
Oxidative Properties of Polystyrene Nanoparticles with Different Diameters in Human Peripheral Blood Mononuclear Cells (In Vitro Study)


Abstract
:1. Introduction
2. Results
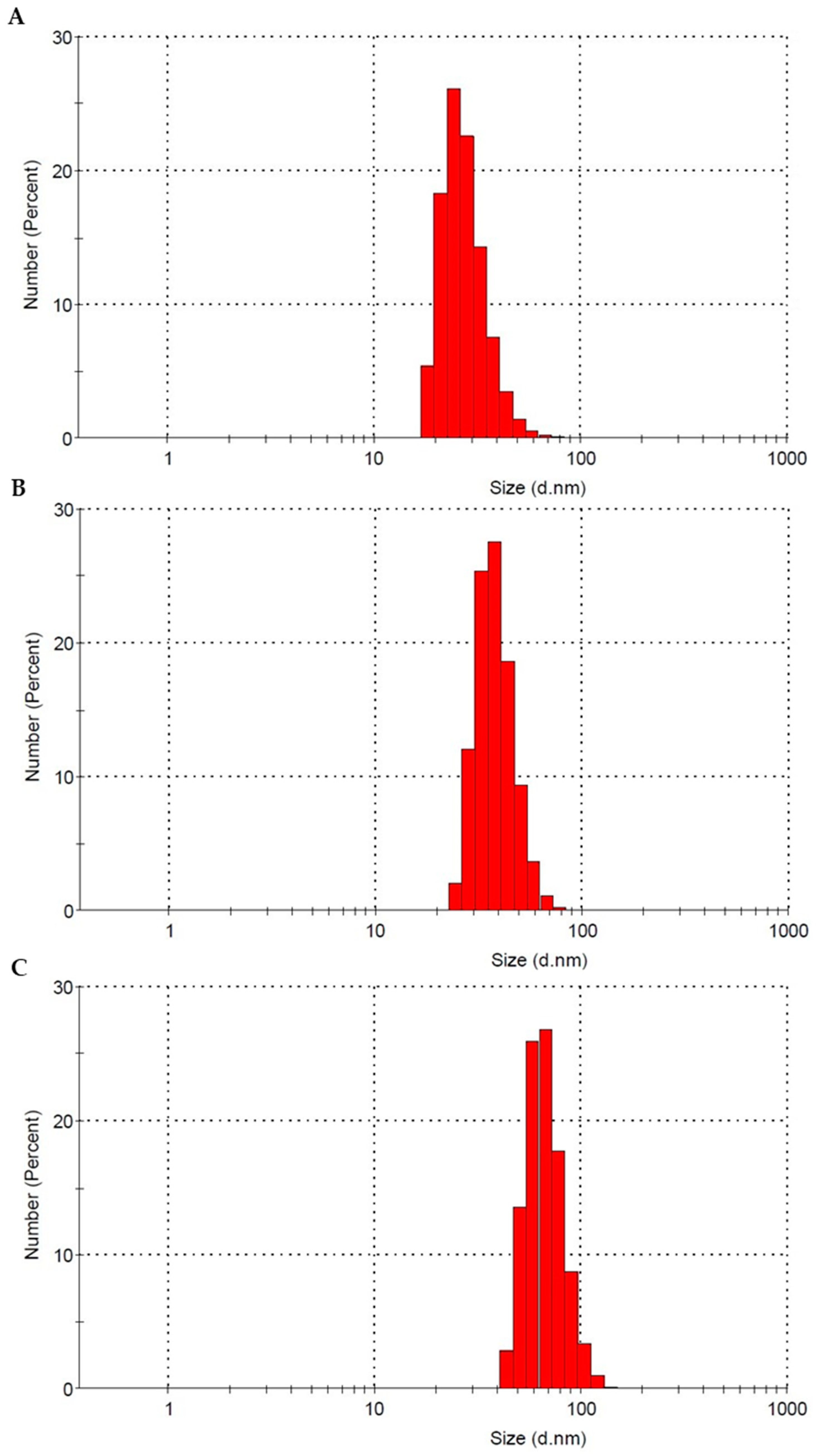
2.1. Characterization of Polystyrene Nanoparticles
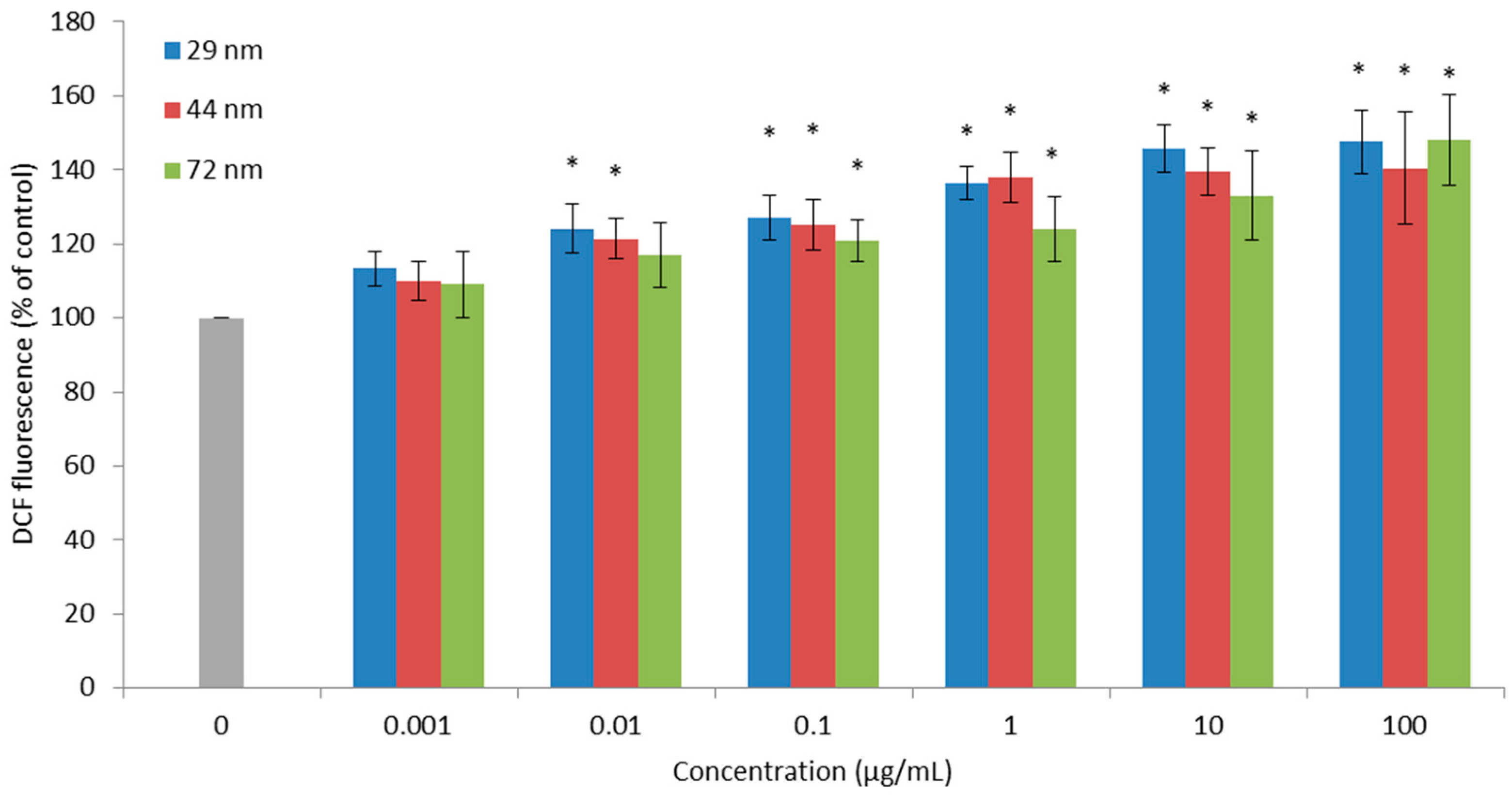
2.2. Reactive Oxygen Species
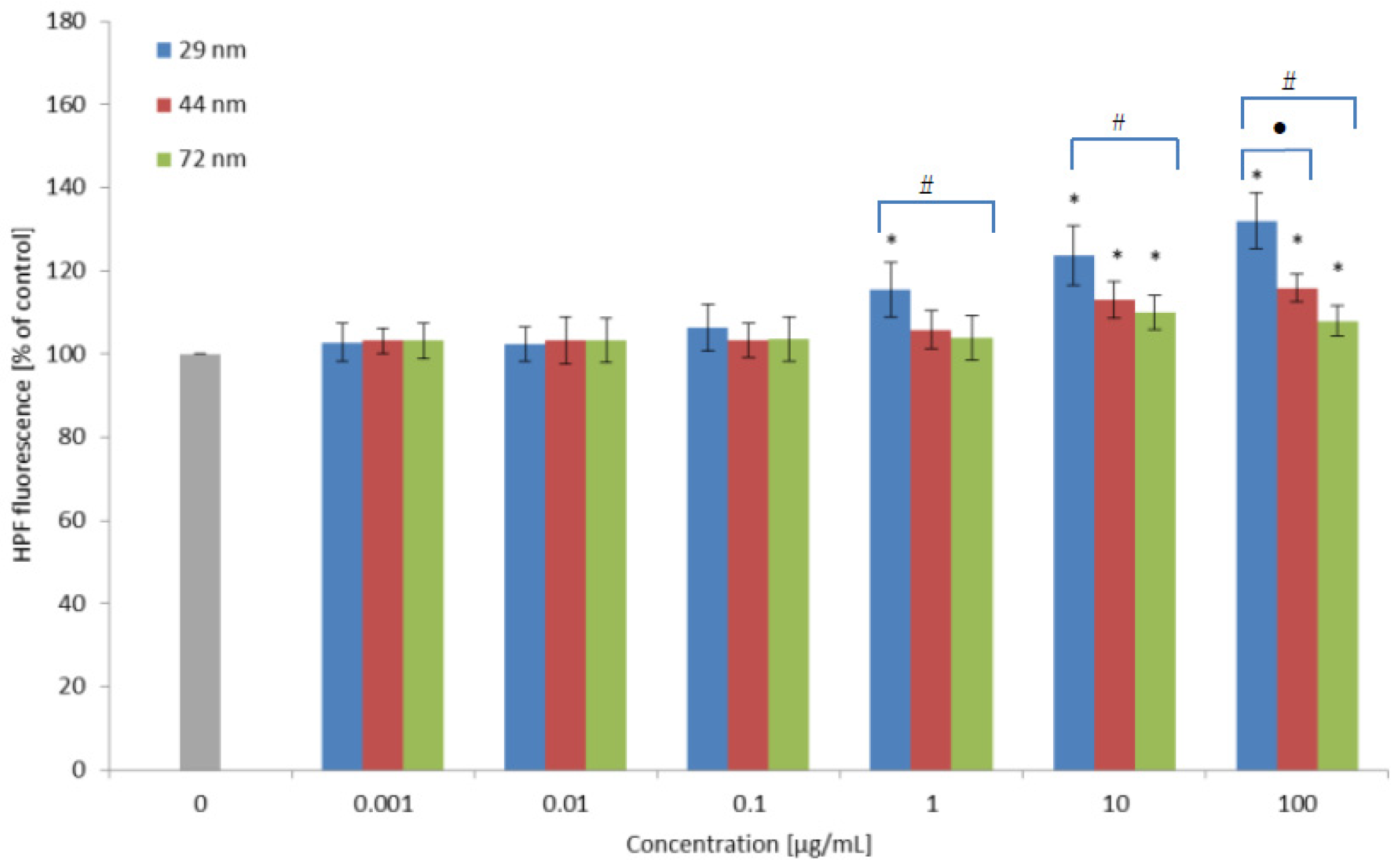
2.3. Hydroxyl Radical Level
2.4. Lipid Peroxidation
2.5. Protein Damages
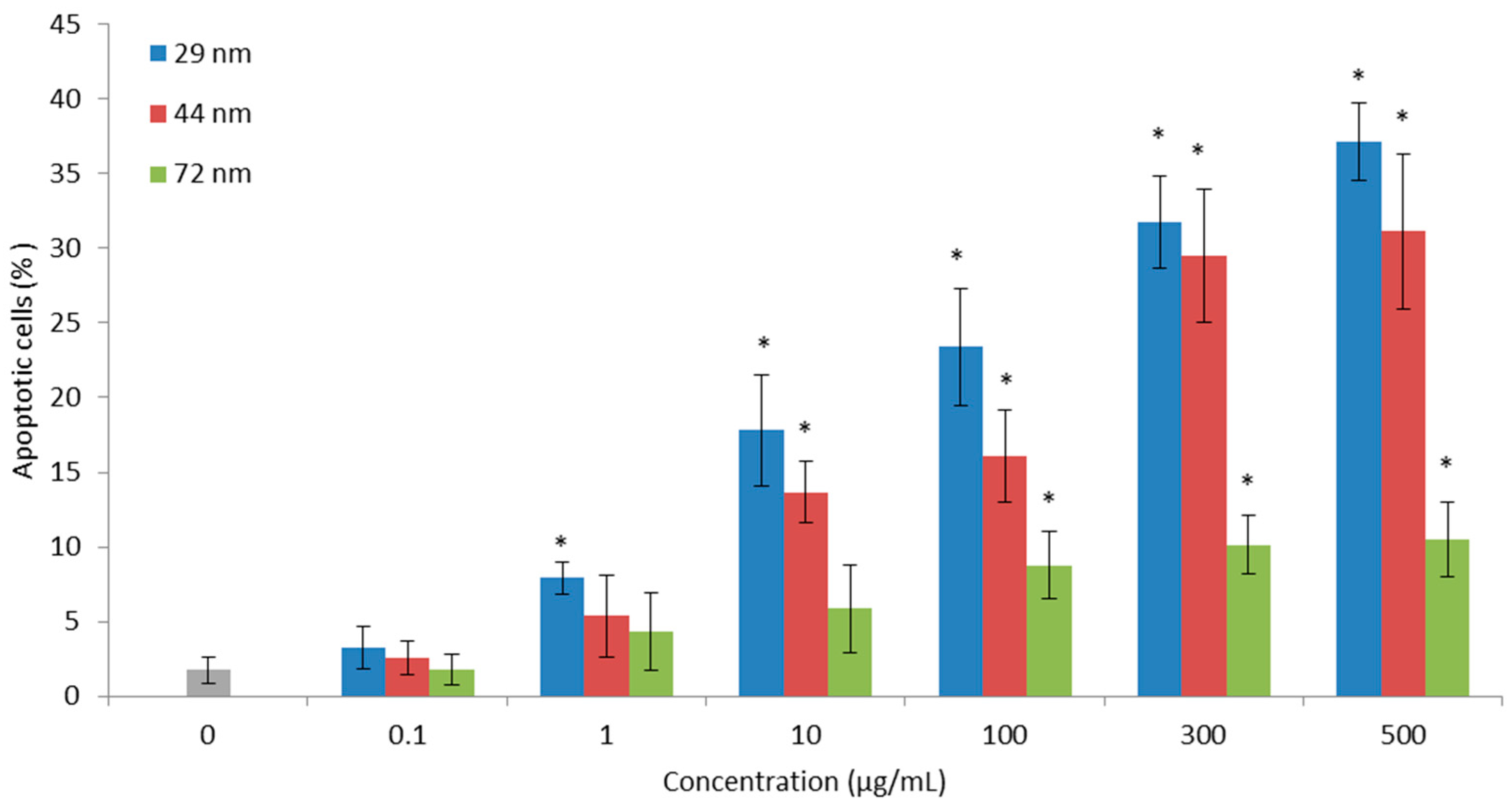
2.6. Cell Apoptosis
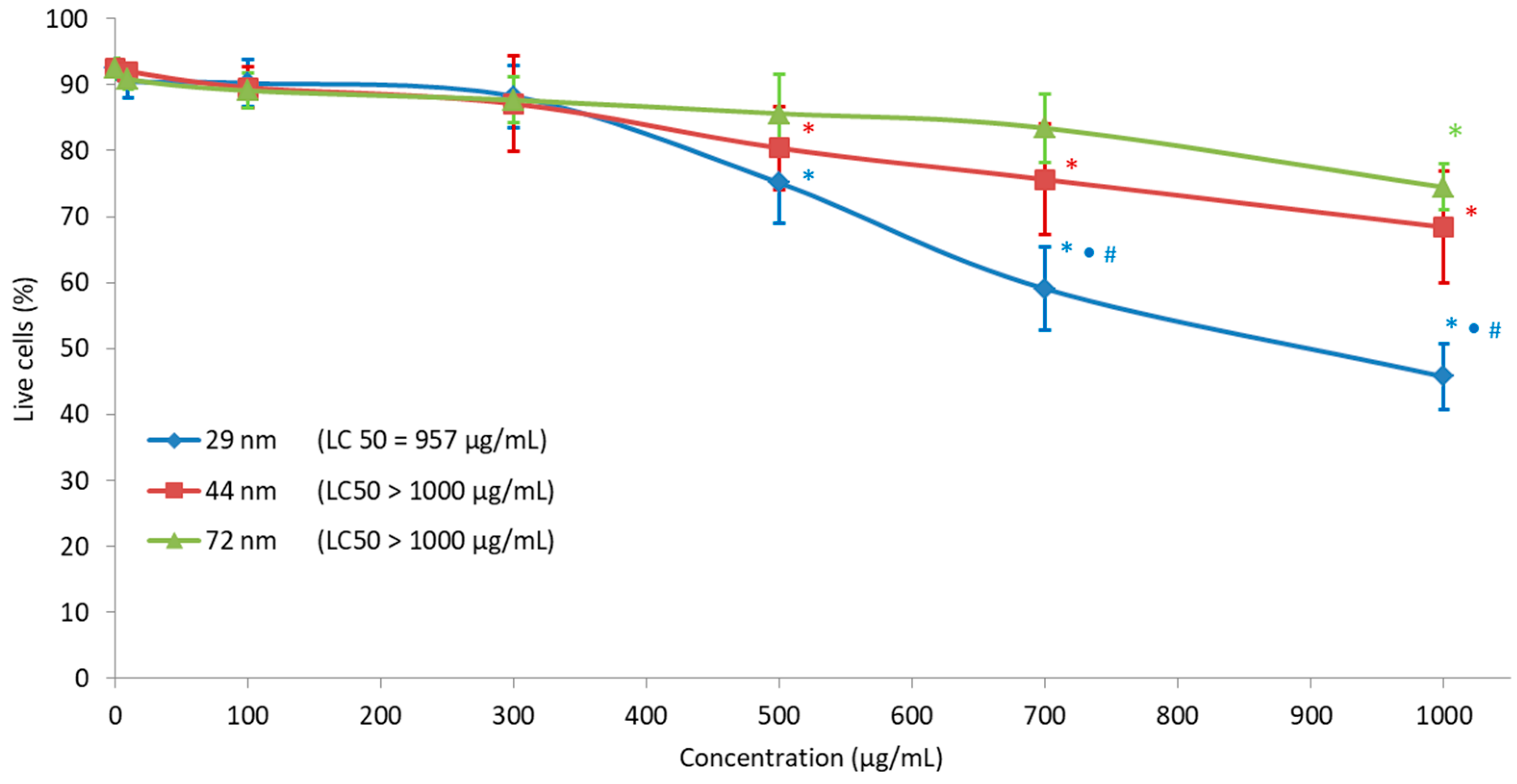
2.7. Cell Viability
3. Discussion
4. Materials and Methods
4.1. Biological Material
4.2. Chemical Standards
4.3. Characterization of Polystyrene Nanoparticles
4.4. Obtaining Leucocytes
4.5. Determination of Reactive Oxygen Species Levels
4.6. Determination of Hydroxyl Radical Level
4.7. Lipid Peroxidation
4.8. Protein Oxidation
4.9. Detection of Apoptosis
4.10. Analysis of Cells Viability of PBMC
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Plastics Europe. Plastics—The Facts. 2017. Available online: https://www.plasticseurope.org/application/files/5715/1717/4180/Plastics_the_facts_2017_FINAL_for_website_one_page.pdf (accessed on 25 March 2021).
- Hesler, M.; Aengenheister, L.; Ellinger, B.; Drexel, R.; Straskraba, S.; Jost, C.; Wagner, S.; Meier, F.; von Briesen, H.; Buchel, C.; et al. Multi-endpoint toxicological assessment of polystyrene nano- and microparticles in different biological models in vitro. Toxicol. Vitro 2019, 61, 104610. [Google Scholar] [CrossRef]
- Hu, M.; Palić, D. Micro- and nano-plastics activation of oxidative and inflammatory adverse outcome pathways. Redox Biol. 2020, 37, 1–16. [Google Scholar] [CrossRef]
- Koelman, A.; Besseling, E.; Shim, W. Nanoplastics in the aquatic environment. Critical review. Mar. Anthrop. Litter 2015, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Efimova, I.; Bagaeva, M.; Bagaev, A.; Kileso, A.; Chubarenko, I. Secondary microplastics generation in the sea swash zone with coarse bottom sediments: Laboratory experiments. Front. Mar. Sci. 2018, 5, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Mason, S.A.; Garneau, D.; Sutton, R.; Chu, Y.; Ehmann, K.; Barnes, J.; Fink, P.; Papazissimos, D.; Rogers, D.L. Microplastic pollution is widely detected in US municipal wastewater treatment plant effluent. Environ. Poll. 2016, 218, 1045–1054. [Google Scholar] [CrossRef]
- Yooeun, C.; Dokyung, K.; Shin, W.K.; Youn-Joo, A. Trophic transfer and individual impact of Nano-sized polystyrene in a four-species freshwater food chain. Sci. Rep. 2018, 8, 1–11. [Google Scholar]
- Mattsson, K.; Hansson, L.; Cedervall, T. Nanoplastics in the aquatic environment. Environ. Sci. Proc. Imp. 2015, 17, 1712–1721. [Google Scholar]
