
A Yeast-Based Screening Unravels Potential Therapeutic Molecules for Mitochondrial Diseases Associated with Dominant ANT1 Mutations

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
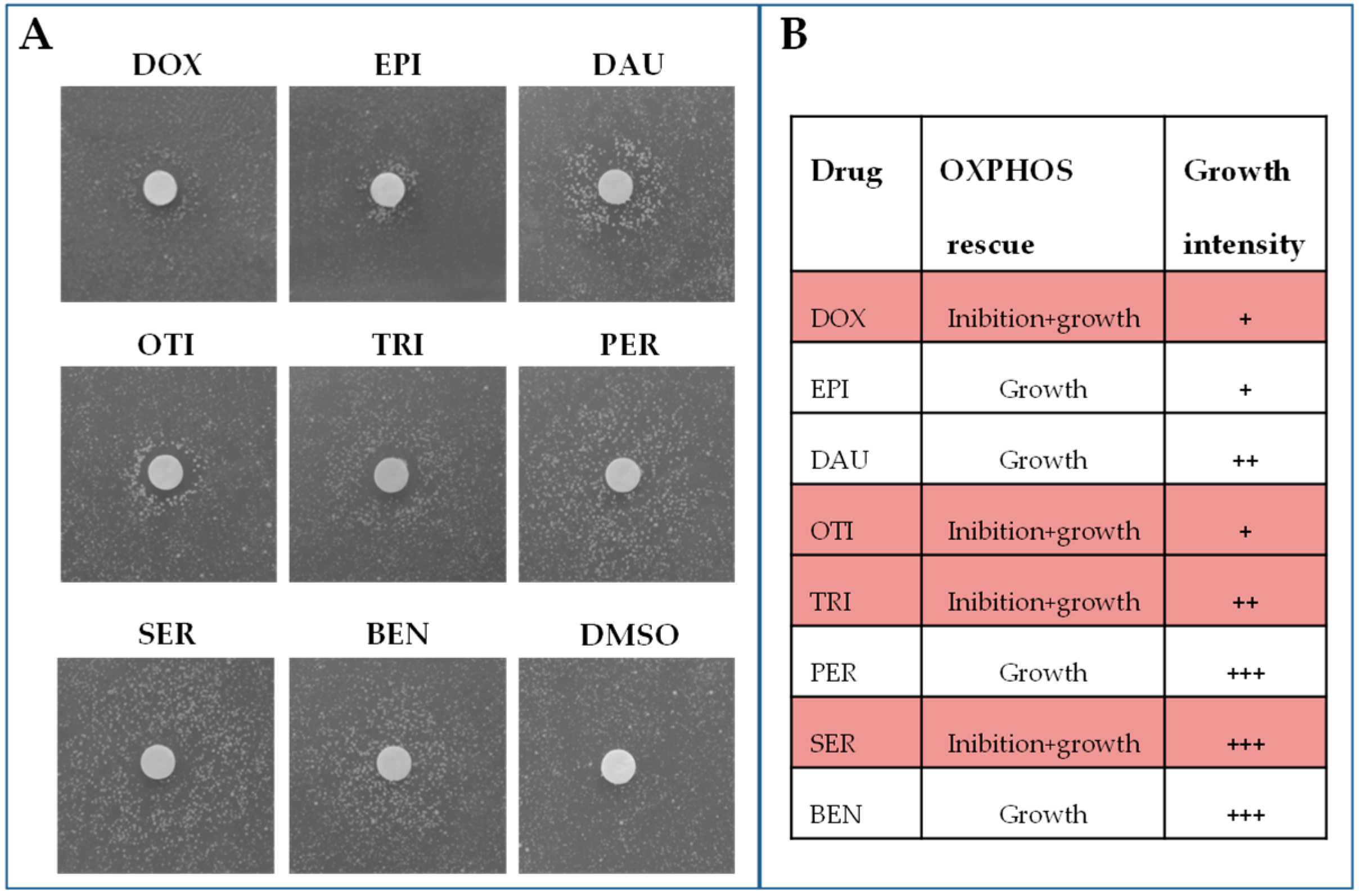
2.1. Identification of Molecules Able to Rescue the Oxidative Growth Defect of the aac2M114P Yeast Mutant
- -
- Compounds that lead to the formation of a halo of growth near the filter. These molecules have a rescuing effect starting from the maximum concentration tested. This was the case of pergolide mesylate and benzydamine HCl.
- -
- Compounds that lead to the formation of an external crown of growth, where no growth is observed near the filter. These molecules are toxic at high concentrations (near the filter) and active at lower concentrations (far from the filter). This was the case of otilonium bromide, trifluoperazine 2HCl, and sertraline HCl.
