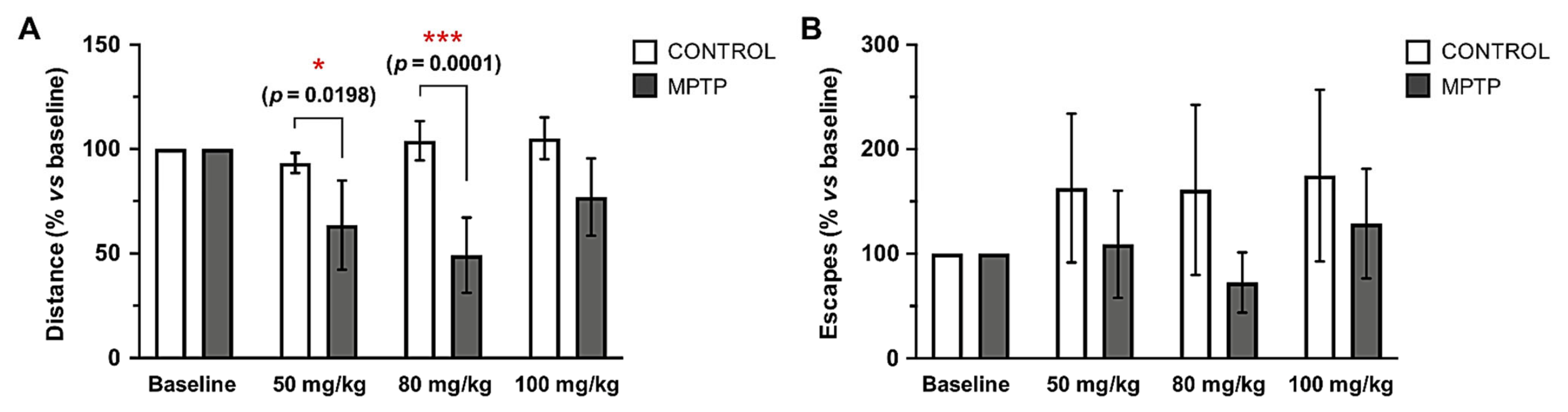
2.3. MPTP Affects Motor Condition in the O. degus
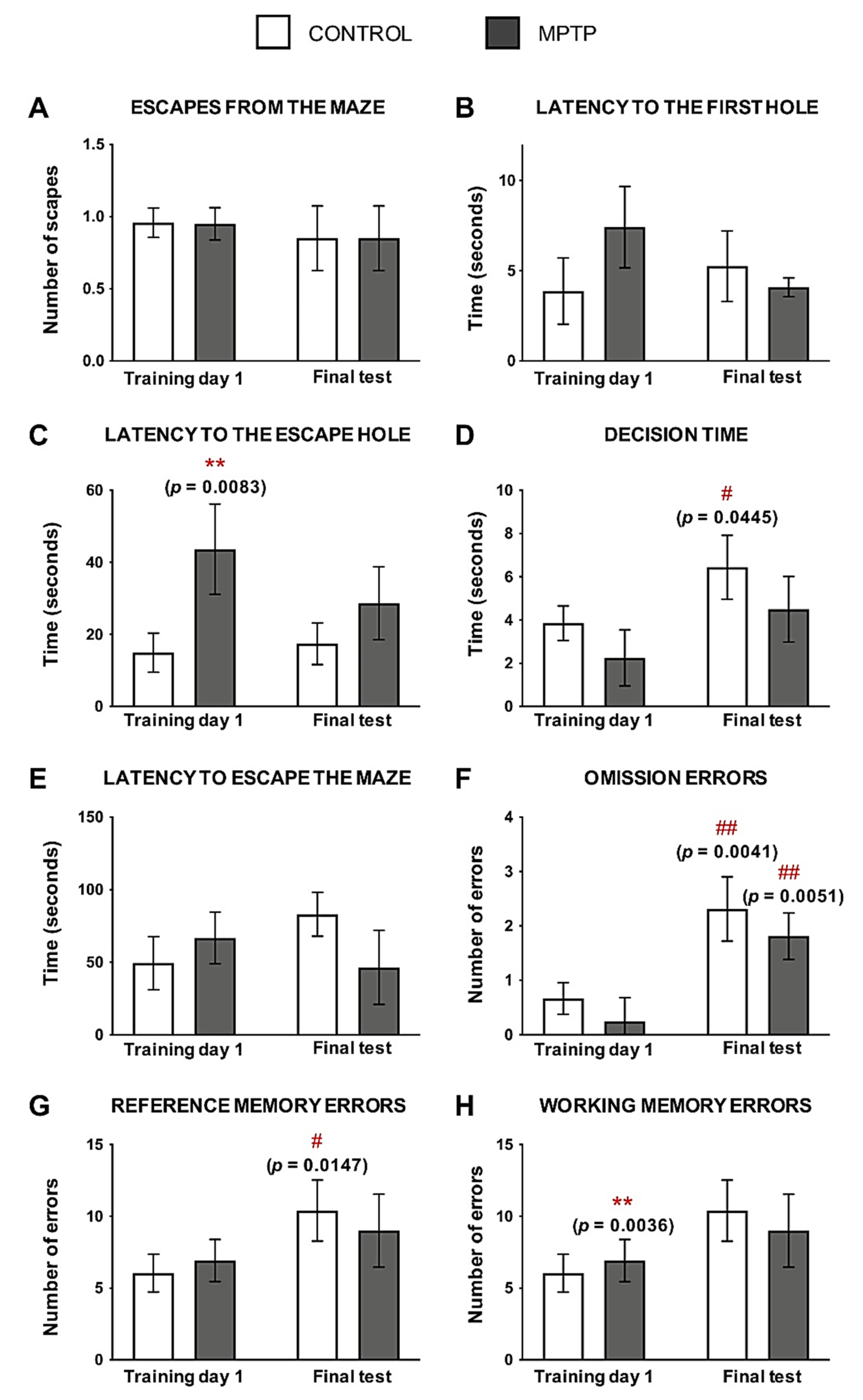
2.4. O. degus Intoxicated with MPTP Show Cognitive Impairment
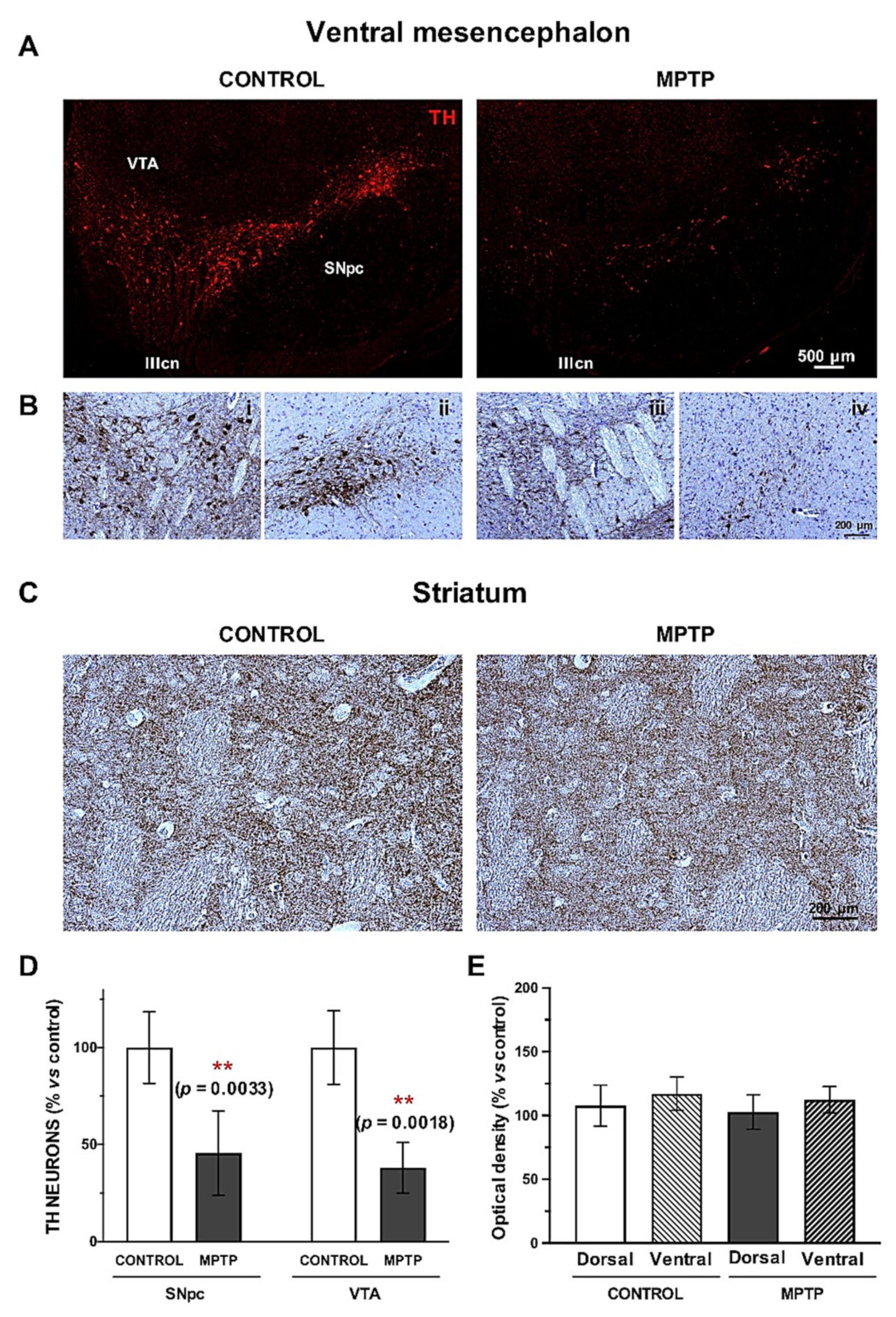
2.5. Dopaminergic Alterations in the MPTP-Intoxicated Animals
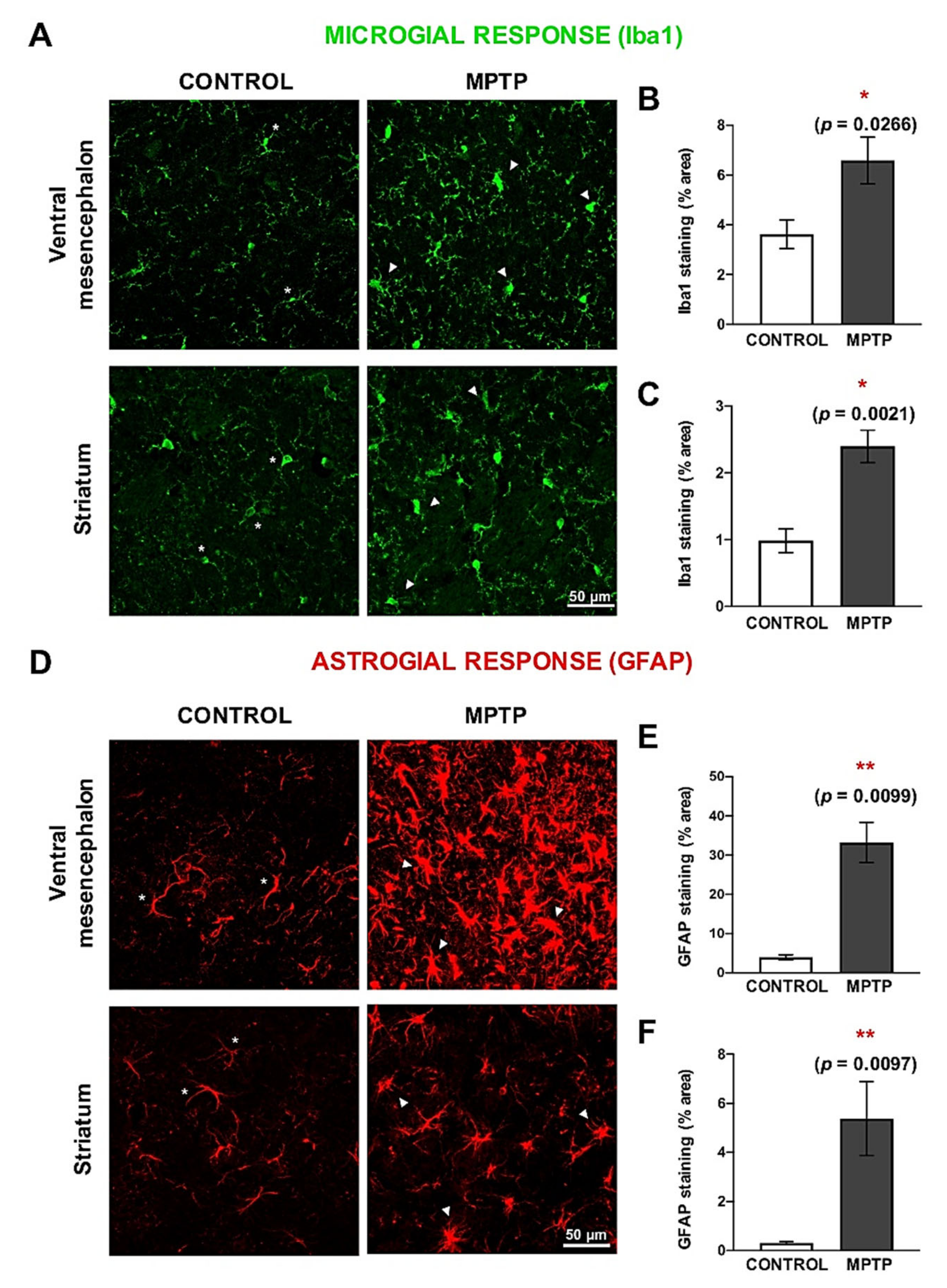
2.6. Increase of Neuroinflammatory Cells in the MPTP-Intoxicated O. degus
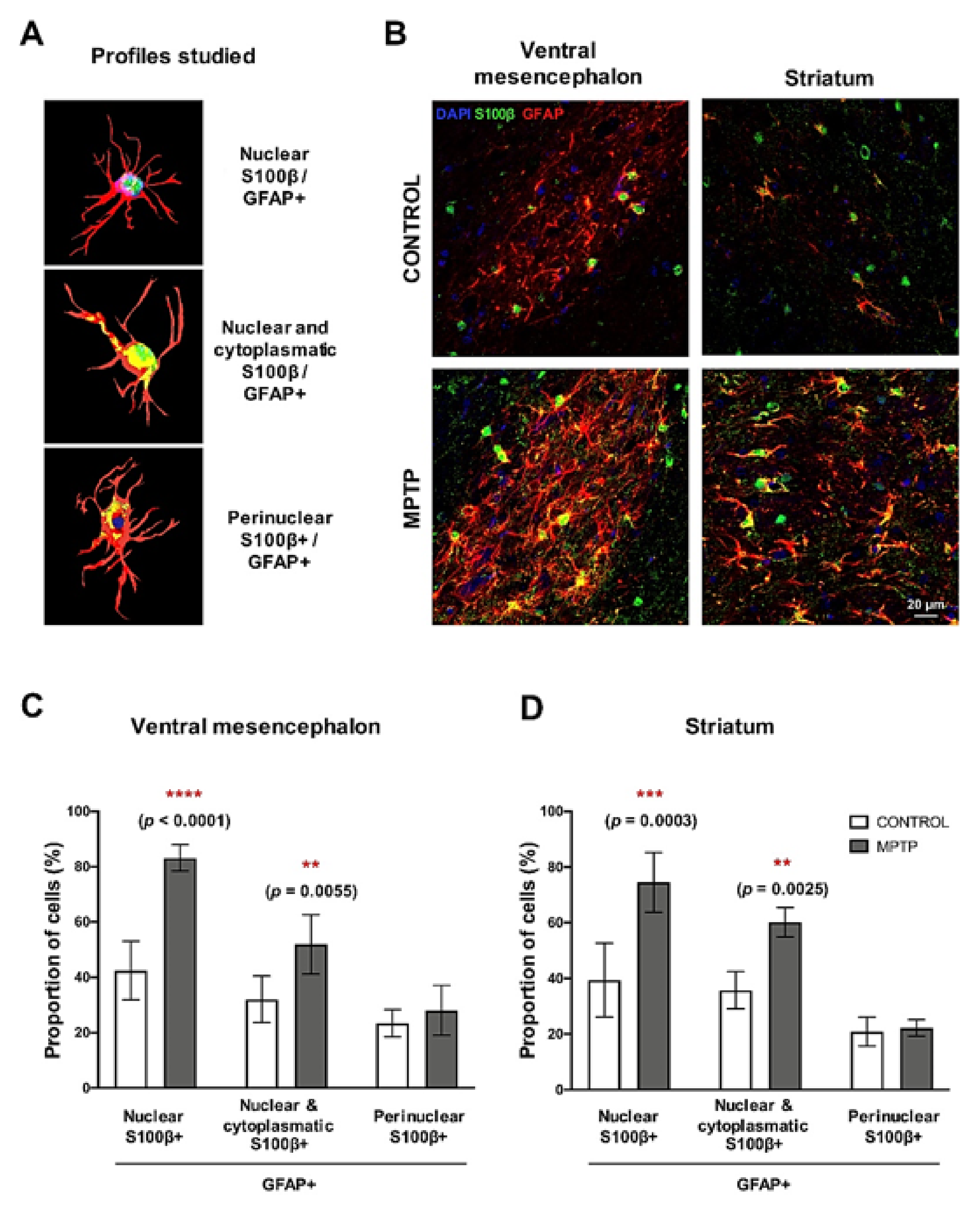
2.7. MPTP Promotes Reactive Astrogliosis in the O. degus
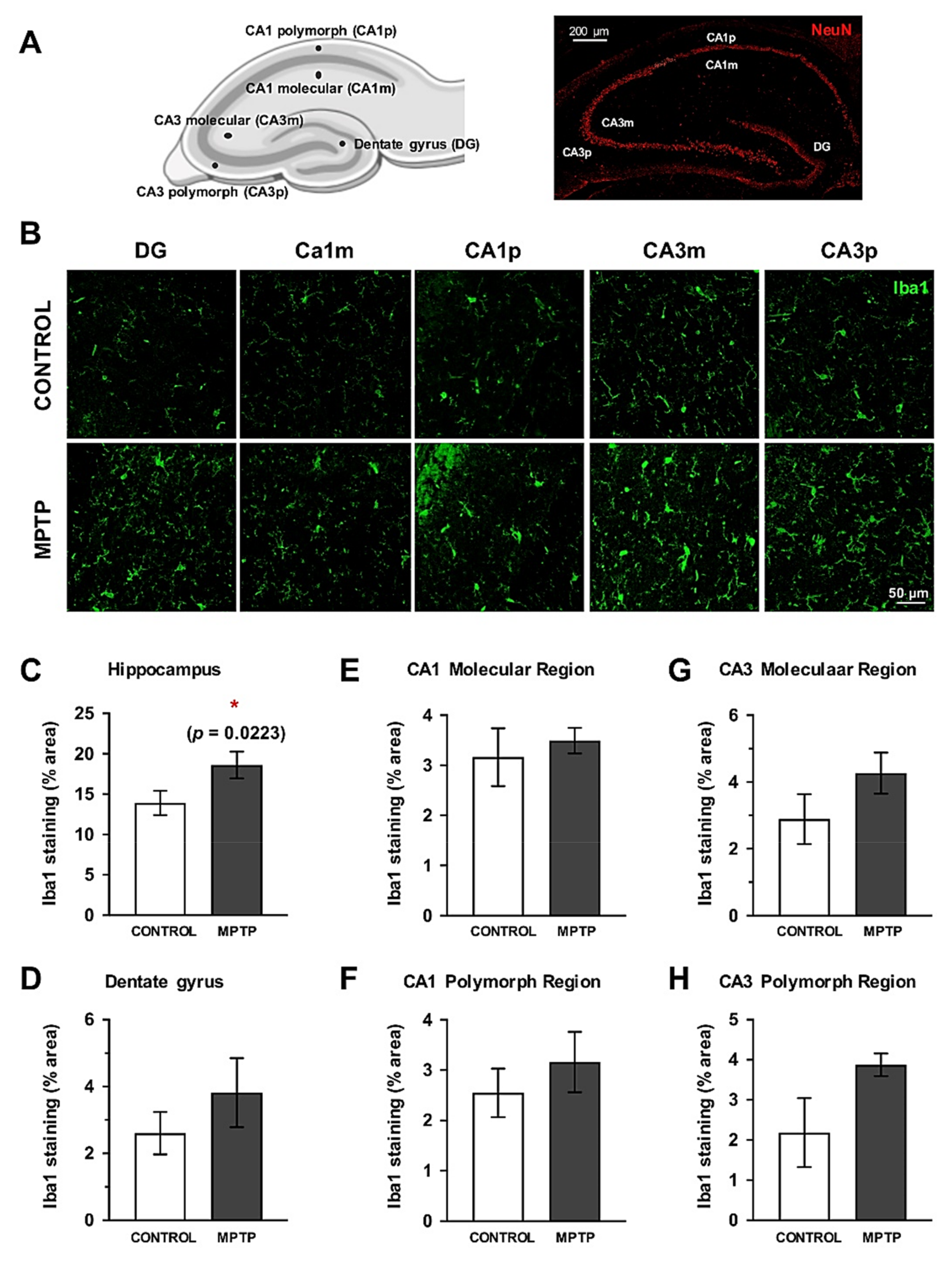
2.8. Increase of Microglial Cells in the Hippocampus
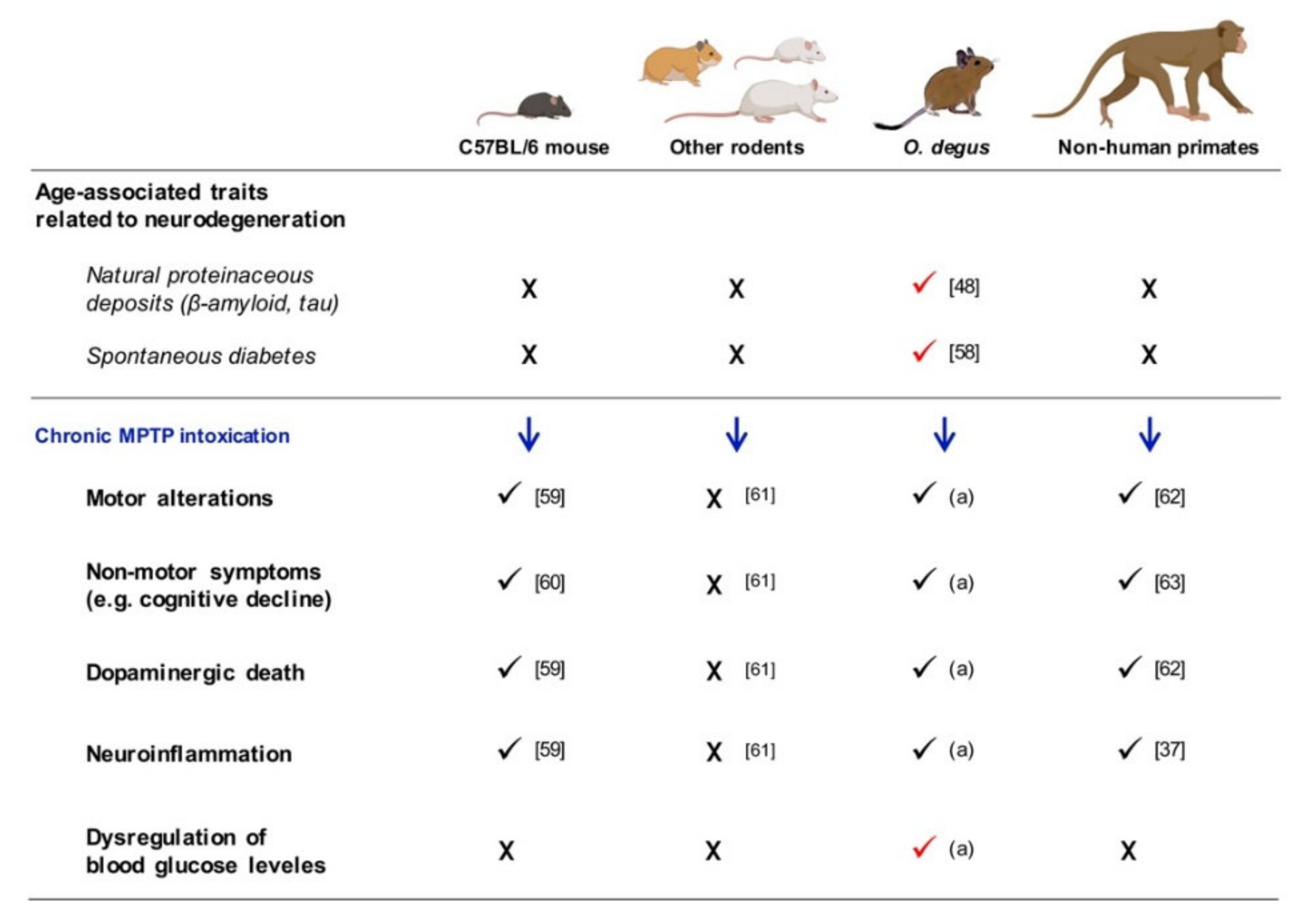
3. Discussion
Limitations of the Study
4. Materials and Methods
4.1. Animals
4.2. MPTP Administration and Quarantine Period
4.3. Behavioral Tests
- Open field test. The open field test is one of the most common locomotor tests for experimental PD because rodents with motor deficits or cognitive decline can show an impaired ability to move and explore the arena [64]. The chamber consisted of a transparent open Plexiglas arena (44.5 cm wide × 44.5 cm long × 40 cm high) located inside a system of two sets of 16 infrared photocells (SAI Electronics & Engineering division, University of Murcia, Murcia, Spain). Horizontal locomotor activity and vertical movements were measured by the registration of the beam breaks, which were recorded and transmitted to a computerized system [65]. Every animal was placed in the center of the arena, and its activity was recorded during 180 s. The chamber was cleaned with 70% ethanol after each trial. The test was done taking the following MPTP injections as a time reference: baseline (prior to the first MPTP injection), 48 h after the fifth MPTP injection (cumulative dose of 50 mg/kg), the eighth MPTP injection (cumulative dose of 80 mg/kg) and the last MPTP injection (cumulative dose of 100 mg/kg). Control animals performed the test the same days in order to have comparable measures.
- Barnes Maze. This cognitive test is performed in a Plexiglass circular platform (160 cm diameter) that contains 18 holes: 17 of them are blocked with a mesh, and one is the exit of the maze to the animal’s cage. The test consists of three phases: (i) the habituation day (day 0), in which the animal is kindly taught by the investigator which is the path to escape the maze from the center of the platform; (ii) the training days (days 1–7), in which the animal is placed in the center of the platform and has 4 min to escape the maze (this is repeated for 4 rounds); and (iii) the retention day (“final test”, day 8) that follows the same protocol as the training days. During the training and the retention days, we measured several parameters to evaluate spatial memory. For a detailed description of the protocol, see [66]. Barnes Maze was performed before the last MPTP injection.