- Kik, K.; Bukowska, B.; Sicińska, P. Polystyrene nanoparticles: Sources, occurrence in the environment, distribution in tissues, accumulation and toxicity to various organisms. Environ. Pollut. 2020, 262, 114297. [Google Scholar] [CrossRef] [PubMed]
- Farely, A.T.; Shaw, C.I. Polystyrene as Hazardous Household Waste. In Household Hazardous Waste Management; IntechOpen: London, UK, 2017; pp. 44–60. [Google Scholar] [CrossRef] [Green Version]
- Eyles, J.; Alpar, H.; Field, W.; Lewis, D.; Keswick, M. The transfer of polystyrene microspheres from the gastrointestinal tract to the circulation after oral administration in the rat. J. Pharm. Pharmacol. 1995, 47, 561–565. [Google Scholar] [CrossRef] [PubMed]
- Jani, P.; Halbert, G.; Langridge, J.; Florence, A. Nanoparticle uptake by the rat gastrointestinal mucosa: Quantification and particle size dependency. J. Pharm. Pharmacol. 1990, 42, 821–826. [Google Scholar] [CrossRef] [PubMed]
- Shang, L.; Nienhaus, K.; Nienhaus, U.G. Engineered nanoparticles interacting with cells: Size matters. J. Nanobiotechnology 2014, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Hussain, N.; Jaitley, V.; Florence, A. Recent advances in the understanding of uptake of microparticulates across the gastrointestinal lymphatics. Adv. Drug Deliv. Rev. 2001, 23, 107–142. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA). Presence of microplastics and nanoplastics in food, with particular focus on seafood. EFSA J. 2016, 14, 1–30. [Google Scholar]
- Smith, M.; Love, C.D.; Rochman, M.C.; Neff, A.R. Microplastics in seafood and the implications for human health. Curr. Environ. Health Rep. 2018, 5, 375–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, S.; Kelly, F. Plastic and human health: A micro issue? Environ. Sci. Technol. 2017, 51, 6634–6647. [Google Scholar] [CrossRef] [PubMed]
- Barboza, A.G.L.; Vethaak, D.A.; Lavorante, O.B.R.B.; Lundebye, K.; Guilhermino, L. Marine microplastics debris: An emerging issue for food security, food safety and human health. Mar. Pollut. Bull. 2018, 133, 336–348. [Google Scholar] [CrossRef] [PubMed]
- Barboza, A.G.L.; Vieira, R.L.; Branco, V.; Carvalho, C.; Guilhermino, L. Microplastics increase mercury bioconcentration in gills and bioaccumulation in the liver, and cause oxidative stress and damage in Dicentrarchuslabrax juveniles. Sci. Rep. 2018, 8, 15655. [Google Scholar] [CrossRef] [Green Version]
- Betzer, O.; Shilo, M.; Opochinsky, R.; Barnoy, E.; Motiei, M.; Okun, E.; Yadid, G.; Popovtzer, R. The effect of nanoparticle size on the ability to cross the blood-brain barrier: An in vivo study. Nanomedicine 2017, 12, 1533–1546. [Google Scholar] [CrossRef]