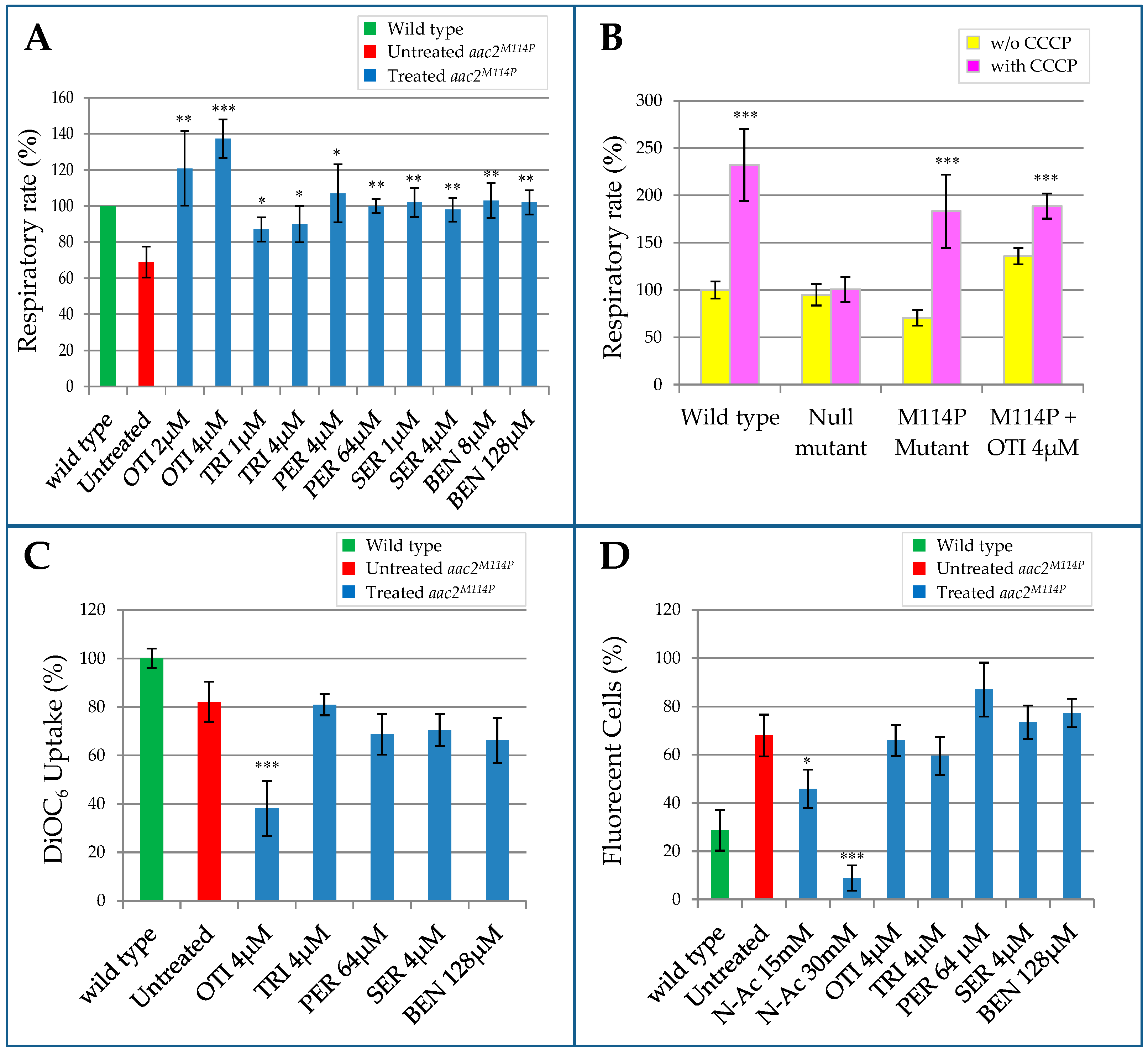
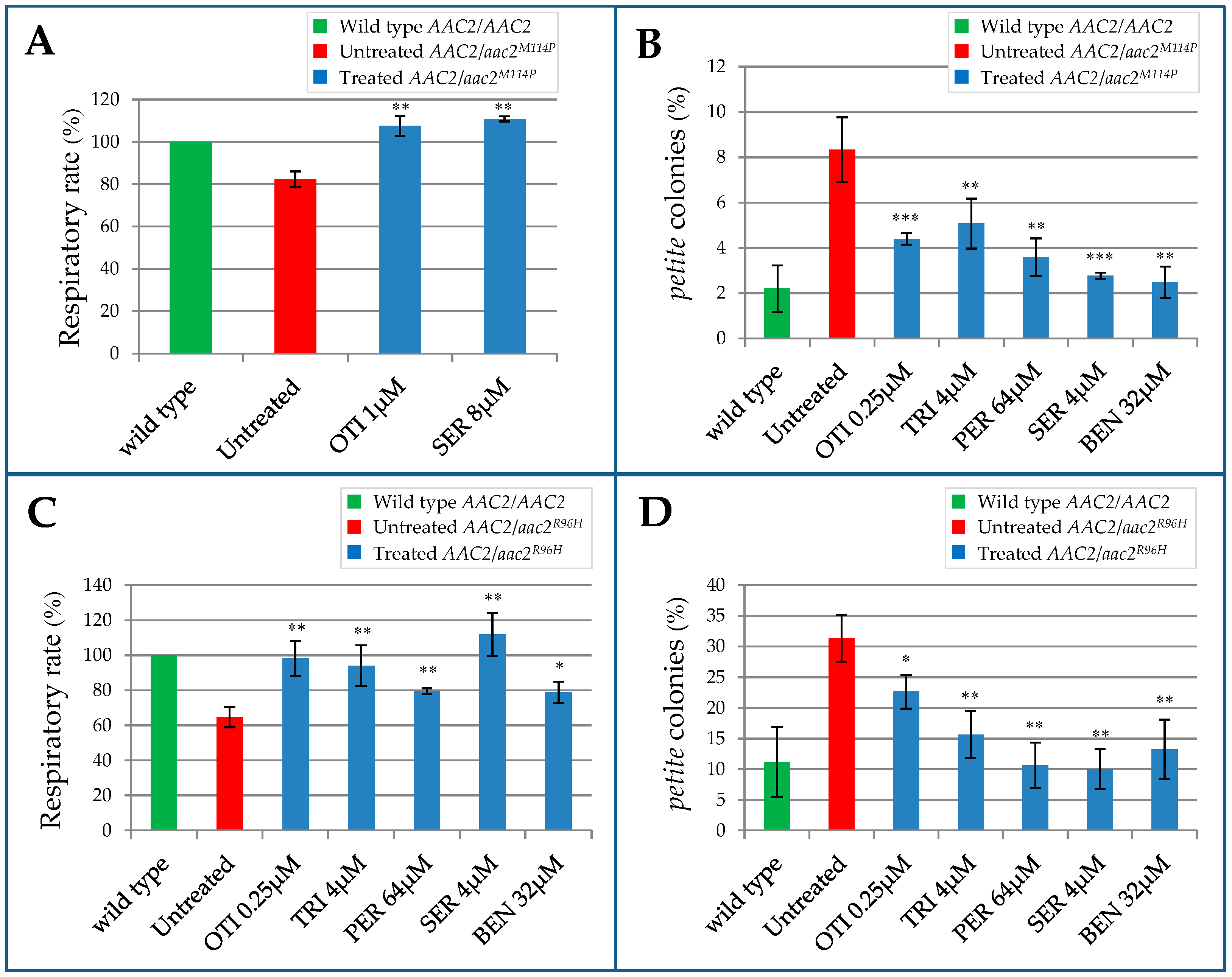
2.2. Effect of the Identified Drugs on WB-12/aac2M114P Phenotypes
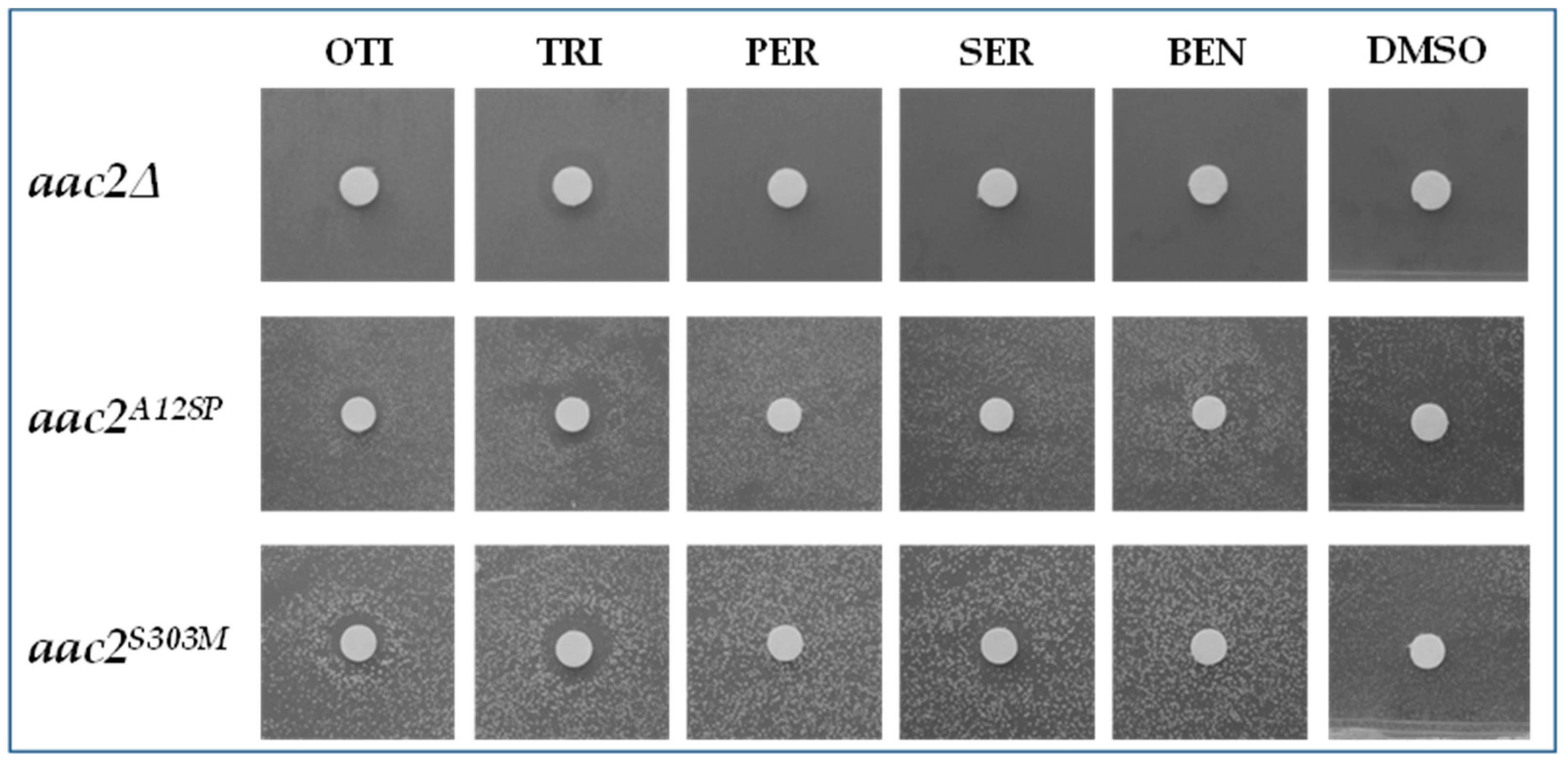
2.3. Evaluation of the Drugs Effect on Other aac2 Pathological Mutants
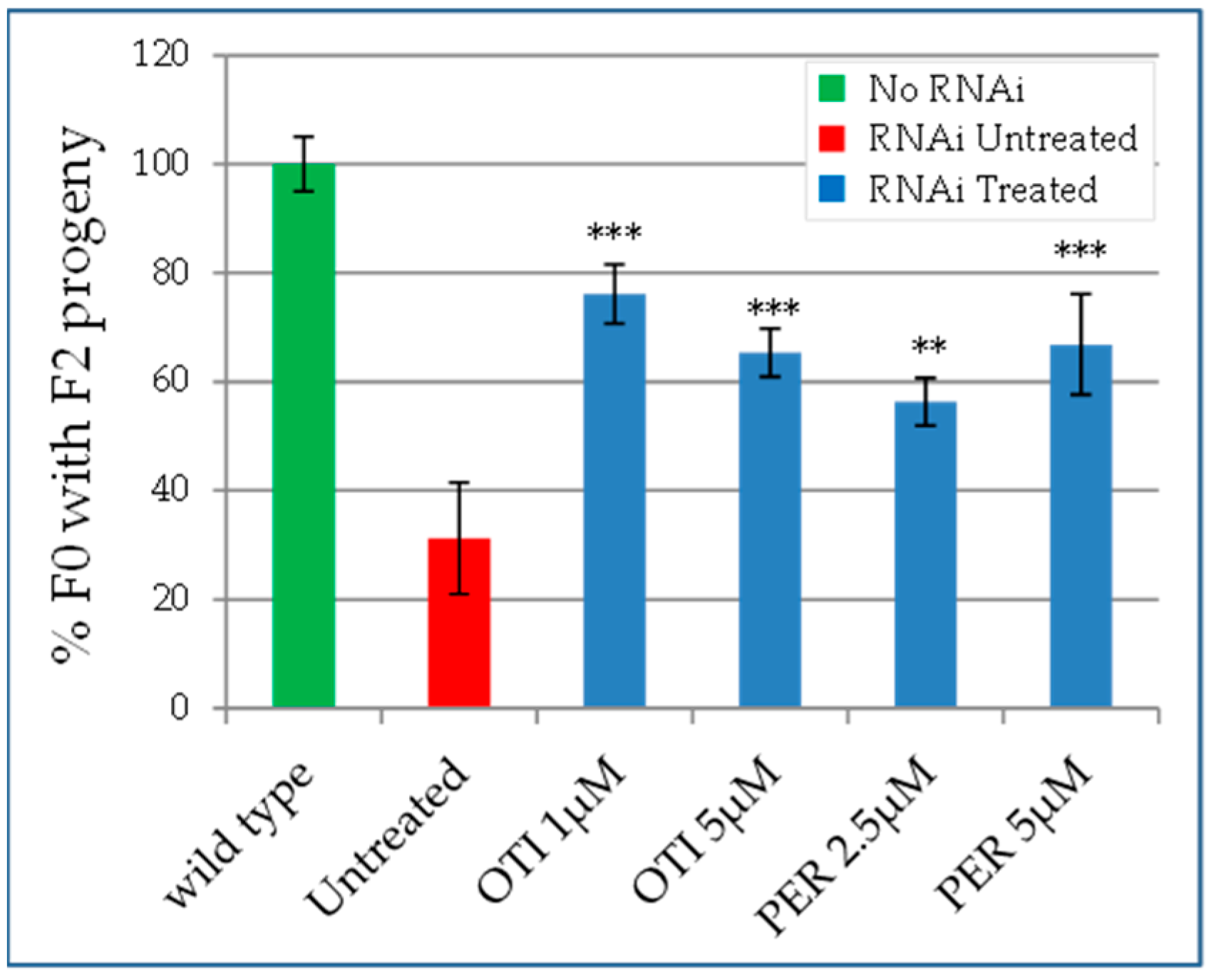
2.4. Characterization of the Drugs Effect on Caenorhabditis elegans
2.5. Effect of the Identified Drugs on Heteroallelic Strains
3. Discussion
4. Materials and Methods
4.1. Yeast Strains and Culture Media
4.2. High Throughput Screening: Drug Drop Test
4.3. Determination of the Minimal Inhibition Concentration
4.4. Respiratory Activity
4.5. Petite Frequency and Fitness Test
4.6. Flow Cytometric Analysis
4.7. Protein Extraction and Western Blot Analysis
4.8. Reconstitution of Aac2 into Liposomes and Transport Measurements
4.9. RNA Interference and Analysis of Embryonic Lethality in C. elegans
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Munnich, A.; Rustin, P.; Rötig, A.; Chretien, D.; Bonnefont, J.P.; Nuttin, C.; Cormier, V.; Vassault, A.; Parvy, P.; Bardet, J. Clinical Aspects of Mitochondrial Disorders. J. Inherit. Metab. Dis. 1992, 15, 448–455. [Google Scholar] [CrossRef] [PubMed]
- Zeviani, M.; Di Donato, S. Mitochondrial Disorders. Brain J. Neurol. 2004, 127, 2153–2172. [Google Scholar] [CrossRef] [PubMed]
- Lightowlers, R.N.; Taylor, R.W.; Turnbull, D.M. Mutations Causing Mitochondrial Disease: What Is New and What Challenges Remain? Science 2015, 349, 1494–1499. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.; Collier, J.J.; Glasgow, R.I.C.; Robertson, F.M.; Pyle, A.; Blakely, E.L.; Alston, C.L.; Oláhová, M.; McFarland, R.; Taylor, R.W. Recent Advances in Understanding the Molecular Genetic Basis of Mitochondrial Disease. J. Inherit. Metab. Dis. 2020, 43, 36–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenton, S.L.; Prokisch, H. Advancing Genomic Approaches to the Molecular Diagnosis of Mitochondrial Disease. Essays Biochem. 2018, 62, 399–408. [Google Scholar] [CrossRef]
- Frazier, A.E.; Thorburn, D.R.; Compton, A.G. Mitochondrial Energy Generation Disorders: Genes, Mechanisms, and Clues to Pathology. J. Biol. Chem. 2019, 294, 5386–5395. [Google Scholar] [CrossRef] [Green Version]
- Dolce, V.; Scarcia, P.; Iacopetta, D.; Palmieri, F. A Fourth ADP/ATP Carrier Isoform in Man: Identification, Bacterial Expression, Functional Characterization and Tissue Distribution. FEBS Lett. 2005, 579, 633–637. [Google Scholar] [CrossRef] [Green Version]
- Klingenberg, M. The ADP and ATP Transport in Mitochondria and Its Carrier. Biochim. Biophys. Acta 2008, 1778, 1978–2021. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, F. The Mitochondrial Transporter Family (SLC25): Physiological and Pathological Implications. Pflugers Arch. 2004, 447, 689–709. [Google Scholar] [CrossRef]
- Palmieri, F. Mitochondrial Transporters of the SLC25 Family and Associated Diseases: A Review. J. Inherit. Metab. Dis. 2014, 37, 565–575. [Google Scholar] [CrossRef]
