
The Effects of Cryogenic Storage on Human Dental Pulp Stem Cells

 ,
,  ,
,  ,
,  and
and
Abstract
:1. Introduction
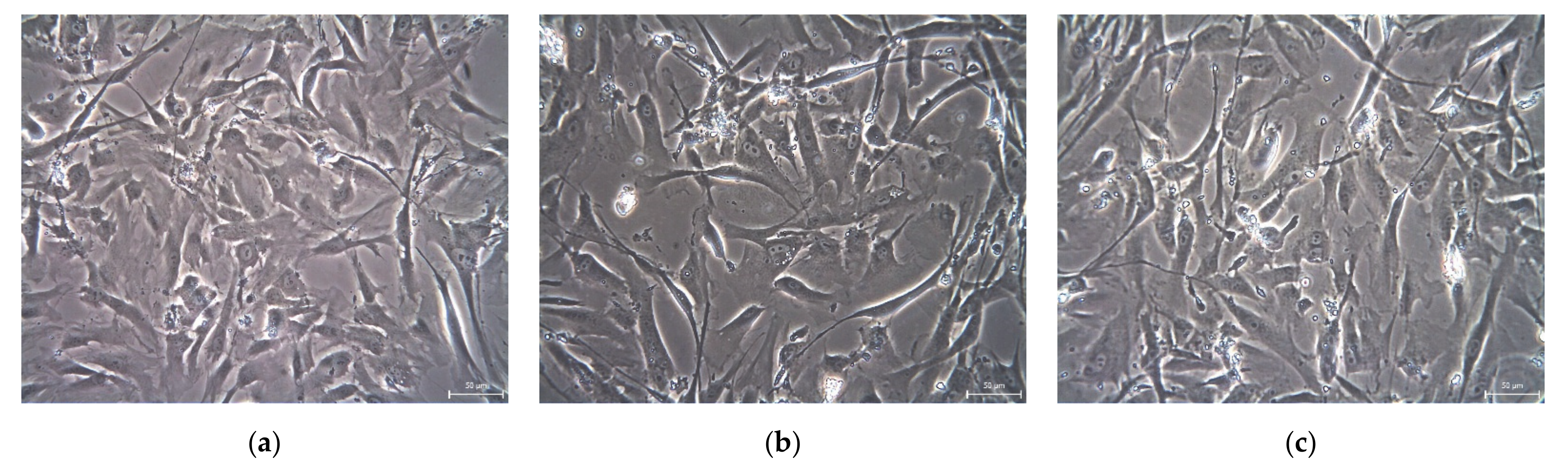
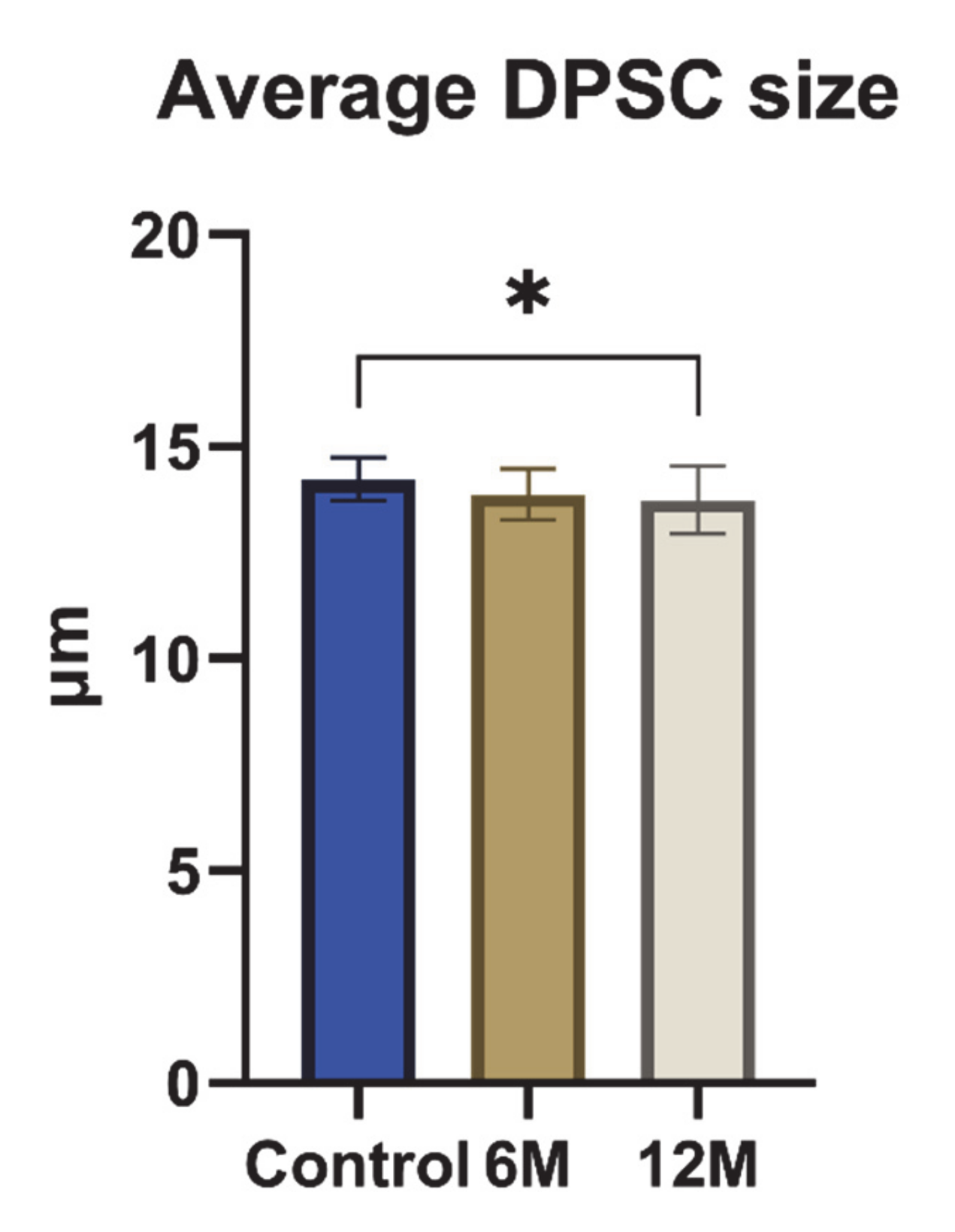
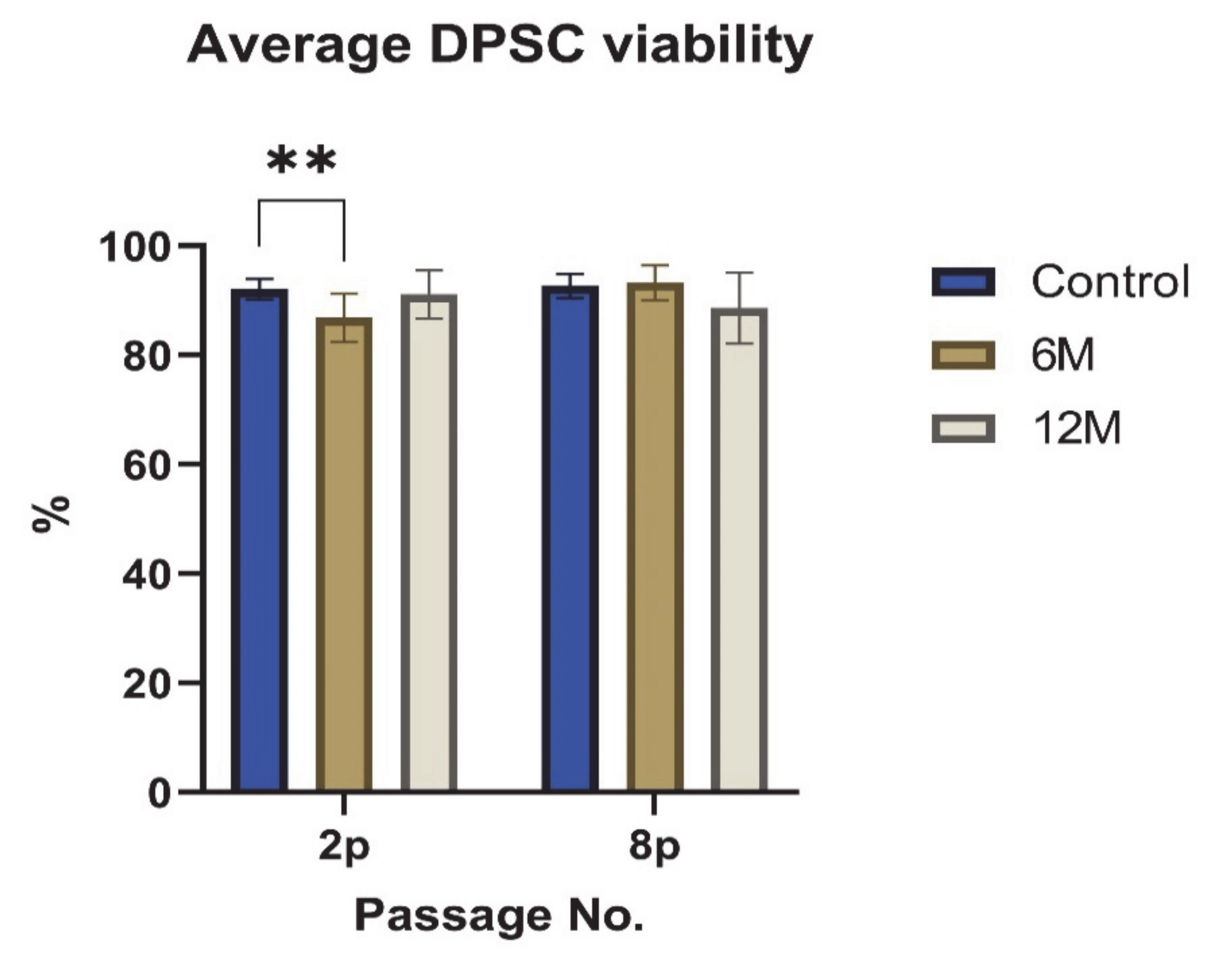
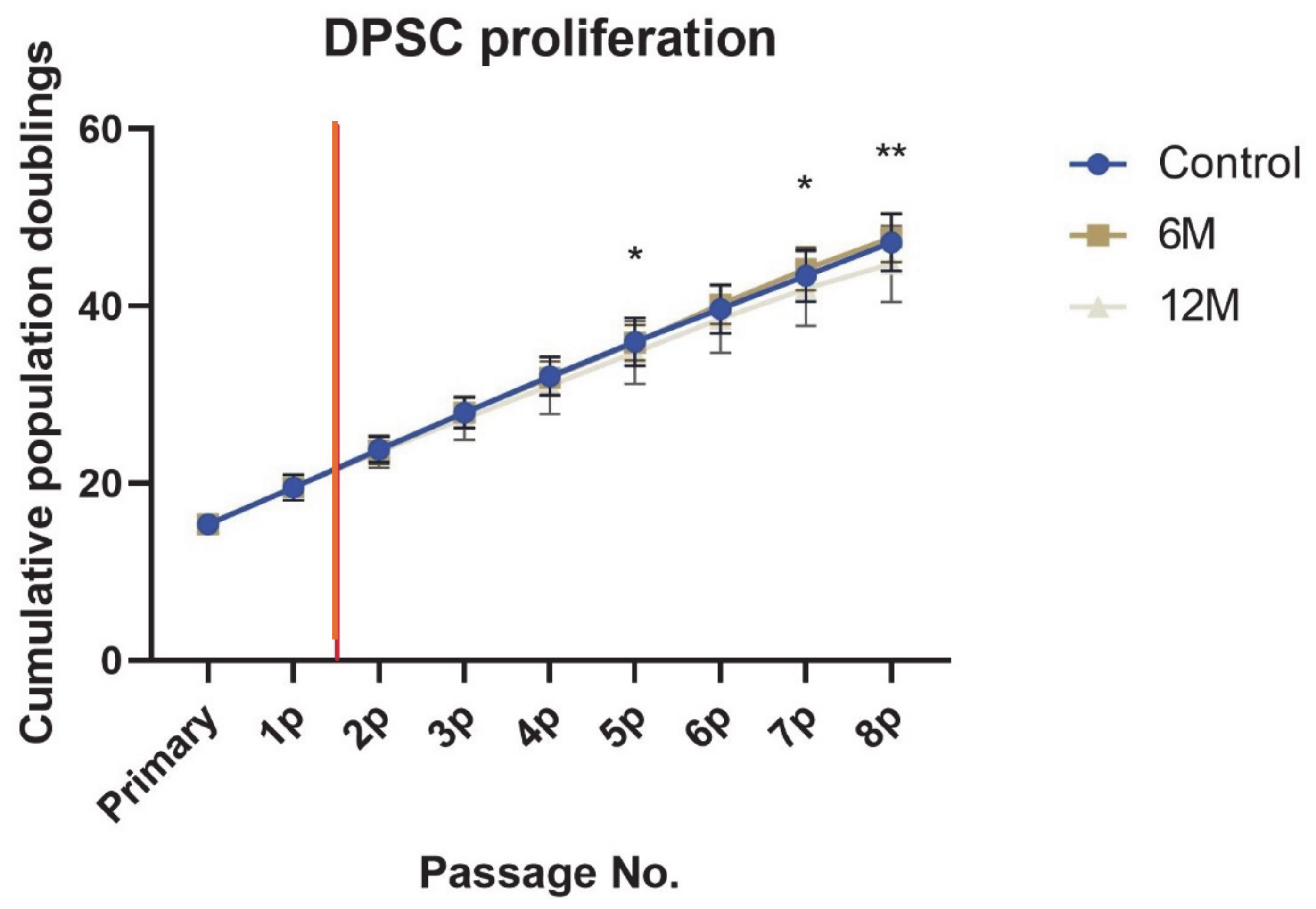
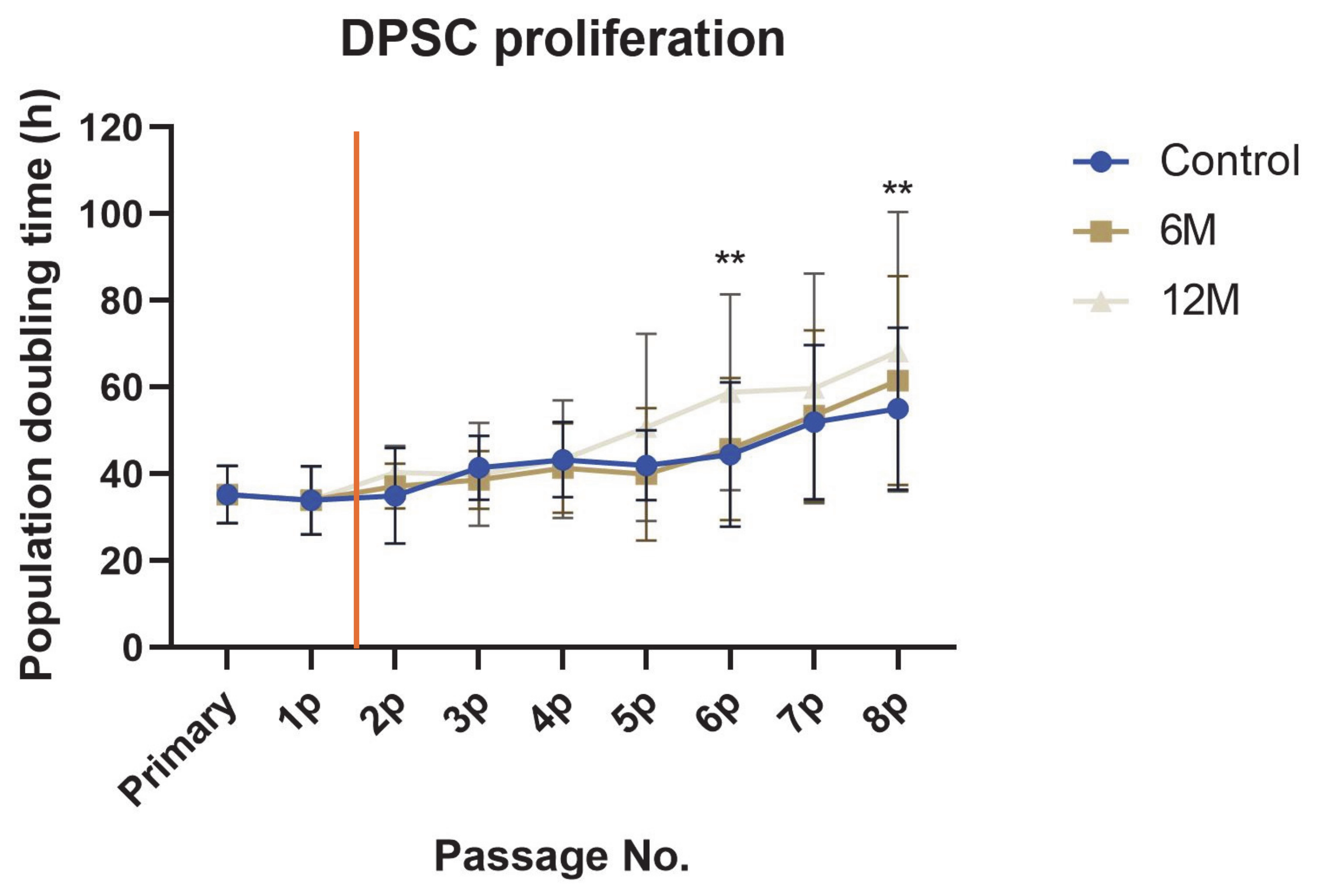
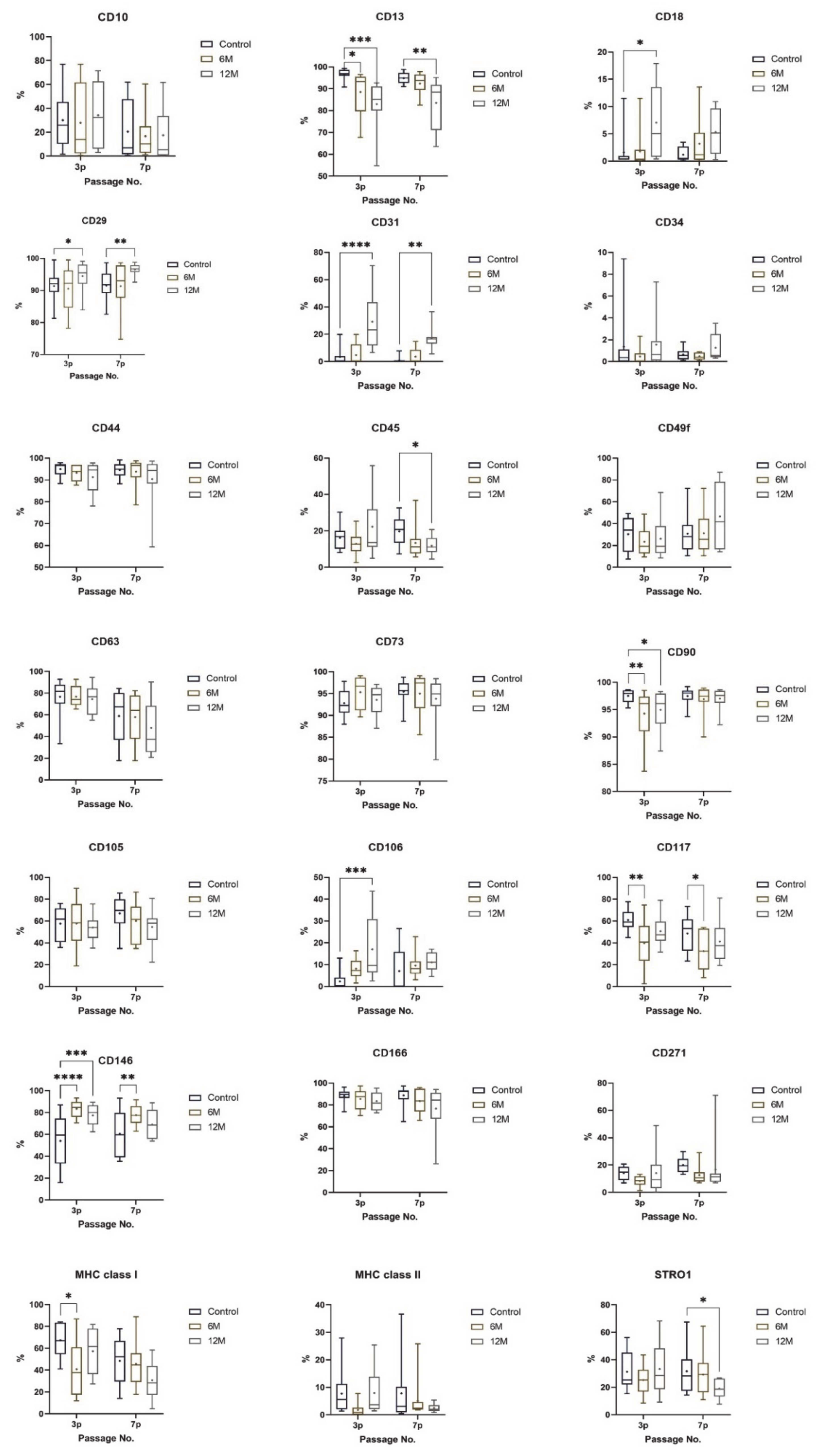
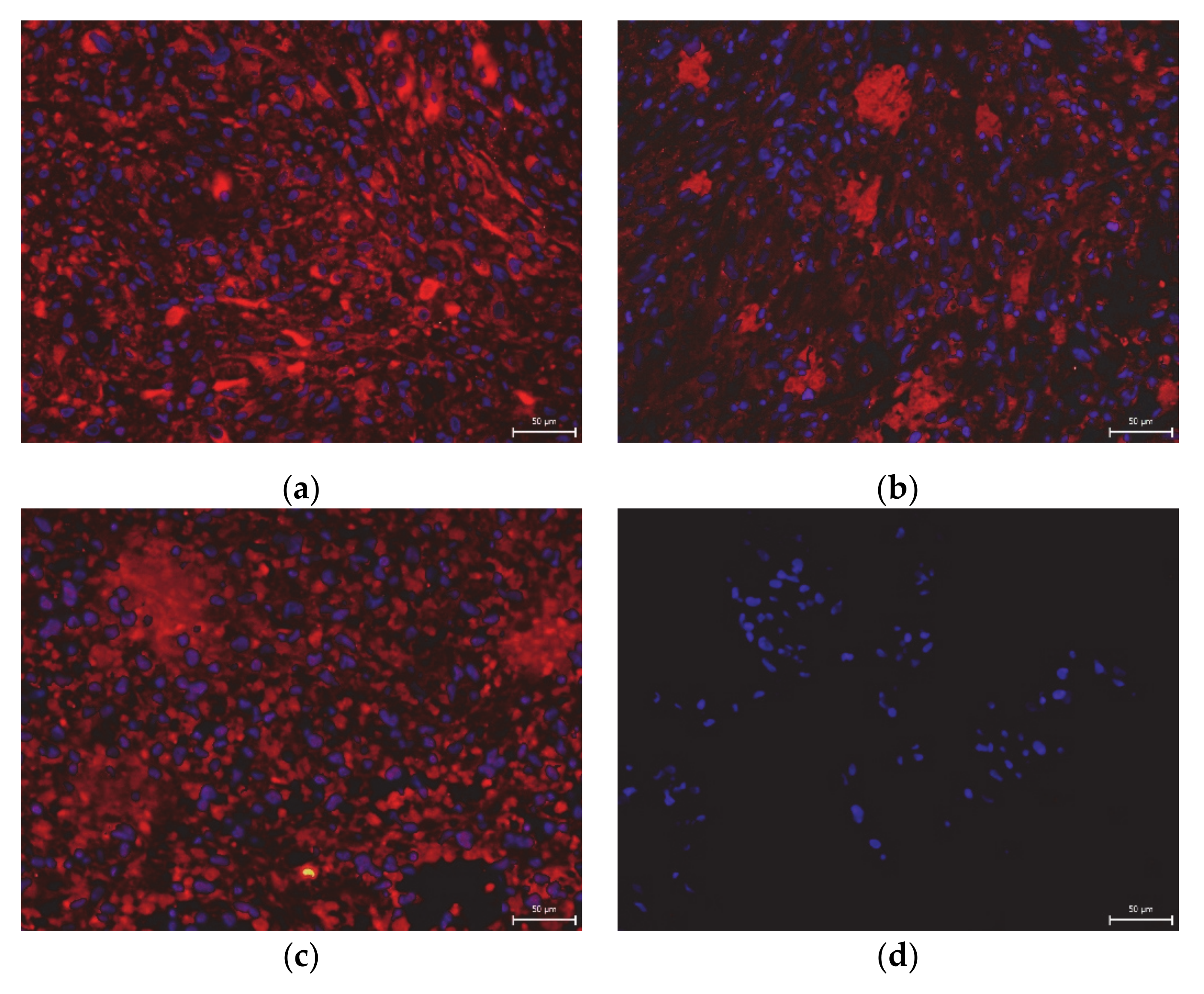
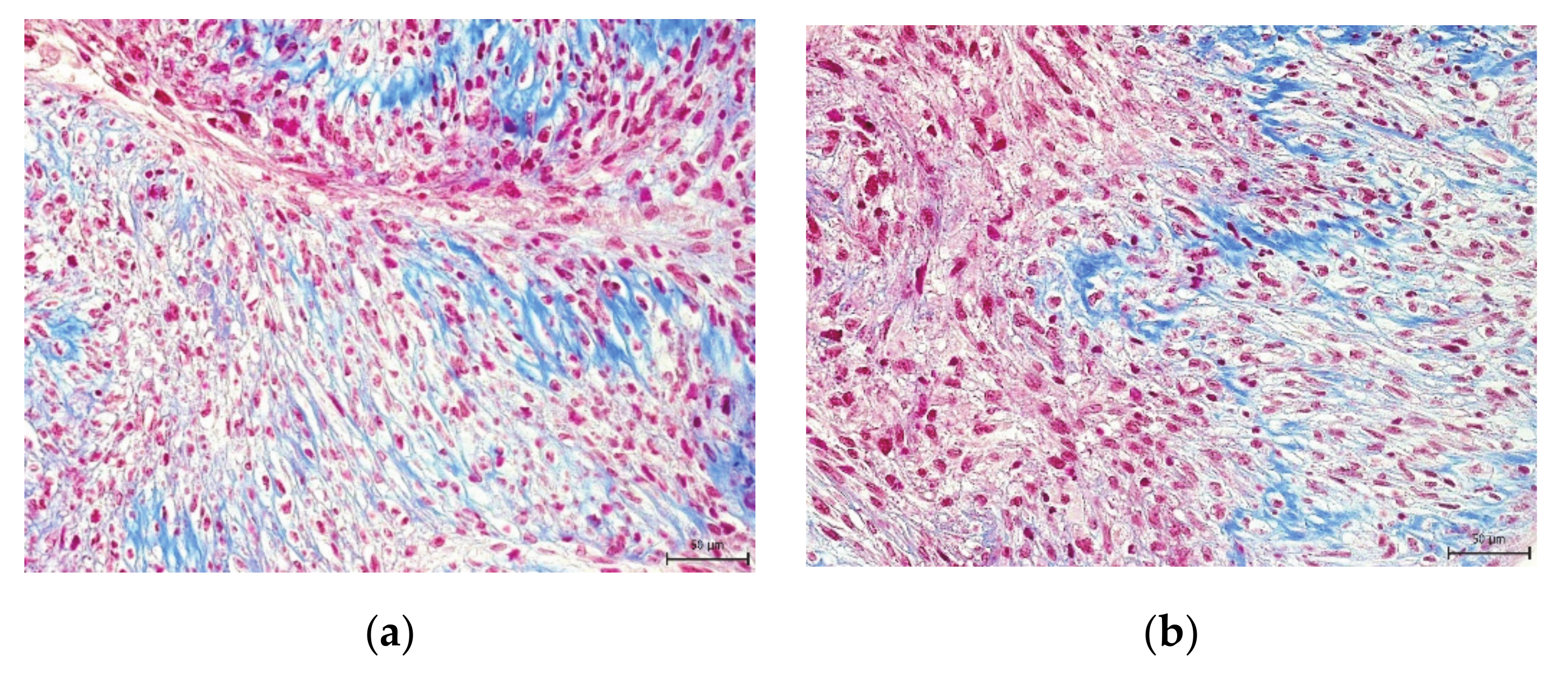
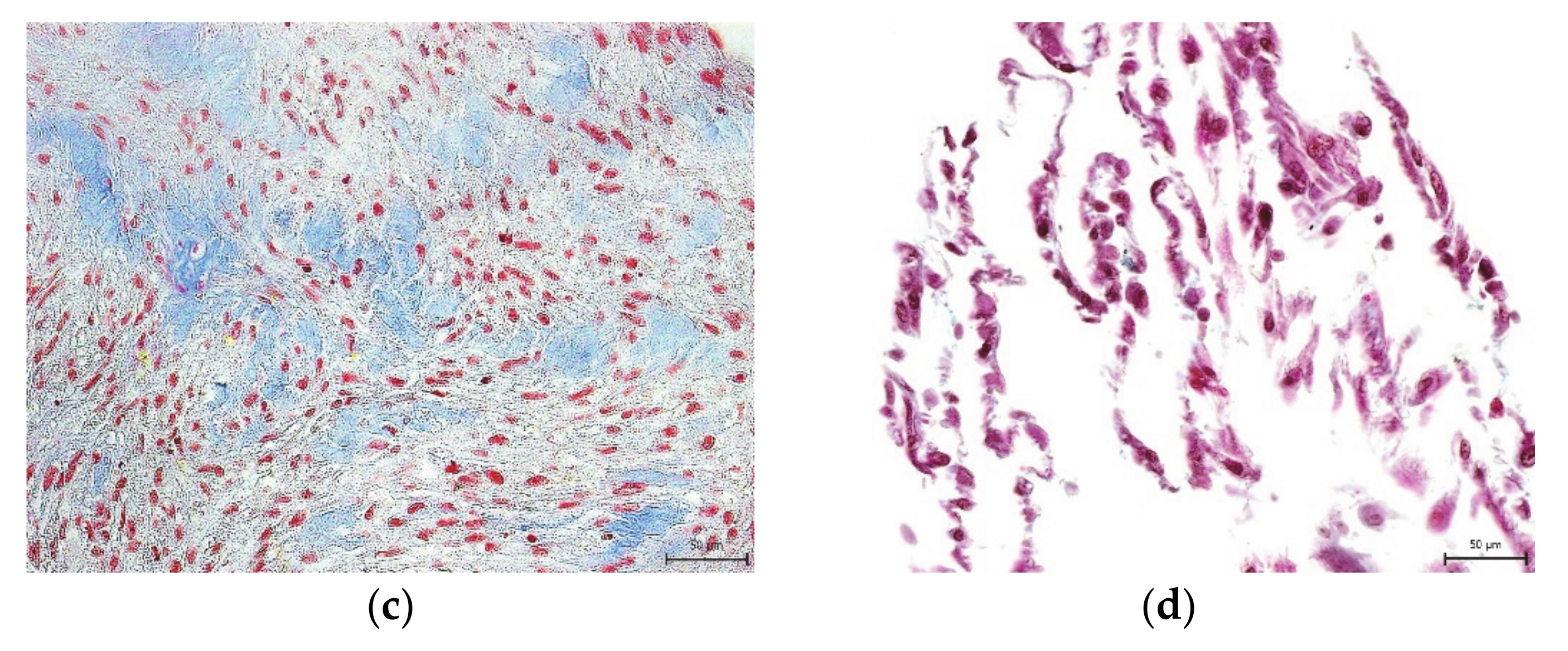
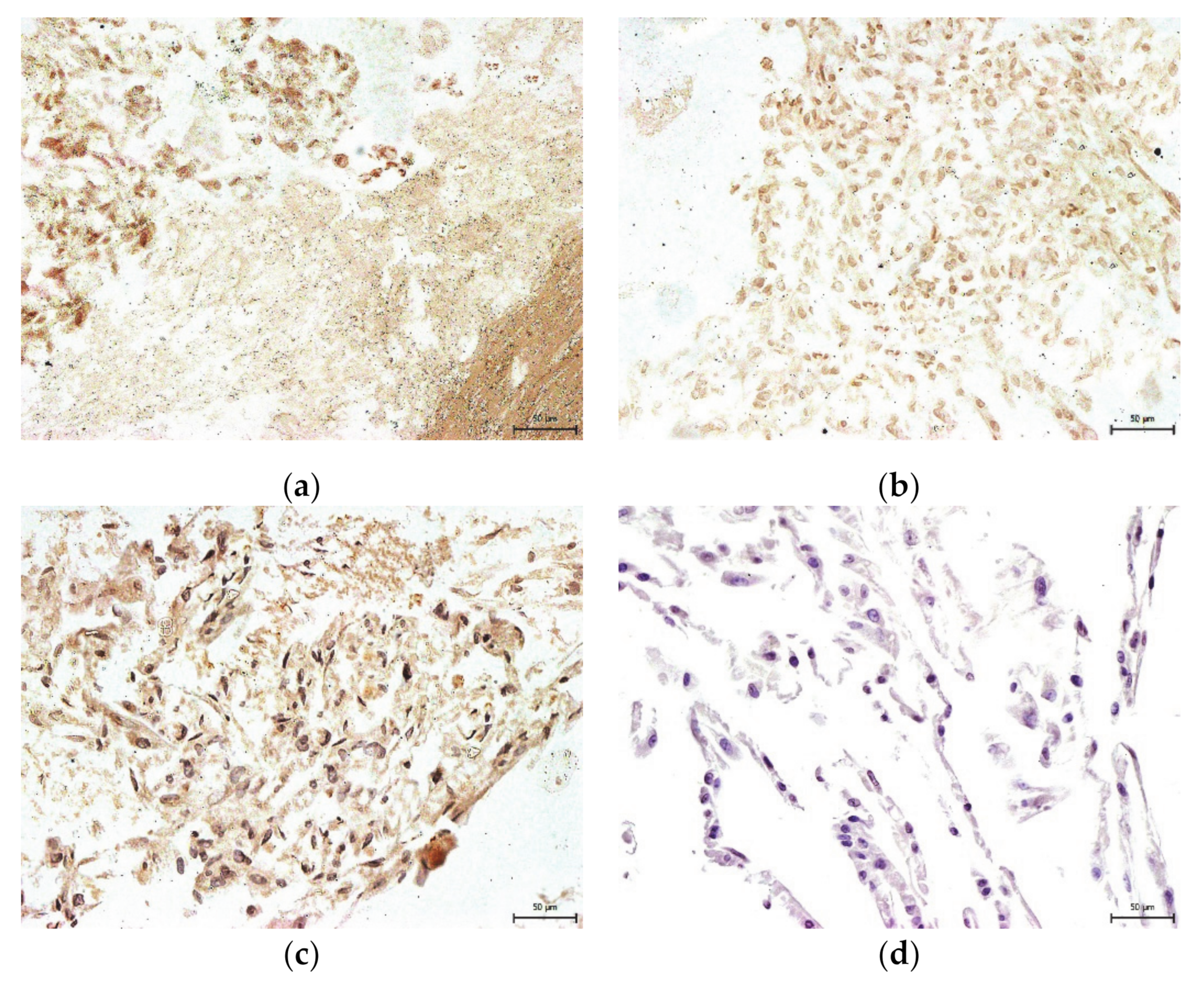
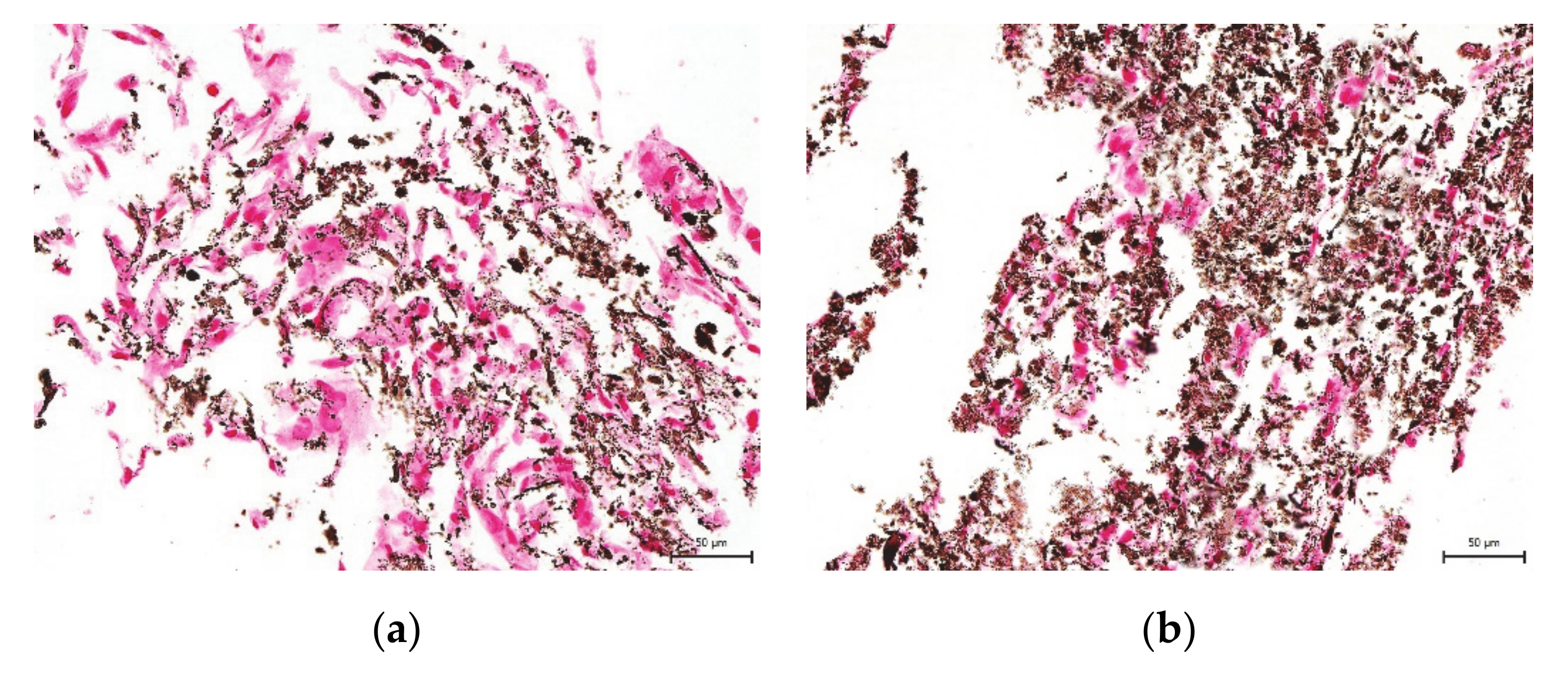
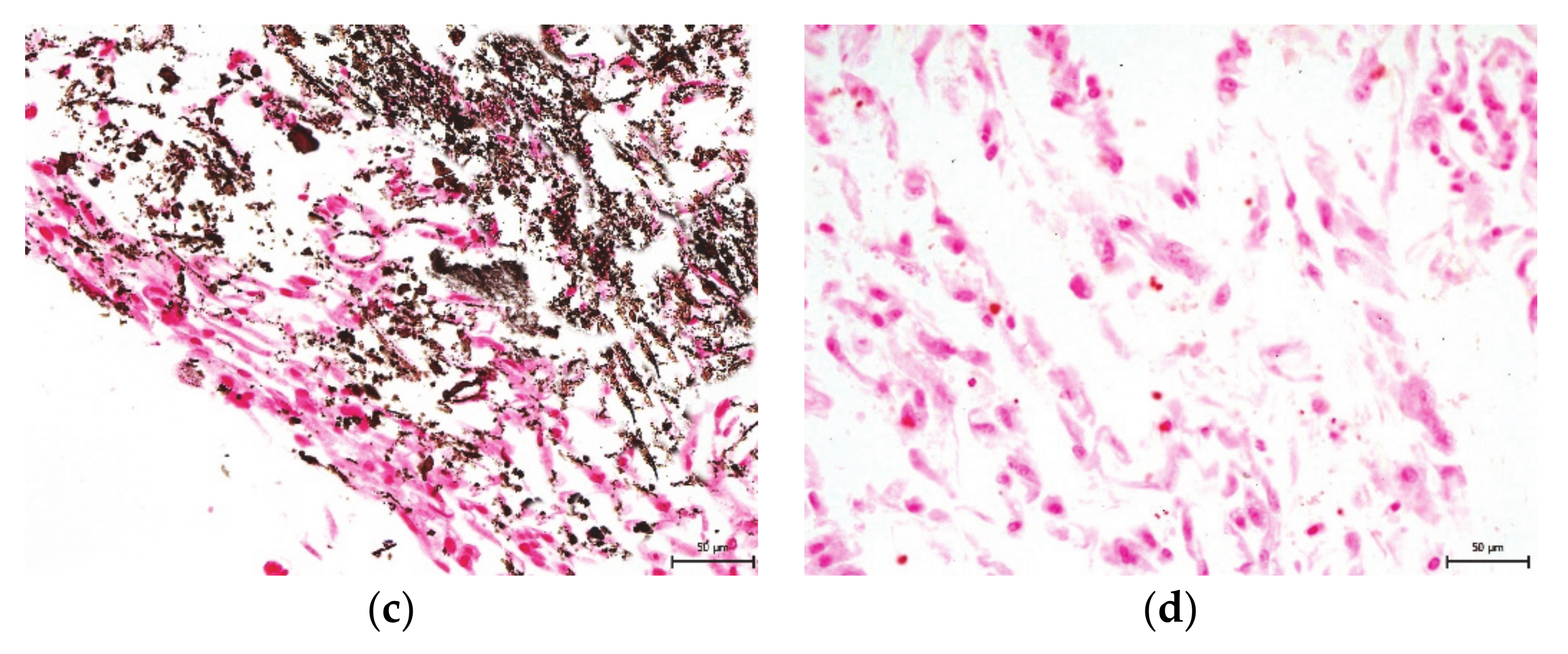
2. Results
3. Discussion
4. Materials and Methods
4.1. DPSC Lineages
4.2. The Uncontrolled-Rate Freezing
4.3. Cell Thawing
4.4. Cryopreservation Effect on DPSC Characteristics
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrúa, C.P.; Centeno, E.G.Z.; Rosa, L.C.D.; Amaral, C.C.D.; Severo, R.F.; Sarkis-Onofre, R.; Nascimento, G.G.; Cordenonzi, G.; Bast, R.K.; Demarco, F.F.; et al. How has dental pulp stem cells isolation been conducted? A scoping review. Braz. Oral Res. 2017, 31, e87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blondeau, F.; Daniel, N.G. Extraction of impacted mandibular third molars: Postoperative complications and their risk factors. J. Can. Dent. Assoc. 2007, 73, 325. [Google Scholar] [PubMed]