- Bouwmeester, H.; Hollman, H.C.P.; Peters, B.J.H. Potential health impact of environmentally released micro- and nanoplastics in the human food production chain: Experiences from nanotoxicology. Environ. Sci. Technol. 2015, 49, 8932–8947. [Google Scholar] [CrossRef] [PubMed]
- Wick, P.; Malek, A.; Manser, P.; Meili, D.; Maeder-Althaus, X.; Diener, L.; Diener, P.-A.; Zisch, A.; Krug, F.H.; von Mandach, U. Barrier capacity of human placenta for nanosized materials. Environ. Health Perspect. 2010, 118, 432–436. [Google Scholar] [CrossRef]
- Ragusa, A.; Svelato, A.; Santacroce, C.; Catalano, P.; Notarstefano, V.; Carnevali, O.; Papa, F.; Rongioletti, A.C.M.; Baiocco, F.; Draghi, S.; et al. Plasticenta: First evidence of microplastics in human placenta. Environ. Int. 2021, 146, 106274. [Google Scholar]
- Prata, J. Airborne microplastics: Consequences to human health? Environ. Pollut. 2018, 234, 115–126. [Google Scholar] [CrossRef]
- Mutti, A.; Buzio, C.; Perazzoli, F.; Bergamaschi, E.; Bocchi, M.C.; Selis, L.; Mineo, F.; Franchini, I. Lymphocyte subpopulations in workers exposed occupationally to styrene. Med. Lav. 1992, 83, 167–177. [Google Scholar] [PubMed]
- Thompson, J.; Quigley, J.; Halfpenny, N.; Scott, D.; Hawkins, N. Importance and methods of searching for E-publications ahead of print in systematic reviews. Evid. Based Med. 2016, 21, 55–59. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zhang, Y.; Deng, Y.; Jiang, W.; Zhao, Y.; Geng, J.; Ding, L.; Ren, H.-Q. Uptake and accumulation of polystyrene microplastics in zebrafish (Danio rerio) and Toxic effects in the Liver. Environ. Sci. Technol. 2016, 50, 4054–4060. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Zhang, Y.; Lemos, B.; Ren, H. Tissue accumulation of microplastics in mice and biomarker responses suggest widespread health risks of exposure. Sci. Rep. 2017, 7, 46687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, Y.; Zhang, Y.; Qiao, R.; Bonilla, M.M.; Yang, X.; Ren, H.; Lemos, B. Evidence that microplastics aggravate the toxicity of organophosphorus flame retardants in mice (Mus musculus). J. Hazard. Mater. 2018, 357, 348–354. [Google Scholar] [CrossRef]
- Yang, Y.F.; Chen, C.Y.; Lu, T.H.; Liao, C.M. Toxicity-based toxicokinetic/toxicodynamic assessment for bioaccumulation of polystyrene microplastics in mice. J. Hazard. Mater. 2019, 366, 703–713. [Google Scholar] [CrossRef]
- Lin, W.; Jiang, R.; Hu, S.; Xiao, X.; Wu, J.; Wei, W.; Xiong, Y. Investigating the toxicites of diffrent functionalized polystyrene nanopalastics on Daphnia magna. Ecotoxicol. Environ. Saf. 2019, 180, 509–516. [Google Scholar] [CrossRef]