- Pebay-Peyroula, E.; Dahout-Gonzalez, C.; Kahn, R.; Trézéguet, V.; Lauquin, G.J.-M.; Brandolin, G. Structure of Mitochondrial ADP/ATP Carrier in Complex with Carboxyatractyloside. Nature 2003, 426, 39–44. [Google Scholar] [CrossRef]
- Kunji, E.R.S.; Aleksandrova, A.; King, M.S.; Majd, H.; Ashton, V.L.; Cerson, E.; Springett, R.; Kibalchenko, M.; Tavoulari, S.; Crichton, P.G.; et al. The Transport Mechanism of the Mitochondrial ADP/ATP Carrier. Biochim. Biophys. Acta 2016, 1863, 2379–2393. [Google Scholar] [CrossRef] [Green Version]
- Ruprecht, J.J.; Kunji, E.R.S. The SLC25 Mitochondrial Carrier Family: Structure and Mechanism. Trends Biochem. Sci. 2020, 45, 244–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deschauer, M.; Hudson, G.; Müller, T.; Taylor, R.W.; Chinnery, P.F.; Zierz, S. A Novel ANT1 Gene Mutation with Probable Germline Mosaicism in Autosomal Dominant Progressive External Ophthalmoplegia. Neuromuscul. Disord. 2005, 15, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Napoli, L.; Bordoni, A.; Zeviani, M.; Hadjigeorgiou, G.M.; Sciacco, M.; Tiranti, V.; Terentiou, A.; Moggio, M.; Papadimitriou, A.; Scarlato, G.; et al. A Novel Missense Adenine Nucleotide Translocator-1 Gene Mutation in a Greek AdPEO Family. Neurology 2001, 57, 2295–2298. [Google Scholar] [CrossRef] [PubMed]
- Siciliano, G.; Tessa, A.; Petrini, S.; Mancuso, M.; Bruno, C.; Grieco, G.S.; Malandrini, A.; DeFlorio, L.; Martini, B.; Federico, A.; et al. Autosomal Dominant External Ophthalmoplegia and Bipolar Affective Disorder Associated with a Mutation in the ANT1 Gene. Neuromuscul. Disord. 2003, 13, 162–165. [Google Scholar] [CrossRef]
- Komaki, H.; Fukazawa, T.; Houzen, H.; Yoshida, K.; Nonaka, I.; Goto, Y. A Novel D104G Mutation in the Adenine Nucleotide Translocator 1 Gene in Autosomal Dominant Progressive External Ophthalmoplegia Patients with Mitochondrial DNA with Multiple Deletions. Ann. Neurol. 2002, 51, 645–648. [Google Scholar] [CrossRef] [PubMed]
- Kaukonen, J.; Juselius, J.K.; Tiranti, V.; Kyttälä, A.; Zeviani, M.; Comi, G.P.; Keränen, S.; Peltonen, L.; Suomalainen, A. Role of Adenine Nucleotide Translocator 1 in MtDNA Maintenance. Science 2000, 289, 782–785. [Google Scholar] [CrossRef]
- Palmieri, L.; Alberio, S.; Pisano, I.; Lodi, T.; Meznaric-Petrusa, M.; Zidar, J.; Santoro, A.; Scarcia, P.; Fontanesi, F.; Lamantea, E.; et al. Complete Loss-of-Function of the Heart/Muscle-Specific Adenine Nucleotide Translocator Is Associated with Mitochondrial Myopathy and Cardiomyopathy. Hum. Mol. Genet. 2005, 14, 3079–3088. [Google Scholar] [CrossRef] [Green Version]
- Körver-Keularts, I.M.L.W.; de Visser, M.; Bakker, H.D.; Wanders, R.J.A.; Vansenne, F.; Scholte, H.R.; Dorland, L.; Nicolaes, G.A.F.; Spaapen, L.M.J.; Smeets, H.J.M.; et al. Two Novel Mutations in the SLC25A4 Gene in a Patient with Mitochondrial Myopathy. JIMD Rep. 2015, 22, 39–45. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.; Majd, H.; Dallabona, C.; Reinson, K.; King, M.S.; Alston, C.L.; He, L.; Lodi, T.; Jones, S.A.; Fattal-Valevski, A.; et al. Recurrent De Novo Dominant Mutations in SLC25A4 Cause Severe Early-Onset Mitochondrial Disease and Loss of Mitochondrial DNA Copy Number. Am. J. Hum. Genet. 2016, 99, 1405. [Google Scholar] [CrossRef]