- Omar, Z.; Short, L.; Banting, D.W.; Saltaji, H. Profile changes following extraction orthodontic treatment: A comparison of first versus second premolar extraction. Int. Orthod. 2018, 16, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Shih, W.Y.; Hsieh, C.Y.; Tsai, T.P. Clinical evaluation of the timing of mesiodens removal. J. Chin. Med. Assoc. 2016, 79, 345–350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullen, S.F.; Critser, J.K. The science of cryobiology. Cancer Treat. Res. 2007, 138, 83–109. [Google Scholar] [CrossRef] [PubMed]
- Gao, D.; Critser, J.K. Mechanisms of cryoinjury in living cells. ILAR J. 2000, 41, 187–196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegg, D.E. Principles of cryopreservation. Methods Mol. Biol. 2015, 1257, 3–19. [Google Scholar] [CrossRef] [PubMed]
- Mazur, P. Kinetics of water loss from cells at subzero temperatures and the likelihood of intracellular freezing. J. Gen. Physiol. 1963, 47, 347–369. [Google Scholar] [CrossRef]
- Zambelli, A.; Poggi, G.; Da Prada, G.; Pedrazzoli, P.; Cuomo, A.; Miotti, D.; Perotti, C.; Preti, P.; Robustelli della Cuna, G. Clinical toxicity of cryopreserved circulating progenitor cells infusion. Anticancer Res. 1998, 18, 4705–4708. [Google Scholar]
- Stolzing, A.; Naaldijk, Y.; Fedorova, V.; Sethe, S. Hydroxyethylstarch in Cryopreservation—Mechanisms, Benefits and Problems. Transfus. Apher. Sci. 2012, 46, 137–147. [Google Scholar] [CrossRef]