- Gennari, A.; Ban, M.; Braun, A.; Casati, S.; Corsini, E.; Dastych, J.; Descotes, J.; Hartung, T.; Hooghe-Peters, R.; House, R. The use of in vitro systems for evaluating immunotoxicity: The report and recommendations of an ECVAM workshop. J. Immunotoxicol. 2005, 2, 61–83. [Google Scholar] [CrossRef] [Green Version]
- Tulinska, J.; Kazimirova, A.; Kuricova, M.; Barancokova, M.; Liskova, A.; Neubauerova, E.; Drlickova, M.; Ciampor, F.; Vavra, I.; Bilanicova, D.; et al. Immunotoxicity and genotoxicity testing of PLGA-PEO nanoparticles in human blood cell model. Nanotoxicology 2015, 9, 33–43. [Google Scholar] [CrossRef]
- Dusinska, M.; Tulinska, J.E.; Yamani, N.; Kuricova, M.; Liskova, A.; Rollerova, E.; Rundén-Pran, E.; Smolkova, B. Immunotoxicity, genotoxicity and epigenetic toxicity of nanomaterials: New strategies for toxicity testing? Food Chem. Toxicol. 2017, 109, 797–811. [Google Scholar] [CrossRef]
- Galloway, T. Micro- and Nano-plastic and Human Health. In Marine Anthropogenic Litter; Bergmann, M., Gutow, L., Klages, M., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 343–366. [Google Scholar]
- Dayem, A.A.; Hossain, K.M.; Lee, B.S.; Kim, K.; Saha, K.S.; Yang, M.-G.; Choi, Y.H.; Cho, G.-S. The role of Reactive Oxygen species (ROS) in the biological activities of metallic nanoparticles. Int. J. Mol. Sci. 2017, 18, 120. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Liu, H.; Cui, H.; Chen, B.; Li, L.; Wu, F. Impacts of polystyrene microplastics on the behavior and metabolism in a marine demersal teleost, black rockfish (Sebastes schlegelii). J. Hazard. Mater. 2019, 380, 120861. [Google Scholar] [CrossRef]
- He, Y.; Li, J.; Chen, J.; Miao, X.; Li, G.; He, Q.; Xu, H.; Li, H.; Wei, Y. Cytotoxic effects of polystyrene nanoplastics with different surface functionalization on human HepG2 cells. Sci. Total Environ. 2020, 723, 138180. [Google Scholar] [CrossRef]
- Yousif, E.; Haddad, R. Photodegradation and photostabilization of polymers especially polystyrene: Review. Springerplus 2013, 398, 1–32. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Li, Y.; Perez, E.; Jiang, Q.; Chen, Q.; Yang, J.; Huang, Y.; Yang, Y.; Zhao, Y. Polystyrene nanoplastic induces oxidative stress, immune defense, and glycometabolism change in Daphnia pulex: Application of transcriptome profiling in risk assessment of nanoplastics. J. Hazard. Mater. 2021, 402, 123778. [Google Scholar] [CrossRef]
- Liu, Z.; Huang, Y.; Jiao, Y.; Chen, Q.; Wu, D.; Yu, P.; Li, Y.; Cai, M.; Zhao, Y. Polystyrene nanoplastic induces ROS production and affects the MAPK-HIF-1/NFkB-mediated antioxidant system in Daphnia pulex. Aquat. Toxicol. 2020, 220, 105420. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Chen, B.; Li, Q.; Liu, N.; Xia, B.; Zhu, L.; Qu, K. Toxicities of polystyrene nano- and microplastics toward marine bacterium Halomonas alkaliphila. Sci. Total Environ. 2018, 642, 1378–1385. [Google Scholar] [CrossRef] [PubMed]