4.4. Glucose Measurement
4.5. Samples Obtention and Brain Tissue Preparation
4.6. Immunohistochemistry and Immunofluorescence
4.7. Image Caption and Stereological Analysis
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Hayes, M.T. Parkinson’s disease and parkinsonism. Am. J. Med. 2019, 132, 802–807. [Google Scholar] [CrossRef]
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Prim. 2017, 3, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Cuenca, L.; Gil-Martinez, A.L.; Cano-Fernandez, L.; Sanchez-Rodrigo, C.; Estrada, C.; Fernandez-Villalba, E.; Herrero, M.T. Parkinson’s disease: A short story of 200 years. Histol. Histopathol. 2019, 34, 573–591. [Google Scholar] [CrossRef]
- Obeso, J.A.; Stamelou, M.; Goetz, C.G.; Poewe, W.; Lang, A.E.; Weintraub, D.; Burn, D.; Halliday, G.M.; Bezard, E.; Przedborski, S.; et al. Present, and future of Parkinson ’s disease: A special essay on the 200th anniversary of the shaking palsy introduction I. Mov. Disord. 2017, 32, 1264–1310. [Google Scholar] [CrossRef]
- Zhu, G.; Li, J.; He, L.; Wang, X.; Hong, X. MPTP-induced changes in hippocampal synaptic plasticity and memory are prevented by memantine through the BDNF-TrkB pathway. Br. J. Pharmacol. 2015, 172, 2354–2368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grandi, L.C.; Di Giovanni, G.; Galati, S. Animal models of early-stage Parkinson’s disease and acute dopamine deficiency to study compensatory neurodegenerative mechanisms. J. Neurosci. Methods 2018, 308, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Hamadjida, A.; Frouni, I.; Kwan, C.; Huot, P. Classic animal models of Parkinson’s disease: A historical perspective. Behav. Pharmacol. 2019, 30, 291–310. [Google Scholar] [CrossRef]
- Kopin, I.J.; Markey, S.P. MPTP toxicity: Implications for research in Parkinson’s disease. Annu. Rev. Neurosci. 1988, 11, 81–96. [Google Scholar] [CrossRef]
- Gil-Martínez, A.L.; Cuenca, L.; Estrada, C.; Sánchez-Rodrigo, C.; Fernández-Villalba, E.; Herrero, M.T. Unexpected exacerbation of neuroinflammatory response after a combined therapy in old Parkinsonian mice. Front. Cell. Neurosci. 2018, 12. [Google Scholar] [CrossRef]
- Huang, D.; Xu, J.; Wang, J.; Tong, J.; Bai, X.; Li, H.; Wang, Z.; Huang, Y.; Wu, Y.; Yu, M.; et al. dynamic changes in the nigrostriatal pathway in the MPTP mouse model of Parkinson’s disease. Parkinsons Dis. 2017, 9349487. [Google Scholar] [CrossRef] [PubMed]