- King, M.S.; Thompson, K.; Hopton, S.; He, L.; Kunji, E.R.S.; Taylor, R.W.; Ortiz-Gonzalez, X.R. Expanding the Phenotype of de Novo SLC25A4-Linked Mitochondrial Disease to Include Mild Myopathy. Neurol. Genet. 2018, 4, e256. [Google Scholar] [CrossRef] [Green Version]
- Lawson, J.E.; Douglas, M.G. Separate Genes Encode Functionally Equivalent ADP/ATP Carrier Proteins in Saccharomyces cerevisiae. Isolation and Analysis of AAC2. J. Biol. Chem. 1988, 263, 14812–14818. [Google Scholar] [CrossRef]
- Kolarov, J.; Kolarova, N.; Nelson, N. A Third ADP/ATP Translocator Gene in Yeast. J. Biol. Chem. 1990, 265, 12711–12716. [Google Scholar] [CrossRef]
- Lasserre, J.-P.; Dautant, A.; Aiyar, R.S.; Kucharczyk, R.; Glatigny, A.; Tribouillard-Tanvier, D.; Rytka, J.; Blondel, M.; Skoczen, N.; Reynier, P.; et al. Yeast as a System for Modeling Mitochondrial Disease Mechanisms and Discovering Therapies. Dis. Model. Mech. 2015, 8, 509–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceccatelli Berti, C.; di Punzio, G.; Dallabona, C.; Baruffini, E.; Goffrini, P.; Lodi, T.; Donnini, C. The Power of Yeast in Modelling Human Nuclear Mutations Associated with Mitochondrial Diseases. Genes 2021, 12, 300. [Google Scholar] [CrossRef]
- Fontanesi, F.; Palmieri, L.; Scarcia, P.; Lodi, T.; Donnini, C.; Limongelli, A.; Tiranti, V.; Zeviani, M.; Ferrero, I.; Viola, A.M. Mutations in AAC2, Equivalent to Human AdPEO-Associated ANT1 Mutations, Lead to Defective Oxidative Phosphorylation in Saccharomyces cerevisiae and Affect Mitochondrial DNA Stability. Hum. Mol. Genet. 2004, 13, 923–934. [Google Scholar] [CrossRef] [PubMed]
- Lodi, T.; Bove, C.; Fontanesi, F.; Viola, A.M.; Ferrero, I. Mutation D104G in ANT1 Gene: Complementation Study in Saccharomyces cerevisiae as a Model System. Biochem. Biophys. Res. Commun. 2006, 341, 810–815. [Google Scholar] [CrossRef]
- Couplan, E.; Aiyar, R.S.; Kucharczyk, R.; Kabala, A.; Ezkurdia, N.; Gagneur, J.; St Onge, R.P.; Salin, B.; Soubigou, F.; Le Cann, M.; et al. A Yeast-Based Assay Identifies Drugs Active against Human Mitochondrial Disorders. Proc. Natl. Acad. Sci. USA 2011, 108, 11989–11994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viscomi, C.; Zeviani, M. Strategies for Fighting Mitochondrial Diseases. J. Intern. Med. 2020, 287, 665–684. [Google Scholar] [CrossRef]
- Pitayu, L.; Baruffini, E.; Rodier, C.; Rötig, A.; Lodi, T.; Delahodde, A. Combined Use of Saccharomyces cerevisiae, Caenorhabditis elegans and Patient Fibroblasts Leads to the Identification of Clofilium Tosylate as a Potential Therapeutic Chemical against POLG-Related Diseases. Hum. Mol. Genet. 2016, 25, 715–727. [Google Scholar] [CrossRef] [Green Version]
- Panozzo, C.; Laleve, A.; Tribouillard-Tanvier, D.; Ostojić, J.; Sellem, C.H.; Friocourt, G.; Bourand-Plantefol, A.; Burg, A.; Delahodde, A.; Blondel, M.; et al. Chemicals or Mutations That Target Mitochondrial Translation Can Rescue the Respiratory Deficiency of Yeast Bcs1 Mutants. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 2297–2307. [Google Scholar] [CrossRef]