- Pilbauerova, N.; Soukup, T.; Suchankova Kleplova, T.; Suchanek, J. Enzymatic Isolation, Amplification and Characterization of Dental Pulp Stem Cells. Folia Biol. 2019, 65, 124–133. [Google Scholar]
- Li, T.; Mai, Q.; Gao, J.; Zhou, C. Cryopreservation of human embryonic stem cells with a new bulk vitrification method. Biol. Reprod. 2010, 82, 848–853. [Google Scholar] [CrossRef] [Green Version]
- Bagchi, A.; Woods, E.J.; Critser, J.K. Cryopreservation and vitrification: Recent advances in fertility preservation technologies. Expert Rev. Med. Devices 2008, 5, 359–370. [Google Scholar] [CrossRef]
- Thirumala, S.; Zvonic, S.; Floyd, E.; Gimble, J.M.; Devireddy, R.V. Effect of various freezing parameters on the immediate post-thaw membrane integrity of adipose tissue derived adult stem cells. Biotechnol. Prog. 2005, 21, 1511–1524. [Google Scholar] [CrossRef]
- Kojima, S.; Kaku, M.; Kawata, T.; Sumi, H.; Shikata, H.; Abonti, T.R.; Kojima, S.; Fujita, T.; Motokawa, M.; Tanne, K. Cryopreservation of rat MSCs by use of a programmed freezer with magnetic field. Cryobiology 2013, 67, 258–263. [Google Scholar] [CrossRef]
- Almici, C.; Ferremi, P.; Lanfranchi, A.; Ferrari, E.; Verardi, R.; Marini, M.; Rossi, G. Uncontrolled-rate freezing of peripheral blood progenitor cells allows successful engraftment by sparing primitive and committed hematopoietic progenitors. Haematologica 2003, 88, 1390–1395. [Google Scholar]