- Annese, V.; Barcia, C.; Ros-Bernal, F.; Gómez, A.; Ros, C.M.; De Pablos, V.; Fernández-Villalba, E.; De Stefano, M.E.; Herrero, M.-T. Evidence of oligodendrogliosis in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinsonism. Neuropathol. Appl. Neurobiol. 2013, 39, 132–143. [Google Scholar] [CrossRef] [PubMed]
- Tarragon, E.; Lopez, D.; Estrada, C.; Ana, G.-C.; Schenker, E.; Pifferi, F.; Bordet, R.; Richardson, J.C.; Herrero, M.-T. Octodon degus: A model for the cognitive impairment associated with Alzheimer’s disease basic research in Alzheimer’s disease. CNS Neurosci. Ther. 2013, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Hurley, M.J.; Deacon, R.M.J.; Beyer, K.; Ioannou, E.; Ibáñez, A.; Teeling, J.L.; Cogram, P. The long-lived Octodon degus as a rodent drug discovery model for Alzheimer’s and other age-related diseases. Pharmacol. Ther. 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuenca-Bermejo, L.; Pizzichini, E.; Gonzalez-Cuello, A.M.; De Stefano, M.E.; Fernandez-Villalba, E.; Herrero, M.T. Octodon degus: A natural model of multimorbidity for ageing research. Ageing Res. Rev. 2020, 64, 101204. [Google Scholar] [CrossRef]
- Colonnello, V.; Iacobucci, P.; Fuchs, T.; Newberry, R.C.; Panksepp, J. Octodon degus. A useful animal model for social-affective neuroscience research: Basic description of separation distress, social attachments and play. Neurosci. Biobehav. Rev. 2011, 35, 1854–1863. [Google Scholar] [CrossRef]
- Refinetti, R.; Kenagy, G.J. Diurnally active rodents for lab oratory research. Lab. Anim. 2018, 52, 577–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gil-Martínez, A.-L.; Cuenca, L.; Sánchez, C.; Estrada, C.; Fernández-Villalba, E.; Herrero, M.T. Effect of NAC treatment and physical activity on neuroinflammation in subchronic Parkinsonism; is physical activity essential? J. Neuroinflammation 2018, 15, 328. [Google Scholar] [CrossRef] [PubMed]
- Chia, S.J.; Tan, E.K.; Chao, Y.X. Historical perspective: Models of Parkinson’s disease. Int. J. Mol. Sci. 2020, 21, 2464. [Google Scholar] [CrossRef] [Green Version]
- Inestrosa, N.C.; Reyes, A.E.; Chacón, M.A.; Cerpa, W.; Villalón, A.; Montiel, J.; Merabachvili, G.; Aldunate, R.; Bozinovic, F.; Aboitiz, F. Human-like rodent amyloid-β-peptide determines Alzheimer pathology in aged wild-type Octodon degu. Neurobiol. Aging 2005, 26, 1023–1028. [Google Scholar] [CrossRef]
- Spear, G.S.; Caple, M.V. The Pancreas in the degu. Exp. Mol. Pathol. 1984, 40, 295–310. [Google Scholar] [CrossRef]