- Delerue, T.; Tribouillard-Tanvier, D.; Daloyau, M.; Khosrobakhsh, F.; Emorine, L.J.; Friocourt, G.; Belenguer, P.; Blondel, M.; Arnauné-Pelloquin, L. A Yeast-Based Screening Assay Identifies Repurposed Drugs That Suppress Mitochondrial Fusion and MtDNA Maintenance Defects. Dis. Model. Mech. 2019, 12. [Google Scholar] [CrossRef] [Green Version]
- Aleo, S.J.; Del Dotto, V.; Fogazza, M.; Maresca, A.; Lodi, T.; Goffrini, P.; Ghelli, A.; Rugolo, M.; Carelli, V.; Baruffini, E.; et al. Drug Repositioning as a Therapeutic Strategy for Neurodegenerations Associated with OPA1 Mutations. Hum. Mol. Genet. 2021, 29, 3631–3645. [Google Scholar] [CrossRef] [PubMed]
- Traba, J.; Satrústegui, J.; del Arco, A. Transport of Adenine Nucleotides in the Mitochondria of Saccharomyces cerevisiae: Interactions between the ADP/ATP Carriers and the ATP-Mg/Pi Carrier. Mitochondrion 2009, 9, 79–85. [Google Scholar] [CrossRef]
- Farina, F.; Alberti, A.; Breuil, N.; Bolotin-Fukuhara, M.; Pinto, M.; Culetto, E. Differential Expression Pattern of the Four Mitochondrial Adenine Nucleotide Transporter Ant Genes and Their Roles during the Development of Caenorhabditis elegans. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2008, 237, 1668–1681. [Google Scholar] [CrossRef]
- Shen, Q.; Qin, F.; Gao, Z.; Cui, J.; Xiao, H.; Xu, Z.; Yang, C. Adenine Nucleotide Translocator Cooperates with Core Cell Death Machinery to Promote Apoptosis in Caenorhabditis elegans. Mol. Cell. Biol. 2009, 29, 3881–3893. [Google Scholar] [CrossRef] [Green Version]
- Dujon, B. Mitochondrial genetics and functions. In The Molecular Biology of the Yeast Saccharomyces. Life Cycle and Inheritance; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1981; pp. 505–635. [Google Scholar]
- Baruffini, E.; Ferrero, I.; Foury, F. In Vivo Analysis of MtDNA Replication Defects in Yeast. Methods 2010, 51, 426–436. [Google Scholar] [CrossRef]
- Dallabona, C.; Baruffini, E.; Goffrini, P.; Lodi, T. Dominance of Yeast aac2R96H and aac2R252G Mutations, Equivalent to Pathological Mutations in Ant1, Is Due to Gain of Function. Biochem. Biophys. Res. Commun. 2017, 493, 909–913. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.D.; Pakay, J.L.; Ocloo, A.; Kokoszka, J.; Wallace, D.C.; Brookes, P.S.; Cornwall, E.J. The Basal Proton Conductance of Mitochondria Depends on Adenine Nucleotide Translocase Content. Biochem. J. 2005, 392, 353–362. [Google Scholar] [CrossRef] [Green Version]
- Appleby, R.D.; Porteous, W.K.; Hughes, G.; James, A.M.; Shannon, D.; Wei, Y.H.; Murphy, M.P. Quantitation and Origin of the Mitochondrial Membrane Potential in Human Cells Lacking Mitochondrial DNA. Eur. J. Biochem. 1999, 262, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Zoratti, M.; Szabò, I. The Mitochondrial Permeability Transition. Biochim. Biophys. Acta 1995, 1241, 139–176. [Google Scholar] [CrossRef]