- Sarasamma, S.; Audira, G. Nanoplastics Cause Neurobehavioral Impairments, Reproductive and Oxidative Damages, and Biomarker Responses in Zebrafish: Throwing up Alarms of Wide Spread Health Risk of Exposure. Int. J. Mol. Sci. 2020, 21, 1410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poma, A.; Vecchiotti, G.; Colafarina, S.; Zarivi, O.; Aloisi, M.; Arrizza, L.; Chichiriccò, G.; Di Carlo, P. In vitro genotoxicity of polystyrene nanoparticles on the human fibroblast Hs27 cell line. Nanomaterials 2019, 9, 1299. [Google Scholar] [CrossRef] [Green Version]
- Cortés, C.; Domenech, J.; Salazar, M.; Pastor, S.; Marcos, R.; Hernández, A. Nanoplastics as a potential environmental health factor: Effects of polystyrene nanoparticleson human intestinal epithelial Caco-2 cells. Environ. Sci. Nano 2020, 7, 272–285. [Google Scholar] [CrossRef]
- Schirinzi, G.F.; Perez-Pomeda, I.; Sanchis, J.; Rossini, C.; Farre, M.; Barcelo, D. Cytotoxic effects of commonly used nanomaterials and microplastics on cerebral and epithelial human cells. Environ. Res. 2017, 159, 579–587. [Google Scholar] [CrossRef]
- Rubio, L.; Barguill, I.; Domenech, J.; Marcos., R.; Hernández, A. Biological effects, including oxidative stress and genotoxic damage, of polystyrene nanoparticles in different human hematopoietic cell lines. J. Hazard. Mater. 2020, 398, 122900. [Google Scholar] [CrossRef]
- Li, Z.; Feng, C.; Wu, Y.; Guo, X. Impacts of nanoplastics on bivalve: Fluorescence tracing of organ accumulation, oxidative stress and damage. J. Hazard. Mater. 2020, 392, 122418. [Google Scholar] [CrossRef] [PubMed]
- Alomar, C.; Sureda, A.; Capo, X.; Guijarro, B.; Tejada, S.; Deudero, S. Microplastic ingestion by Mullussurmuletus Linnaeus, 1758 fish and its potential for causing oxidative stress. Environ. Res. 2017, 159, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Moor, A.C.E. Signaling pathways in cell death and survival after photodynamic therapy. J. Photochem. Photobiol. B 2000, 57, 1–13. [Google Scholar] [CrossRef]
- Zheng, T.; Yuan, D.; Liu, C. Molecular toxicity of nanoplastics involving in oxidative stress and desoxyribonucleic acid damage. J. Mol. Recognit. 2019, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Barboza, A.G.L.; Vieira, R.L.; Branco, V.; Figueiredo, N.; Carvalho, F.; Carvalho, C.; Guilhermino, L. Microplastics cause neurotoxicity, oxidative damage and energy-related changes and interact with the bioaccumulation of mercury in the European seabass, Dicentrarchus labrax (Linnaeus, 1758). Aquat. Toxicol. 