- Luchtman, D.W.; Shao, D.; Song, C. Behavior, neurotransmitters and inflammation in three regimens of the MPTP mouse model of Parkinson’s disease. Physiol. Behav. 2009, 98, 130–138. [Google Scholar] [CrossRef]
- Deguil, J.; Chavant, F.; Lafay-Chebassier, C.; Pérault-Pochat, M.-C.; Fauconneau, B.; Pain, S. Neuroprotective effect of PACAP on translational control alteration and cognitive decline in MPTP Parkinsonian mice. Neurotox. Res. 2010, 17, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Riachi, N.J.; Lamanna, J.C.; I Harik, S. Entry of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine into the rat brain. J. Pharmacol. Exp. Ther. 1989, 249, 744–748. [Google Scholar]
- Pérez-Otaño, I.; Oset, C.; Luquin, M.; Herrero, M.; Obeso, J.; Del Río, J. MPTP-induced parkinsonism in primates: Pattern of striatal dopamine loss following acute and chronic administration. Neurosci. Lett. 1994, 175, 121–125. [Google Scholar] [CrossRef]
- Vezoli, J.; Fifel, K.; Leviel, V.; Dehay, C.; Kennedy, H.; Cooper, H.M.; Gronfier, C.; Procyk, E. Early presymptomatic and long-term changes of rest activity cycles and cognitive behavior in a MPTP-monkey model of Parkinson’s disease. PLoS ONE 2011, 6, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barcia, C.; Bahillo, A.S.; Fernández-Villalba, E.; Bautista, V.; Poza, M.P.Y.; Fernández-Barreiro, A.; Hirsch, E.C.; Herrero, M.-T. Evidence of active microglia in substantia nigra pars compacta of parkinsonian monkeys 1 year after MPTP exposure. Glia 2004, 46, 402–409. [Google Scholar] [CrossRef]
- Mounayar, S.; Boulet, S.; Tandé, D.; Jan, C.; Pessiglione, M.; Hirsch, E.C.; Féger, J.; Savasta, M.; François, C.; Tremblay, L. A new model to study compensatory mechanisms in MPTP-treated monkeys exhibiting recovery. Brain 2007, 130, 2898–2914. [Google Scholar] [CrossRef]
- Schneider, J.S.; Schroeder, J.A.; Rothblat, D.S. Differential recovery of sensorimotor function in GM1 ganglioside-treated vs. spontaneously recovered MPTP-treated cats: Partial striatal dopaminergic reinnervation vs. neurochemical compensation. Brain Res. 1998, 813, 82–87. [Google Scholar] [CrossRef]
- Gagnon, D.; Eid, L.; Coudé, D.; Whissel, C.; Di Paolo, T.; Parent, M. Evidence for sprouting of dopamine and serotonin axons in the pallidum of parkinsonian monkeys. Front. Neuroanat. 2018, 12, 1–14. [Google Scholar] [CrossRef]
- Rothblat, D.S.; Schneider, J.S. Spontaneous functional recovery from parkinsonism is not due to reinnervation of the dorsal striatum by residual dopaminergic neurons. Brain Res. Bull. 1994, 34, 309–312. [Google Scholar] [CrossRef]