- Kokoszka, J.E.; Waymire, K.G.; Levy, S.E.; Sligh, J.E.; Cai, J.; Jones, D.P.; MacGregor, G.R.; Wallace, D.C. The ADP/ATP Translocator Is Not Essential for the Mitochondrial Permeability Transition Pore. Nature 2004, 427, 461–465. [Google Scholar] [CrossRef]
- Marzo, I.; Brenner, C.; Zamzami, N.; Jürgensmeier, J.M.; Susin, S.A.; Vieira, H.L.; Prévost, M.C.; Xie, Z.; Matsuyama, S.; Reed, J.C.; et al. Bax and Adenine Nucleotide Translocator Cooperate in the Mitochondrial Control of Apoptosis. Science 1998, 281, 2027–2031. [Google Scholar] [CrossRef] [PubMed]
- Hoshino, A.; Wang, W.-J.; Wada, S.; McDermott-Roe, C.; Evans, C.S.; Gosis, B.; Morley, M.P.; Rathi, K.S.; Li, J.; Li, K.; et al. The ADP/ATP Translocase Drives Mitophagy Independent of Nucleotide Exchange. Nature 2019, 575, 375–379. [Google Scholar] [CrossRef]
- Bauer, M.K.; Schubert, A.; Rocks, O.; Grimm, S. Adenine Nucleotide Translocase-1, a Component of the Permeability Transition Pore, Can Dominantly Induce Apoptosis. J. Cell Biol. 1999, 147, 1493–1502. [Google Scholar] [CrossRef]
- Kawamata, H.; Tiranti, V.; Magrané, J.; Chinopoulos, C.; Manfredi, G. AdPEO Mutations in ANT1 Impair ADP-ATP Translocation in Muscle Mitochondria. Hum. Mol. Genet. 2011, 20, 2964–2974. [Google Scholar] [CrossRef] [Green Version]
- El-Khoury, R.; Sainsard-Chanet, A. Suppression of Mitochondrial DNA Instability of Autosomal Dominant Forms of Progressive External Ophthalmoplegia-Associated ANT1 Mutations in Podospora anserina. Genetics 2009, 183, 861–871. [Google Scholar] [CrossRef] [Green Version]
- Rainey, M.M.; Korostyshevsky, D.; Lee, S.; Perlstein, E.O. The Antidepressant Sertraline Targets Intracellular Vesiculogenic Membranes in Yeast. Genetics 2010, 185, 1221–1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, W.H. Cationic Amphiphilic Drug-Induced Phospholipidosis. Toxicol. Pathol. 1997, 25, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Mileykovskaya, E.; Dowhan, W. Gluing the Respiratory Chain Together. Cardiolipin Is Required for Supercomplex Formation in the Inner Mitochondrial Membrane. J. Biol. Chem. 2002, 277, 43553–43556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfeiffer, K.; Gohil, V.; Stuart, R.A.; Hunte, C.; Brandt, U.; Greenberg, M.L.; Schägger, H. Cardiolipin Stabilizes Respiratory Chain Supercomplexes. J. Biol. Chem. 2003, 278, 52873–52880. [Google Scholar] [CrossRef] [Green Version]
- Claypool, S.M.; Oktay, Y.; Boontheung, P.; Loo, J.A.; Koehler, C.M. Cardiolipin Defines the Interactome of the Major ADP/ATP Carrier Protein of the Mitochondrial Inner Membrane. J. Cell Biol. 2008, 182, 937–950. [Google Scholar] [CrossRef] [Green Version]
- Jiang, F.; Ryan, M.T.; Schlame, M.; Zhao, M.; Gu, Z.; Klingenberg, M.; Pfanner, N.; Greenberg, M.L. Absence of Cardiolipin in the Crd1 Null Mutant Results in Decreased Mitochondrial Membrane Potential and Reduced Mitochondrial Function. J. Biol. Chem. 2000, 275, 22387–22394. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, X.; Chen, X.J. Misfolding of Mutant Adenine Nucleotide Translocase in Yeast Supports a Novel Mechanism of Ant1-Induced Muscle Diseases. Mol. Biol. Cell 2015, 26, 1985–1994. [Google Scholar] [CrossRef] [PubMed]