- Detry, G.; Calvet, L.; Straetmans, N.; Cabrespine, A.; Ravoet, C.; Bay, J.O.; Petre, H.; Paillard, C.; Husson, B.; Merlin, E.; et al. Impact of uncontrolled freezing and long-term storage of peripheral blood stem cells at −80 °C on haematopoietic recovery after autologous transplantation. Report from two centres. Bone Marrow Transpl. 2014, 49, 780–785. [Google Scholar] [CrossRef] [Green Version]
- Setia, R.D.; Arora, S.; Handoo, A.; Choudhary, D.; Sharma, S.K.; Khandelwal, V.; Kapoor, M.; Bajaj, S.; Dadu, T.; Dhamija, G.; et al. Outcome of 51 autologous peripheral blood stem cell transplants after uncontrolled-rate freezing (“dump freezing”) using −80 °C mechanical freezer. Asian J. Transfus. Sci. 2018, 12, 117–122. [Google Scholar] [CrossRef]
- Zeng, G.; Hu, Y.; Hu, X.; Zeng, W.; Liang, X.; Liu, Y.; Peng, H.; Liao, Y.; Ren, Y.; Tang, Z.; et al. Cryopreservation of peripheral blood mononuclear cells using uncontrolled rate freezing. Cell Tissue Bank 2020, 21, 631–641. [Google Scholar] [CrossRef]
- Kumar, A.; Bhattacharyya, S.; Rattan, V. Effect of uncontrolled freezing on biological characteristics of human dental pulp stem cells. Cell Tissue Bank 2015, 16, 513–522. [Google Scholar] [CrossRef]
- Raik, S.; Kumar, A.; Rattan, V.; Seth, S.; Kaur, A.; Bhatta Charyya, S. Assessment of Post-thaw Quality of Dental Mesenchymal Stromal Cells After Long-Term Cryopreservation by Uncontrolled Freezing. Appl. Biochem. Biotechnol. 2020, 191, 728–743. [Google Scholar] [CrossRef]