- Blesa, J.; Trigo-Damas, I.; Dileone, M.; del Rey, N.L.G.; Hernandez, L.F.; Obeso, J.A. Compensatory mechanisms in Parkinson’s disease: Circuits adaptations and role in disease modification. Exp. Neurol. 2017, 298, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Monje, M.H.G.; Blesa, J.; García-Cabezas, M.Á.; Obeso, J.A.; Cavada, C. Changes in thalamic dopamine innervation in a progressive Parkinson’s disease model in monkeys. Mov. Disord. 2020, 35, 419–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bezard, E.; Gross, C.E.; Brotchie, J.M. Presymptomatic compensation in Parkinson’s disease is not dopamine-mediated. Trends Neurosci. 2004, 26, 215–221. [Google Scholar] [CrossRef]
- Obeso, J.A.; Rodriguez-Oroz, M.C.; Lanciego, J.L.; Rodriguez Diaz, M.; Bezard, E.; Gross, C.E.; Brotchie, J.M. How does Parkinson’s disease begin? The role of compensatory mechanisms (multiple letters). Trends Neurosci. 2004, 27, 125–127. [Google Scholar] [CrossRef] [Green Version]
- Langston, J.W. The MPTP story. J. Parkinsons Dis. 2017, 7, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, M.; Hirsch, E.; Kastner, A.; Ruberg, M.; Luquin, M.; Laguna, J.; Javoy-Agid, F.; Obeso, J.; Agid, Y. Does neuromelanin contribute to the vulnerability of catecholaminergic neurons in monkeys intoxicated with MPTP? Neuroscience 1993, 56, 499–511. [Google Scholar] [CrossRef]
- Blesa, J.; Vila, M. Parkinson disease, substantia nigra vulnerability, and calbindin expression: Enlightening the darkness? Mov. Disord. 2019, 34, 161–163. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.; Huang, J.; Zheng, Y.; Ding, Y.; Li, S.; Zhang, J.; Lyu, J.; Zeng, Q. UCP2 silencing aggravates mitochondrial dysfunction in astrocytes under septic conditions. Mol. Med. Rep. 2019, 20, 4459–4466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paradis, E.; Clavel, S.; Bouillaud, F.; Ricquier, D.; Richard, D. Uncoupling protein 2: A novel player in neuroprotection. Trends Mol. Med. 2003, 9, 522–525. [Google Scholar] [CrossRef]
- Heman, P.; Barcia, C.; Gómez, A.; Ros, C.M.; Ros-Bernal, F.; Yuste, J.E.; De Pablos, V.; Fernandez-Villalba, E.; Toledo-Cárdenas, M.R.; Herrero, M.T. Nigral degeneration correlates with persistent activation of cerebellar Purkinje cells in MPTP-treated monkeys. Histol. Histopathol. 2012, 1, 89–94. [Google Scholar] [CrossRef]
- Pewzner-Jung, Y.; Joseph, T.; Blumenreich, S.; Vardi, A.; Ferreira, N.S.; Cho, S.M.; Eilam, R.; Tsoory, M.; Biton, I.E.; Brumfeld, V.; et al. Brain pathology and cerebellar purkinje cell loss in a mouse model of chronic neuronopathic Gaucher disease. Prog. Neurobiol. 2020, 197, 101939. [Google Scholar] [CrossRef]