2018, 195, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, F.; Garcia, R.A.; Pereira, P.B.; Fonseca, M.; Mestre, C.N.; Fonseca, G.T.; Ilharco, M.L.; Bebianno, J.M. Microplastics effects in Scrobicularia plana. Mar. Pollut. Bull. 2017, 122, 379–391. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Jin, R.S.; Chen, Z.Z.; Gao, Z.J.; Liu, N.Y.; Liu, H.J.; Feng, S.X. Single and combined effects of microplastics and cadmium on the cadmium accumulation, antioxidant defence and innate immunity of the discus fish (Symphysodon aequifasciatus). Environ. Pollut. 2018, 243, 462–471. [Google Scholar] [CrossRef] [PubMed]
- Paul-Pont, I.; Lacroix, C.; Fernandez, G.C.; Hegaret, H.; Lambert, C.; Le Goïc, N.; Frere, L.; Cassone, L.-A.; Sussarellu, R.; Fabioux, C.; et al. Exposure of marine mussels Mytilus spp. to polystyrene microplastics: Toxicity and influence on fluoranthene bioaccumulation. Environ Pollut. 2016, 216, 724–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowis, D.; Legat, M.; Grzela, T.; Niderla, J.; Wilczek, E.; Wilczynski, G.M.; Glodkowska, E.; Mrowka, P.; Issat, T.; Dulak, J.; et al. Heme oxygenase-1 protects tumor cells against photodynamic therapy-mediated cytotoxicity. Oncogene 2006, 25, 3365–3374. [Google Scholar] [CrossRef] [Green Version]
- Kulbacka, J.; Saczko, J.; Chwiłkowska, A. Stres oksydacyjny w procesach uszkodzenia komórek. Pol. Merk. Lek. 2009, 44, 1–4. [Google Scholar]
- Levine, L.R.; Stadtman, R.E. Oxidative modification of proteins during aging. Exp. Gerontol. 2001, 36, 1495–1502. [Google Scholar] [CrossRef]
- Zhang, W.; Xiao, S.; Ahn, U.D. Protein oxidation: Basic principles and implications for meat quality. Crit. Rev. Food Sci. Nat. 2013, 53, 1191–1201. [Google Scholar] [CrossRef]
- Shacter, E. Quantification and significance of protein oxidation in biological Samales. Drug Metab. Rev. 2000, 32, 307–326. [Google Scholar] [CrossRef]
- Hollóczki, O.; Gehrke, S. Nanoplastics can change the secondary structure of proteins. Sci. Rep. 2019, 9, 16013. [Google Scholar] [CrossRef] [Green Version]
- Gopinath, M.P.; Saranya, V.; Vijayakumar, S.; Meera, M.M.; Ruprekha, S.; Kunal, R.; Pranay, A.; Thomas, J.; Mukherjee, A.; Chandrasekaran, N. Assessment on interactive prospectives of nanoplastics with plasma proteins and the toxicological impacts of virgin, coronated and environmentally released-nanoplastics. Sci. Rep. 2019, 9, 8860. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Zhu, X.; Wu, D.; Song, E.; Song, Y. Compromised Autophagic Effect of Polystyrene Nanoplastics Mediated by protein corona was recovered after lysosomal degradation of corona. Environ. Sci. Technol. 2020, 54, 11485–11493. [Google Scholar] [CrossRef]