- Miyakawa, I. Organization and Dynamics of Yeast Mitochondrial Nucleoids. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2017, 93, 339–359. [Google Scholar] [CrossRef] [Green Version]
- Senoo, N.; Kandasamy, S.; Ogunbona, O.B.; Baile, M.G.; Lu, Y.; Claypool, S.M. Cardiolipin, Conformation, and Respiratory Complex-Dependent Oligomerization of the Major Mitochondrial ADP/ATP Carrier in Yeast. Sci. Adv. 2020, 6, eabb0780. [Google Scholar] [CrossRef]
- Quane, P.A.; Graham, G.G.; Ziegler, J.B. Pharmacology of Benzydamine. Inflammopharmacology 1998, 6, 95–107. [Google Scholar] [CrossRef] [PubMed]
- Opaleye, E.S.; Noto, A.R.; Sanchez, Z.; van der Meer Sanchez, Z.; de Moura, Y.G.; Galduróz, J.C.F.; Carlini, E.A. Recreational Use of Benzydamine as a Hallucinogen among Street Youth in Brazil. Revista Brasileira de Psiquiatria 2009, 31, 208–213. [Google Scholar] [CrossRef] [Green Version]
- Stefania, C.; Andrea, M.; Alessio, M.; Mauro, P.; Amira, G.; Martin, C.J.; Giovanni, M.; Massimo, D.G.; Fabrizio, S. The Benzydamine Experience: A Systematic Review of Benzydamine Abuse. Curr. Neuropharmacol. 2021. [Google Scholar] [CrossRef]
- Eilam, Y.; Polacheck, I.; Ben-Gigi, G.; Chernichovsky, D. Activity of Phenothiazines against Medically Important Yeasts. Antimicrob. Agents Chemother. 1987, 31, 834–836. [Google Scholar] [CrossRef] [Green Version]
- Siavoshi, F.; Tavakolian, A.; Foroumadi, A.; Hosseini, N.M.; Massarrat, S.; Pedramnia, S.; Saniee, P. Comparison of the Effect of Non-Antifungal and Antifungal Agents on Candida Isolates from the Gastrointestinal Tract. Arch. Iran. Med. 2012, 15, 27–31. [Google Scholar] [PubMed]
- Sharma, S.; Kaur, H.; Khuller, G.K. Cell Cycle Effects of the Phenothiazines: Trifluoperazine and Chlorpromazine in Candida Albicans. FEMS Microbiol. Lett. 2001, 199, 185–190. [Google Scholar] [CrossRef] [PubMed]
- Eilam, Y. Membrane Effects of Phenothiazines in Yeasts. I. Stimulation of Calcium and Potassium Fluxes. Biochim. Biophys. Acta 1983, 733, 242–248. [Google Scholar] [CrossRef]
- Hong, Z.; Smith, A.J.; Archer, S.L.; Wu, X.-C.; Nelson, D.P.; Peterson, D.; Johnson, G.; Weir, E.K. Pergolide Is an Inhibitor of Voltage-Gated Potassium Channels, Including Kv1.5, and Causes Pulmonary Vasoconstriction. Circulation 2005, 112, 1494–1499. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J. Sal1p, a Calcium-Dependent Carrier Protein That Suppresses an Essential Cellular Function Associated with the Aac2 Isoform of ADP/ATP Translocase in Saccharomyces cerevisiae. Genetics 2004, 167, 607–617. [Google Scholar] [CrossRef] [Green Version]
- Traba, J.; Froschauer, E.M.; Wiesenberger, G.; Satrústegui, J.; Del Arco, A. Yeast Mitochondria Import ATP through the Calcium-Dependent ATP-Mg/Pi Carrier Sal1p, and Are ATP Consumers during Aerobic Growth in Glucose. Mol. Microbiol. 2008, 69, 570–585. [Google Scholar] [CrossRef]