- Fuzzati-Armentero, M.T.; Cerri, S.; Blandini, F. Peripheral-central neuroimmune crosstalk in Parkinson’s disease: What do patients and animal models tell us? Front. Neurol. 2019, 10, 1–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Martinez, L.; Maccioni, R.B.; Andrade, V.; Navarrete, L.P.; Pastor, M.G.; Ramos-Escobar, N. Neuroinflammation as a common feature of neurodegenerative disorders. Front. Pharmacol. 2019, 10, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Kam, T.-I.; Hinkle, J.T.; Dawson, T.M.; Dawson, V.L. Microglia and astrocyte dysfunction in parkinson’s disease. Neurobiol. Dis. 2020, 144, 105028. [Google Scholar] [CrossRef] [PubMed]
- Angelopoulou, E.; Paudel, Y.N.; Piperi, C. Emerging role of S100B protein implication in Parkinson’s disease pathogenesis. Cell. Mol. Life Sci. 2020, 78, 1445–1453. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, Y.; Kurosaki, R.; Watanabe, H.; Michimata, M.; Matsubara, M.; Imai, Y.; Araki, T. Expression of S-100 protein is related to neuronal damage in MPTP-treated mice. Glia 2003, 42, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Donato, R.; Sorci, G.; Riuzzi, F.; Arcuri, C.; Bianchi, R.; Brozzi, F.; Tubaro, C.; Giambanco, I. S100B’s double life: Intracellular regulator and extracellular signal. Biochim. Biophys. Acta Bioenerg. 2009, 1793, 1008–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarragon, E.; Lopez, D.; Estrada, C.; Gonzalez-Cuello, A.; Cros, E.T.; Lamberty, Y.; Pifferi, F.; Cella, M.; Canovi, M.; Guiso, G.; et al. Memantine prevents reference and working memory impairment caused by sleep deprivation in both young and aged Octodon degus. Neuropharmacology 2014, 85, 206–214. [Google Scholar] [CrossRef]
- Knierim, J.J. The hippocampus. Curr. Biol. 2015, 25, R1116–R1121. [Google Scholar] [CrossRef] [Green Version]
- Singh, S.; Mishra, A.; Tiwari, V.; Shukla, S. Enhanced neuroinflammation and oxidative stress are associated with altered hippocampal neurogenesis in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine treated mice. Behav. Pharmacol. 2019, 30, 688–698. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Huang, Y.; Chen, Y.; Zhuang, Y.; Behnisch, T. MPTP modulates hippocampal synaptic transmission and activity-dependent synaptic plasticity via dopamine receptors. J. Neurochem. 2012, 122, 582–593. [Google Scholar] [CrossRef]