- Davies, M.J. Singlet oxygen-mediated damage to proteins and its consequences. Biochem. Biophys. Res. Commun. 2003, 305, 761–770. [Google Scholar] [CrossRef]
- Davies, M.J. The oxidative environment and protein damage. Biochim. Biophys. Acta 2005, 1703, 93–109. [Google Scholar] [CrossRef]
- Gracanin, M.; Hawkins, L.C.; Pattison, I.D.; Davies, J.M. Singlet-oxygen-mediated amino acid and protein oxidation: Formation of tryptophan peroxides and decomposition products. Free Radic. Biol. Med. 2009, 47, 92–102. [Google Scholar] [CrossRef] [PubMed]
- Thubagere, A.; Reinhard, B. Nanoparticle-induced apoptosis propagates through hydro-gen-peroxide-mediated bystander killing: Insights from a human intestinal epithelium in vitro model. ACS Nano 2010, 4, 3611–3622. [Google Scholar] [CrossRef] [PubMed]
- Wang., F.; Bexiga, M.; Anguissola, S.; Boya, P.; Simpson, J.C.; Salvati, A.; Dawson, K.A. Time resolved study of cell death mechanisms induced by amine-modified polystyrene nanoparticles. Nanoscale 2013, 5, 10868–10876. [Google Scholar] [CrossRef] [PubMed]
- Phuc, L.; Taniguchi, A. Polystyrene Nanoparticles Induce Apoptosis or Necrosis with or without Epidermal Growth Factor. J. Nanosci. Nanotechnol. 2019, 19, 4812–4817. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Wang, H.; He, C.; Jin, Y.; Fu, Z. Polystyrene nanoparticles trigger the activation of p38 MAPK and apoptosis via inducing oxidative stress in zebrafish and macrophage cells. Environ. Pollut. 2021, 269, 116075. [Google Scholar] [CrossRef]
- Xu, M.; Halimu, G.; Zhang, Q.; Song, Y.; Fu, X.; Li, Y.; Zhang, H. Internalization and toxicity: A preliminary study of effects of nanoplastic particles on human lung epithelial cell. Sci. Total Environ. 2019, 1, 133794. [Google Scholar] [CrossRef]
- Sicińska, P. Di-n-butyl phthalate, butylbenzyl phthalate, and their metabolites exhibit different apoptotic potential in human peripheral blood mononuclear cells. Food Chem. Toxicol. 2019, 133, 110750. [Google Scholar] [CrossRef]
- Sicińska, P.; Mokra, K.; Wozniak, K.; Michałowicz, J.; Bukowska, B. Genotoxic risk assessment and mechanism of DNA damage induced by phthalates and their metabolites in human peripheral blood mononuclear cells. Sci. Rep. 2021, 11, 1658. [Google Scholar] [CrossRef] [PubMed]