- Woods, E.J.; Perry, B.C.; Hockema, J.J.; Larson, L.; Zhou, D.; Goebel, W.S. Optimized cryopreservation method for human dental pulp-derived stem cells and their tissues of origin for banking and clinical use. Cryobiology 2009, 59, 150–157. [Google Scholar] [CrossRef] [Green Version]
- Suchánek, J.; Visek, B.; Soukup, T.; El-Din Mohamed, S.K.; Ivancaková, R.; Mokrỳ, J.; Aboul-Ezz, E.H.; Omran, A. Stem cells from human exfoliated deciduous teeth--isolation, long term cultivation and phenotypical analysis. Acta Med. 2010, 53, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Li, L.; Xiao, L.; Zhang, D. Recycle the dental fairy’s package: Overview of dental pulp stem cells. Stem Cell. Res. Ther. 2018, 9, 347. [Google Scholar] [CrossRef]
- Suchanek, J.; Nasry, S.A.; Soukup, T. The Differentiation Potential of Human Natal Dental Pulp Stem Cells into Insulin-Producing Cells. Folia Biol. 2017, 63, 132–138. [Google Scholar]
- Nuti, N.; Corallo, C.; Chan, B.M.; Ferrari, M.; Gerami-Naini, B. Multipotent Differentiation of Human Dental Pulp Stem Cells: A Literature Review. Stem Cell Rev. Rep. 2016, 12, 511–523. [Google Scholar] [CrossRef]
- Smith, E.R.; Hadidian, Z.; Mason, M.M. The single--and repeated--dose toxicity of dimethyl sulfoxide. Ann. N. Y. Acad. Sci. 1967, 141, 96–109. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration (FDA). Guidance for Industry: Q3C Tables and List; Food and Drug Administration: Silver Spring, MD, USA, 2017.