- Cataldi, S.; Arcuri, C.; Hunot, S.; Légeron, F.-P.; Mecca, C.; Garcia-Gil, M.; Lazzarini, A.; Codini, M.; Beccari, T.; Tasegian, A.; et al. Neutral sphingomyelinase behaviour in hippocampus neuroinflammation of MPTP-induced mouse model of Parkinson’s disease and in embryonic hippocampal cells. Mediat. Inflamm. 2017, 2017, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Maluf, F.C.; Feder, D.; Carvalho, A.A.D.S. Analysis of the relationship between type II Diabetes Mellitus and Parkinson’s disease: A systematic review. Parkinsons Dis. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cisternas, P.; Zolezzi, J.M.; Lindsay, C.; Rivera, D.S.; Martinez, A.; Bozinovic, F.; Inestrosa, N.C. New insights into the spontaneous human Alzheimer’s disease-like model Octodon degus: Unraveling amyloid-β peptide aggregation and age-related amyloid pathology. J. Alzheimer’s Dis. 2018, 66, 1145–1163. [Google Scholar] [CrossRef] [PubMed]
- Bourdenx, M.; Dovero, S.; Thiolat, M.-L.; Bezard, E.; Dehay, B. Lack of spontaneous age-related brain pathology in Octodon degus: A reappraisal of the model. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Cadillac, J.M.; Rush, H.G.; Sigler, R.E. Polycystic and chronic kidney disease in a young degu (Octodon degus). Contemp. Top Lab. Anim. Sci. 2003, 42, 43–45. [Google Scholar] [PubMed]
- Švara, T.; Gombač, M.; Poli, A.; Račnik, J.; Zadravec, M. Spontaneous tumors and non-neoplastic proliferative lesions in pet degus (Octodon degus). Vet. Sci. 2020, 7, 32. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, S.; Ito, T.; He, X.J.; Uchida, K.; Nakayama, H. Resistance of the golden hamster to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-neurotoxicity is not only related with low levels of cerebral monoamine oxidase-B. Exp. Toxicol. Pathol. 2013, 65, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Zuddas, A.; Fascetti, F.; Corsini, G.U.; Piccardi, M.P. In brown Norway rats, MPP+ is accumulated in the nigrostriatal dopaminergic terminals but it is not neurotoxic: A model of natural resistance to MPTP toxicity. Exp. Neurol. 1994, 127, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Meredith, G.E.; Rademacher, D.J. MPTP mouse models of Parkinson’s disease: An update. N. Libr. World 2016, 96, 16–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Stefano, M.E.; Herrero, M.T. The multifaceted role of metalloproteinases in physiological and pathological conditions in embryonic and adult brains. Prog. Neurobiol. 2017, 155, 36–56. [Google Scholar] [CrossRef] [PubMed]