- Setsukinai, K.I.; Urano, Y.; Katsuko, K.; Majima, H.; Nagano, T. Development of novel fluorescence probes that can reliably detect reactive oxygen species and distinguish specific species. J. Biol. Chem. 2002, 278, 3170–3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steenbergen, R.; Drummen, G.; den Kamp, J.O.; Post, J. The use of cis-parinaric acid to measure lipid peroxidation in cardiomyocytes during ischemia reperfusion. Biochim. Biophys. Acta 1997, 1330, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Vivian, J.T.; Callis, P.R. Mechanisms of tryptophan fluorescence shifts in 858 proteins. Biophys. J. 2001, 80, 2093–2109. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, G.N.; Dedoussis, V.G.; Spanakos, G.; Gritzapis, D.A.; Baxevanis, N.C.; Papamichail, M. An improved fluoerescence assay for the determination of lymphocyte-mediated cytotoxicity using flow cytometry. J. Immunol. Methods 1994, 177, 101–111. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diameter in Dicated by Manufacturer | 29 nm | 44 nm | 72 nm |
|---|---|---|---|
| Mean Diameter ± SD | 27.96 ± 8.03 | 38.61 ± 8.61 | 68.45 ± 15.19 |
| Concentration (µg/mL) | Fluorescence of Cis-Parinaric Acid (% of Control) | Tryptofan Fluorescence (% of Control) | ||||
|---|---|---|---|---|---|---|
| 29 nm | 44 nm | 72 nm | 29 nm | 44 nm | 72 nm | |
| 0 | 100 | 100 | 100 | 100 | 100 | 100 |
| 0.001 | 96.83 ± 5.09 | 97.86 ± 5.86 | 98.30 ± 4.99 | 95.65 ± 5.28 | 96.28 ± 6.36 | 98.38 ± 7.34 |
| 0.01 | 94.80 ± 6.50 | 95.81 ± 6.27 | 95.13 ± 6.91 | 94.39 ± 6.71 | 95.97 ± 7.01 | 97.46 ± 5.71 |
| 0.1 | 83.30 ± 5.60 * | 94.00 ± 6.83 | 95.31 ± 6.99 | 86.19 ± 5.82 * | 94.66 ± 6.71 | 98.56 ± 7.51 |
| 1 | 79.04 ± 4.77 * | 91.03 ± 5.89 * | 89.46 ± 4.41 * | 82.88 ± 7.01 * | 89.29 ± 7.25 * | 95.06 ± 6.57 * |
| 10 | 76.21 ± 4.63 * | 83.32 ± 5.45 * | 85.56 ± 5.93 * | 83.00 ± 5.53 * | 87.92 ± 4.45 * | 83.75 ± 5.42 * |
| 100 | 74.23 ± 4.08 * | 77.29 ± 7.45 * | 74.22 ± 5.60 * | 83.31 ± 4.47 * | 84.88 ± 5.98 * | 78.86 ± 4.81 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kik, K.; Bukowska, B.; Krokosz, A.; Sicińska, P. Oxidative Properties of Polystyrene Nanoparticles with Different Diameters in Human Peripheral Blood Mononuclear Cells (In Vitro Study). Int. J. Mol. Sci. 2021, 22, 4406. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094406
Kik K, Bukowska B, Krokosz A, Sicińska P. Oxidative Properties of Polystyrene Nanoparticles with Different Diameters in Human Peripheral Blood Mononuclear Cells (In Vitro Study). International Journal of Molecular Sciences. 2021; 22(9):4406. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094406
Chicago/Turabian StyleKik, Kinga, Bożena Bukowska, Anita Krokosz, and Paulina Sicińska. 2021. "Oxidative Properties of Polystyrene Nanoparticles with Different Diameters in Human Peripheral Blood Mononuclear Cells (In Vitro Study)" International Journal of Molecular Sciences 22, no. 9: 4406. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094406