- Suchánek, J.; Soukup, T.; Ivancaková, R.; Karbanová, J.; Hubková, V.; Pytlík, R.; Kucerová, L. Human dental pulp stem cells--isolation and long term cultivation. Acta Med. 2007, 50, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Laino, G.; d’Aquino, R.; Graziano, A.; Lanza, V.; Carinci, F.; Naro, F.; Pirozzi, G.; Papaccio, G. A new population of human adult dental pulp stem cells: A useful source of living autologous fibrous bone tissue (LAB). J. Bone Min. Res. 2005, 20, 1394–1402. [Google Scholar] [CrossRef] [PubMed]
- Mokry, J.; Soukup, T.; Micuda, S.; Karbanova, J.; Visek, B.; Brcakova, E.; Suchanek, J.; Bouchal, J.; Vokurkova, D.; Ivancakova, R. Telomere attrition occurs during ex vivo expansion of human dental pulp stem cells. J. Biomed. Biotechnol. 2010, 2010, 673513. [Google Scholar] [CrossRef]
- Pilbauerova, N.; Soukup, T.; Suchankova Kleplova, T.; Schmidt, J.; Suchanek, J. The Effect of Cultivation Passaging on the Relative Telomere Length and Proliferation Capacity of Dental Pulp Stem Cells. Biomolecules 2021, 11, 464. [Google Scholar] [CrossRef]
- Wang, X.; Hua, T.C.; Sun, D.W.; Liu, B.; Yang, G.; Cao, Y. Cryopreservation of tissue-engineered dermal replacement in Me2SO: Toxicity study and effects of concentration and cooling rates on cell viability. Cryobiology 2007, 55, 60–65. [Google Scholar] [CrossRef]
- Lin, C.S.; Xin, Z.C.; Dai, J.; Lue, T.F. Commonly used mesenchymal stem cell markers and tracking labels: Limitations and challenges. Histol. Histopathol. 2013, 28, 1109–1116. [Google Scholar] [CrossRef]
- Suchanek, J.; Kleplova, T.S.; Kapitan, M.; Soukup, T. The effect of fetal calf serum on human dental pulp stem cells. Acta Med. 2013; 56, 142–149. [Google Scholar] [CrossRef]
- Venkataramani, V.; Küffer, S.; Cheung, K.C.P.; Jiang, X.; Trümper, L.; Wulf, G.G.; Ströbel, P. CD31 Expression Determines Redox Status and Chemoresistance in Human Angiosarcomas. Clin. Cancer Res. 2018, 24, 460–473. [Google Scholar] [CrossRef] [Green Version]
- Vishwanath, V.R.; Nadig, R.R.; Nadig, R.; Prasanna, J.S.; Karthik, J.; Pai, V.S. Differentiation of isolated and characterized human dental pulp stem cells and stem cells from human exfoliated deciduous teeth: An in vitro study. J. Conserv. Dent. 2013, 16, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Mazzilli, F.; Rossi, T.; Sabatini, L.; Pulcinelli, F.M.; Rapone, S.; Dondero, F.; Gazzaniga, P.P. Human sperm cryopreservation and reactive oxygen species (ROS) production. Acta Eur. Fertil. 1995, 26, 145–148. [Google Scholar]
- Cook-Mills, J.M.; Marchese, M.E.; Abdala-Valencia, H. Vascular cell adhesion molecule-1 expression and signaling during disease: Regulation by reactive oxygen species and antioxidants. Antioxid. Redox Signal. 2011, 15, 1607–1638. [Google Scholar] [CrossRef] [Green Version]
- Gardin, C.; Ricci, S.; Ferroni, L. Dental Stem Cells (DSCs): Classification and Properties. In Dental Stem Cells: Regenerative Potential; Zavan, B., Bressan, E., Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–25. [Google Scholar] [CrossRef]
- Matsui, M.; Kobayashi, T.; Tsutsui, T.W. CD146 positive human dental pulp stem cells promote regeneration of dentin/pulp-like structures. Hum. Cell 2018, 31, 127–138. [Google Scholar] [CrossRef] [Green Version]
- Karbanová, J.; Soukup, T.; Suchánek, J.; Pytlík, R.; Corbeil, D.; Mokrý, J. Characterization of dental pulp stem cells from impacted third molars cultured in low serum-containing medium. Cells Tissues Organs 2011, 193, 344–365. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.T.; Rao, N.Q.; Fang, T.J.; Zhao, Y.M.; Ge, L.H. Comparison of the properties of CD146 positive and CD146 negative subpopulations of stem cells from human exfoliated deciduous teeth. Beijing Da Xue Xue Bao Yi Xue Ban 2018, 50, 284–292. [Google Scholar] [PubMed]
- Rusu, M.C.; Loreto, C.; Sava, A.; Manoiu, V.; Didilescu, A.C. Human adult dental pulp CD117/c-kit-positive networks of stromal cells. Folia Morphol. 2014, 73, 68–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blume-Jensen, P.; Claesson-Welsh, L.; Siegbahn, A.; Zsebo, K.M.; Westermark, B.; Heldin, C.H. Activation of the human c-kit product by ligand-induced dimerization mediates circular actin reorganization and chemotaxis. EMBO J. 1991, 10, 4121–4128. [Google Scholar] [CrossRef] [PubMed]
- LeBien, T.W.; McCormack, R.T. The common acute lymphoblastic leukemia antigen (CD10)--emancipation from a functional enigma. Blood 1989, 73, 625–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongeneel, C.V.; Quackenbush, E.J.; Ronco, P.; Verroust, P.; Carrel, S.; Letarte, M. Common acute lymphoblastic leukemia antigen expressed on leukemia and melanoma cell lines has neutral endopeptidase activity. J. Clin. Invest. 1989, 83, 713–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledesma-Martinez, E.; Mendoza-Nunez, V.M.; Santiago-Osorio, E. Mesenchymal Stem Cells Derived from Dental Pulp: A Review. Stem Cells Int. 2016, 2016, 4709572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, N.K.; Kelleher, D. Not Just an Adhesion Molecule: LFA-1 Contact Tunes the T Lymphocyte Program. J. Immunol. 2017, 199, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Todd, R.F., 3rd. The continuing saga of complement receptor type 3 (CR3). J. Clin. Invest. 1996, 98, 1–2. [Google Scholar] [CrossRef]
- Gomez, J.C.; Doerschuk, C.M. The role of CD18 in the production and release of neutrophils from the bone marrow. Lab. Invest. 2010, 90, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Lozano, F.J.; Bueno, C.; Insausti, C.L.; Meseguer, L.; Ramirez, M.C.; Blanquer, M.; Marin, N.; Martinez, S.; Moraleda, J.M. Mesenchymal stem cells derived from dental tissues. Int. Endod. J. 2011, 44, 800–806. [Google Scholar] [CrossRef] [Green Version]
- Lertkiatmongkol, P.; Liao, D.; Mei, H.; Hu, Y.; Newman, P.J. Endothelial functions of platelet/endothelial cell adhesion molecule-1 (CD31). Curr. Opin. Hematol. 2016, 23, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Sidney, L.E.; Branch, M.J.; Dunphy, S.E.; Dua, H.S.; Hopkinson, A. Concise review: Evidence for CD34 as a common marker for diverse progenitors. Stem Cells 2014, 32, 1380–1389. [Google Scholar] [CrossRef] [Green Version]
- Morath, I.; Hartmann, T.N.; Orian-Rousseau, V. CD44: More than a mere stem cell marker. Int. J. Biochem. Cell Biol. 2016, 81, 166–173. [Google Scholar] [CrossRef]
- Rheinländer, A.; Schraven, B.; Bommhardt, U. CD45 in human physiology and clinical medicine. Immunol. Lett. 2018, 196, 22–32. [Google Scholar] [CrossRef]
- Sundberg, M.; Jansson, L.; Ketolainen, J.; Pihlajamäki, H.; Suuronen, R.; Skottman, H.; Inzunza, J.; Hovatta, O.; Narkilahti, S. CD marker expression profiles of human embryonic stem cells and their neural derivatives, determined using flow-cytometric analysis, reveal a novel CD marker for exclusion of pluripotent stem cells. Stem Cell Res. 2009, 2, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Weng, J.; Krementsov, D.N.; Khurana, S.; Roy, N.H.; Thali, M. Formation of syncytia is repressed by tetraspanins in human immunodeficiency virus type 1-producing cells. J. Virol. 2009, 83, 7467–7474. [Google Scholar] [CrossRef] [Green Version]
- Kweekel, D.M.; Antonini, N.F.; Nortier, J.W.; Punt, C.J.; Gelderblom, H.; Guchelaar, H.J. Explorative study to identify novel candidate genes related to oxaliplatin efficacy and toxicity using a DNA repair array. Br. J. Cancer 2009, 101, 357–362. [Google Scholar] [CrossRef]