- Annese, V.; Herrero, M.-T.; Di Pentima, M.; Gomez, A.; Lombardi, L.; Ros, C.M.; De Pablos, V.; Fernandez-Villalba, E.; De Stefano, M.E. Metalloproteinase-9 contributes to inflammatory glia activation and nigro-striatal pathway degeneration in both mouse and monkey models of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced Parkinsonism. Brain Struct. Funct. 2015, 220, 703–727. [Google Scholar] [CrossRef]
- Jackson-Lewis, V.; Przedborski, S. Protocol for the MPTP mouse model of Parkinson’s disease. Nat. Protoc. 2007, 2, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Hutter-Saunders, J.A.L.; Gendelman, H.E.; Mosley, R.L. Murine motor and behavior functional evaluations for acute 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) intoxication. J. Neuroimmune Pharmacol. 2012, 7, 279–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melo-Thomas, L.; Gil-Martínez, A.; Cuenca, L.; Estrada, C.; Gonzalez-Cuello, A.; Schwarting, R.; Herrero, M. Electrical stimulation or MK-801 in the inferior colliculus improve motor deficits in MPTP-treated mice. Neurotoxicology 2018, 65, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Estrada, C.; López, D.; Conesa, A.; Fernandez-Gomez, F.J.; Gonzalez-Cuello, A.; Toledo, F.; Túnez, I.; Blin, O.; Bordet, R.; Richardson, J.C.; et al. Cognitive impairment after sleep deprivation rescued by transcranial magnetic stimulation application in Octodon degus. Neurotox. Res. 2015, 28, 361–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods. 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cuenca-Bermejo, L.; Pizzichini, E.; Gonçalves, V.C.; Guillén-Díaz, M.; Aguilar-Moñino, E.; Sánchez-Rodrigo, C.; González-Cuello, A.-M.; Fernández-Villalba, E.; Herrero, M.T. A New Tool to Study Parkinsonism in the Context of Aging: MPTP Intoxication in a Natural Model of Multimorbidity. Int. J. Mol. Sci. 2021, 22, 4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094341
Cuenca-Bermejo L, Pizzichini E, Gonçalves VC, Guillén-Díaz M, Aguilar-Moñino E, Sánchez-Rodrigo C, González-Cuello A-M, Fernández-Villalba E, Herrero MT. A New Tool to Study Parkinsonism in the Context of Aging: MPTP Intoxication in a Natural Model of Multimorbidity. International Journal of Molecular Sciences. 2021; 22(9):4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094341
Chicago/Turabian StyleCuenca-Bermejo, Lorena, Elisa Pizzichini, Valeria C. Gonçalves, María Guillén-Díaz, Elena Aguilar-Moñino, Consuelo Sánchez-Rodrigo, Ana-María González-Cuello, Emiliano Fernández-Villalba, and María Trinidad Herrero. 2021. "A New Tool to Study Parkinsonism in the Context of Aging: MPTP Intoxication in a Natural Model of Multimorbidity" International Journal of Molecular Sciences 22, no. 9: 4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094341