- Laco, J.; Zeman, I.; Pevala, V.; Polcic, P.; Kolarov, J. Adenine Nucleotide Transport via Sal1 Carrier Compensates for the Essential Function of the Mitochondrial ADP/ATP Carrier. FEMS Yeast Res. 2010, 10, 290–296. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.J.; Rothstein, R. Elevated Recombination Rates in Transcriptionally Active DNA. Cell 1989, 56, 619–630. [Google Scholar] [CrossRef]
- Hashimoto, M.; Shinohara, Y.; Majima, E.; Hatanaka, T.; Yamazaki, N.; Terada, H. Expression of the Bovine Heart Mitochondrial ADP/ATP Carrier in Yeast Mitochondria: Significantly Enhanced Expression by Replacement of the N-Terminal Region of the Bovine Carrier by the Corresponding Regions of the Yeast Carriers. Biochim. Biophys. Acta 1999, 1409, 113–124. [Google Scholar] [CrossRef] [Green Version]
- Gomez, M.; Pérez-Gallardo, R.V.; Sánchez, L.A.; Díaz-Pérez, A.L.; Cortés-Rojo, C.; Meza Carmen, V.; Saavedra-Molina, A.; Lara-Romero, J.; Jiménez-Sandoval, S.; Rodríguez, F.; et al. Malfunctioning of the Iron-Sulfur Cluster Assembly Machinery in Saccharomyces cerevisiae Produces Oxidative Stress via an Iron-Dependent Mechanism, Causing Dysfunction in Respiratory Complexes. PLoS ONE 2014, 9, e111585. [Google Scholar] [CrossRef] [Green Version]
- Miceli, M.V.; Jiang, J.C.; Tiwari, A.; Rodriguez-Quiñones, J.F.; Jazwinski, S.M. Loss of Mitochondrial Membrane Potential Triggers the Retrograde Response Extending Yeast Replicative Lifespan. Front. Genet. 2011, 2, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmieri, F.; Indiveri, C.; Bisaccia, F.; Iacobazzi, V. Mitochondrial Metabolite Carrier Proteins: Purification, Reconstitution, and Transport Studies. Methods Enzymol. 1995, 260, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Brenner, S. The Genetics of Caenorhabditis Elegans. Genetics 1974, 77, 71–94. [Google Scholar] [CrossRef]
- Kamath, R.S.; Ahringer, J. Genome-Wide RNAi Screening in Caenorhabditis elegans. Methods 2003, 30, 313–321. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
di Punzio, G.; Di Noia, M.A.; Delahodde, A.; Sellem, C.; Donnini, C.; Palmieri, L.; Lodi, T.; Dallabona, C. A Yeast-Based Screening Unravels Potential Therapeutic Molecules for Mitochondrial Diseases Associated with Dominant ANT1 Mutations. Int. J. Mol. Sci. 2021, 22, 4461. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094461
di Punzio G, Di Noia MA, Delahodde A, Sellem C, Donnini C, Palmieri L, Lodi T, Dallabona C. A Yeast-Based Screening Unravels Potential Therapeutic Molecules for Mitochondrial Diseases Associated with Dominant ANT1 Mutations. International Journal of Molecular Sciences. 2021; 22(9):4461. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094461
Chicago/Turabian Styledi Punzio, Giulia, Maria Antonietta Di Noia, Agnès Delahodde, Carole Sellem, Claudia Donnini, Luigi Palmieri, Tiziana Lodi, and Cristina Dallabona. 2021. "A Yeast-Based Screening Unravels Potential Therapeutic Molecules for Mitochondrial Diseases Associated with Dominant ANT1 Mutations" International Journal of Molecular Sciences 22, no. 9: 4461. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094461