- Craig, W.; Kay, R.; Cutler, R.L.; Lansdorp, P.M. Expression of Thy-1 on human hematopoietic progenitor cells. J. Exp. Med. 1993, 177, 1331–1342. [Google Scholar] [CrossRef] [Green Version]
- Saalbach, A.; Anderegg, U. Thy-1: More than a marker for mesenchymal stromal cells. FASEB J. 2019, 33, 6689–6696. [Google Scholar] [CrossRef]
- Dallas, N.A.; Samuel, S.; Xia, L.; Fan, F.; Gray, M.J.; Lim, S.J.; Ellis, L.M. Endoglin (CD105): A marker of tumor vasculature and potential target for therapy. Clin. Cancer Res. 2008, 14, 1931–1937. [Google Scholar] [CrossRef] [Green Version]
- Fonsatti, E.; Maio, M. Highlights on endoglin (CD105): From basic findings towards clinical applications in human cancer. J. Transl. Med. 2004, 2, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, R.; Gegg, M.; Longbottom, R.; Adamson, P.; Turowski, P.; Greenwood, J. ICAM-1-mediated endothelial nitric oxide synthase activation via calcium and AMP-activated protein kinase is required for transendothelial lymphocyte migration. Mol. Biol. Cell 2009, 20, 995–1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cook-Mills, J.M. VCAM-1 signals during lymphocyte migration: Role of reactive oxygen species. Mol. Immunol. 2002, 39, 499–508. [Google Scholar] [CrossRef] [Green Version]
- Covas, D.T.; Panepucci, R.A.; Fontes, A.M.; Silva, W.A., Jr.; Orellana, M.D.; Freitas, M.C.; Neder, L.; Santos, A.R.; Peres, L.C.; Jamur, M.C.; et al. Multipotent mesenchymal stromal cells obtained from diverse human tissues share functional properties and gene-expression profile with CD146+ perivascular cells and fibroblasts. Exp. Hematol. 2008, 36, 642–654. [Google Scholar] [CrossRef]
- Russell, K.C.; Phinney, D.G.; Lacey, M.R.; Barrilleaux, B.L.; Meyertholen, K.E.; O’Connor, K.C. In vitro high-capacity assay to quantify the clonal heterogeneity in trilineage potential of mesenchymal stem cells reveals a complex hierarchy of lineage commitment. Stem Cells 2010, 28, 788–798. [Google Scholar] [CrossRef]
- Swart, G.W. Activated leukocyte cell adhesion molecule (CD166/ALCAM): Developmental and mechanistic aspects of cell clustering and cell migration. Eur. J. Cell Biol. 2002, 81, 313–321. [Google Scholar] [CrossRef]
- Ponnaiyan, D.; Bhat, K.M.; Bhat, G.S. Comparison of immuno-phenotypes of stem cells from human dental pulp and periodontal ligament. Int. J. Immunopathol. Pharm. 2012, 25, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Quirici, N.; Soligo, D.; Bossolasco, P.; Servida, F.; Lumini, C.; Deliliers, G.L. Isolation of bone marrow mesenchymal stem cells by anti-nerve growth factor receptor antibodies. Exp. Hematol. 2002, 30, 783–791. [Google Scholar] [CrossRef]
- Hewitt, E.W. The MHC class I antigen presentation pathway: Strategies for viral immune evasion. Immunology 2003, 110, 163–169. [Google Scholar] [CrossRef]
- Machado Cde, V.; Telles, P.D.; Nascimento, I.L. Immunological characteristics of mesenchymal stem cells. Rev. Bras. Hematol. Hemoter. 2013, 35, 62–67. [Google Scholar] [CrossRef]
- Lv, F.J.; Tuan, R.S.; Cheung, K.M.; Leung, V.Y. Concise review: The surface markers and identity of human mesenchymal stem cells. Stem Cells 2014, 32, 1408–1419. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
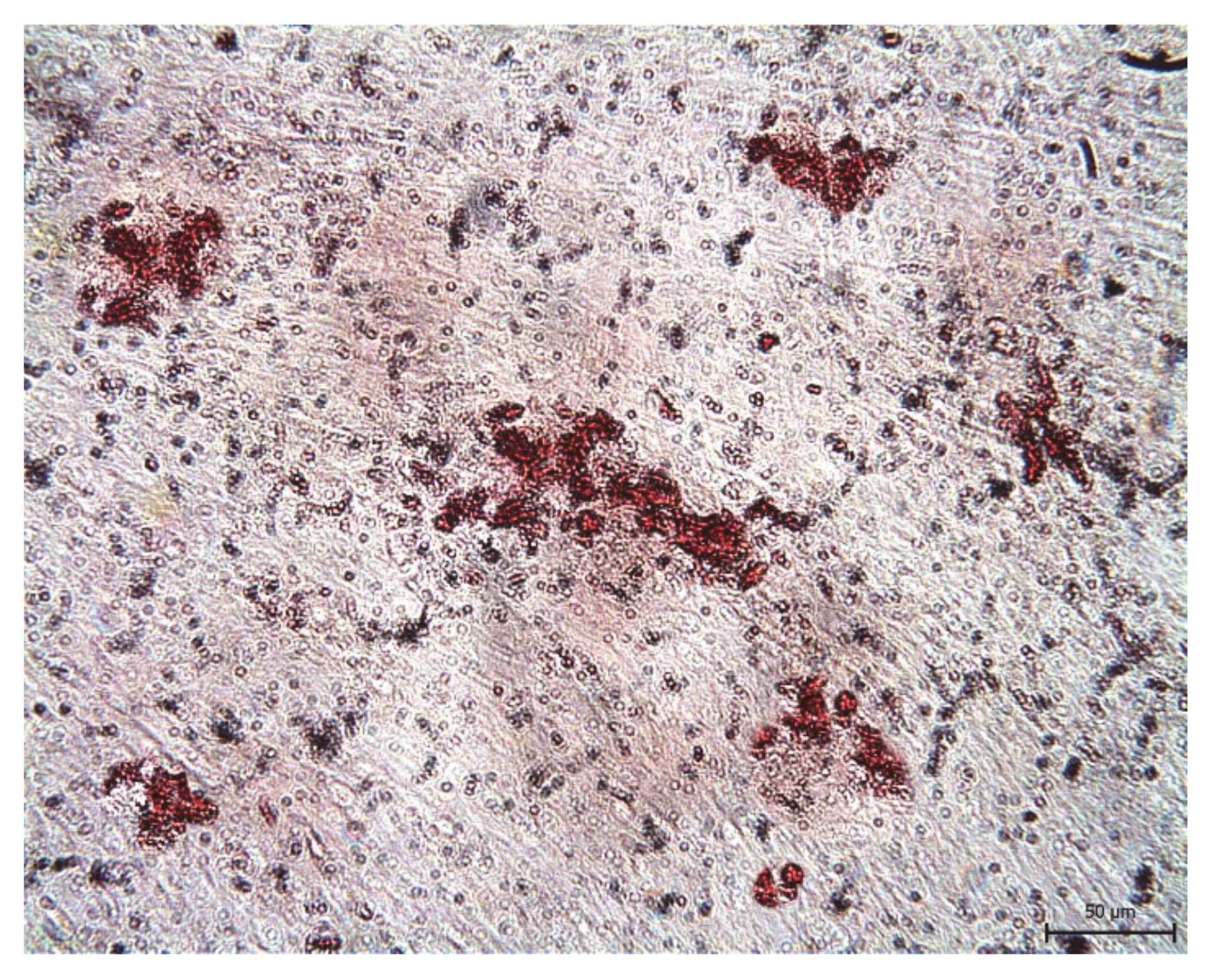{kind=link}
| Lineage | Donors | Age | Tooth | Root Development | Eruption |
|---|---|---|---|---|---|
| Z01 | Female | 14 | Mesiodens | Up to 1/2 | Impacted |
| Z02 | Female | 16 | M2 | More than 1/2 | Impacted |
| Z03 | Female | 16 | M3 | Up to 1/2 | Impacted |
| Z04 | Female | 14 | P1 | Fully developed | Fully erupted |
| Z05 | Female | 15 | M3 | Up to 1/2 | Impacted |
| Z06 | Female | 15 | M3 | Up to 1/2 | Impacted |
| Z07 | Female | 18 | M3 | More than 1/2 | Semi-impacted |
| Z08 | Male | 13 | P1 | More than 1/2 | Impacted |
| Z09 | Male | 13 | P1 | More than 1/2 | Impacted |
| Z10 | Female | 16 | Supernumerary P | More than 1/2 | Impacted |
| CD Marker | Name | Cellular Expression | Function | DPSC Expression | References |
|---|---|---|---|---|---|
| CD10 | Neprilysin: membrane metalloendopeptidase (MME), neutral endopeptidase (NEP) | CD10 is expressed on the B and T cell precursors, bone marrow stromal cells, lymphoblastic cells, Burkitt’s, and follicular germinal center lymphomas, immature B cells within the adult bone marrow, and on cells from patients with chronic myelocytic leukemia (CML). CD10 is also present on myoepithelial breast cells, bile canaliculi, fibroblasts, with incredibly high expression on the brush border of kidney and gut epithelial cells.Thought to be expressed during the first stages of heavy chain gene rearrangement and in an immunological context | It regulates B cell growth; it reduces the cellular response to peptide hormone in an immunological context.It is thought that the enzyme modulates the enkephalin-mediated inflammatory responseinvolved in the regulation of chemotactic and inflammatory processes involving neutrophils | Positive | [41,47,48] |
| CD13 | Alanyl aminopeptidase (AAP) or aminopeptidase N (AP-N) | CD13 is expressed on the surface of early committed progenitors of granulocytes and monocytes CFU-GM, and all these lineages’ cells as they mature. It is also expressed on endothelial cells, epithelial cells from proximal renal tubules and intestinal brush border, bone marrow, stromal cells, fibroblasts, brain cells, osteoclasts, and cells lining bile duct canaliculi, and is expressed on a small proportion of large granular lymphocytes but not on other lymphocytes | It has a role in cell surface antigen presentation by trimming the N-terminal aa from MHC Class II-bound peptides. CD13 ectopeptidase activity is also thought to downregulate cellular responses to peptide hormones by reducing the local concentration of peptide available for receptor binding | Positive | [41,49] |
| CD18 | Integrin beta chain-2: adhesive and signaling molecule for hematopoietic cell line | Lymphocyte Functions Associated Antigen 1 (binding CD18 and CD11) is a protein found on B cells, T cells, macrophages, neutrophils, and Natural Killer cells. The binding of CD18 and CD11b-d results in complement receptors’ formation, which are proteins mainly found on neutrophils, macrophages, and NK cells | The complement receptors participate in the innate immune response by recognizing foreign antigen peptides and phagocytizing them, thus destroying the antigen. The CD18 plays a role in the production and release of neutrophils from bone marrow | Negative | [50,51,52] |
| CD29 | Integrin beta-1 | Various cell types, including mesenchymal stem cells | Integrin family members are membrane receptors involved in cell adhesion and recognition in various processes, including embryogenesis, hemostasis, tissue repair, immune response, and metastatic diffusion of tumor cells | Positive | [41,53] |
| CD31 | Platelet endothelial cell adhesion molecule (PECAM-1) | Human granulocytes, monocytes, and platelets | It plays a role in endothelial cell intercellular junctions. The encoded protein is a member of the immunoglobulin superfamily, and is likely involved in leukocyte transmigration, angiogenesis, and integrin activation | Negative | [54] |
| CD34 | A transmembrane phosphoglycoprotein | Hematopoietic stem cells, hematopoietic progenitor cells, vascular endothelial progenitors, embryonic fibroblasts | CD34 plays a role in cell adhesion and the regulation of cell differentiation and proliferation | Negative | [55] |
| CD44 | A cell-surface glycoprotein, Hermes antigen, Pgp-1 | Leukocytes, erythrocytes; hematopoietic and non-hematopoietic cells (no platelets) | It binds hyaluronic acid, mediates adhesion of leukocytes, adhesion of leukocytes, endothelial cells, stromal cells, and ECM | Positive | [41,56] |
| CD45 | Protein tyrosine phosphatase, receptor type C | It is present in various isoforms on all differentiated hematopoietic cells (except erythrocytes and plasma cells). | CD45 is an essential regulator of T- and B-cell antigen receptor signaling | Negative | [41,57] |
| CD49f | Alpha-6 Integrin | Memory T cells, thymocytes, monocytes, memory B cells, platelets, megakaryocytes, epithelial cells, endothelial cells, cytotrophoblasts | Alpha-6 Integrin, associates with CD29, binds laminin, adhesion and cell migration, embryogenesis | Positive | [58] |
| CD63 | Granulophysin, LAMP-3 (Lysosomal-associated membrane 3), ME491, tetraspanin membrane protein | Several normal tissues, melanoma cells, a lysosomal membrane glycoprotein located mainly in the cytoplasm | It regulates melanoma cell motility and their adhesion and migration on substrates associated with beta1 integrins. It is associated with cell development and activation, and it may also function as a blood platelet activation marker | Positive | [59] |
| CD73 | 5’-nucleotidase, NTE, NT5 | It is a surface enzyme expressed on multiple cells and used as a marker of lymphocyte differentiation | Enzyme catalyzes the conversion at neutral pH of purine 5-prime mononucleotides to nucleosides, the preferred substrate being AMP. It may also have a role in cell adhesion | Positive | [60] |
| CD90 | Thy-1 | It is expressed by hematopoietic stem cells, and neurons in all species studied, and it is highly expressed in connective tissue and various fibroblast and stromal cell lines. It is also expressed on all thymocytes and peripheral T cells in mice, but, in humans, it is expressed only on a small % of fetal thymocytes, 10–40% of CD34+ cells in bone marrow, and <1% of CD3+CD4+ lymphocytes in the peripheral circulation. It is also expressed by human lymph node HEV endothelium but not other endothelia | Thy-1 can be used as a marker for a variety of stem cells and the axonal processes of mature neurons | Positive | [41,61,62] |
| CD105 | EndoglinSH2 | Endothelial cell, mesenchymal stem cell, erythroid precursors, activated monocytes, macrophages, and human vascular endothelial cells. Expression of endoglin is elevated on the endothelial cells of healing wounds, developing embryos, inflammatory tissues, and solid tumors.Endoglin is a marker of activated endothelium, and its vascular expression is limited to proliferating cells. | The possible ligand for an integrin, angiogenesis, modulates response to TGF beta 1postulated that endoglin is involved in the cytoskeletal organization affecting cell morphology and migrationdevelopment of the cardiovascular system, and in vascular remodelingprotective role of CD105 against pro-apoptotic factors | Positive | [41,63,64] |
| CD106 | Vascular cell adhesion protein 1 (VCAM-1) | Activated endothelial cellsmyeloid lineage and bone marrow stromal cellsfollicular dendritic cells, interdigitating reticulum cells, and Kupffer cells | It is an adhesion molecule, ligand for VLA-4, leukocyte adhesion, transmigration, and T cells’ co-stimulation.It encodes a cell surface sialoglycoprotein expressed by cytokine-activated endothelium. This type I membrane protein mediates leukocyte-endothelial cell adhesion and signal transduction, and may play a role in developing atherosclerosis and rheumatoid arthritis.CD106 expressed in non-vascular tissues has been implicated in the interaction of hematopoietic progenitors with bone marrow stromal cells, B cell binding to follicular dendritic cells, co-stimulation of T cells, and embryonic development | Positive | [41,65,66] |
| CD117 | Tyrosine-protein kinase KIT, mast or stem cell growth factor receptor (SCF) | KIT is a cytokine receptor expressed on the surface of hematopoietic stem cells and other cell types | Signaling through KIT plays a role in cell survival, proliferation, and differentiation. For instance, KIT signaling is required for melanocyte survival, and it is also involved in hematopoiesis and gametogenesis | Positive | [45,46] |
| CD146 | Melanoma cell adhesion molecule (MCAM), cell surface glycoprotein (MUC18) | Activated human T cells, endothelial progenitors such as angioblasts and mesenchymal stem cells, and strongly expressed blood vessel endothelium and smooth muscle | CD146 has been seen as a marker for mesenchymal stem cells isolated from multiple adult and fetal organs, and its expression may be linked to multipotency; mesenchymal stem cells with greater differentiation potential express higher levels of CD146 on the cell surface | Positive | [41,67,68] |
| CD166 | Activated leukocyte cell adhesion molecule (ALCAM) | T cells, NK cells, platelets, thymocytes, activated B and T cells, eosinophils, fibroblasts, endothelial cells, keratinocytes, monocytes, epithelial cells, neurons, MSCs | Adhesion molecule, ligand for CD6; T-cell activation. Involved in neurite extension by neurons via heterophilic and homophilic interactions | Positive | [41,69,70] |
| CD 271 | Low-affinity nerve growth factor receptor (LNGFR), p75 neurotrophin receptor | This receptor is currently considered one of the most efficient markers for prospectively isolate BM-MSCs and other adult stem cells | It is a common receptor of neurotrophins that mediates different biological effects in several cell types, like cell survival, apoptosis, migration, and differentiation, and neurotrophin binding, interacting transmembrane co-receptors expression, intracellular adaptor molecule availability, and post-translational modifications, such as regulated proteolytic processing | Positive | [71] |
| MHC class I | Major histocompatibility complex (MHC) class I | MHC molecules are found on the cell surface of all nucleated cells in vertebrates | Major histocompatibility complex (MHC) class I and class II proteins play a pivotal role in the immune system’s adaptive branch. Both classes of proteins share the task of presenting peptides on the cell surface for recognition by T cells | Positive | [72] |
| MHC class II | Major histocompatibility complex (MHC) class I | MHC class II molecules are found only on professional antigen-presenting cells such as dendritic cells, mononuclear phagocytes, some endothelial cells, thymic epithelial cells, and B cells | Negative | [73] | |
| STRO1 | A gene for a protein marker of mesenchymal stem cells (MSC) | Marker for mesenchymal stem cells. | A gene for a protein marker of mesenchymal stem cells (MSC) | Positive | [41,74] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pilbauerova, N.; Schmidt, J.; Soukup, T.; Koberova Ivancakova, R.; Suchanek, J. The Effects of Cryogenic Storage on Human Dental Pulp Stem Cells. Int. J. Mol. Sci. 2021, 22, 4432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094432
Pilbauerova N, Schmidt J, Soukup T, Koberova Ivancakova R, Suchanek J. The Effects of Cryogenic Storage on Human Dental Pulp Stem Cells. International Journal of Molecular Sciences. 2021; 22(9):4432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094432
Chicago/Turabian StylePilbauerova, Nela, Jan Schmidt, Tomas Soukup, Romana Koberova Ivancakova, and Jakub Suchanek. 2021. "The Effects of Cryogenic Storage on Human Dental Pulp Stem Cells" International Journal of Molecular Sciences 22, no. 9: 4432. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094432