
Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Description and Utilization of Hops
2.1. History of Hops
2.2. Hop Botany
2.3. Hop Cultivation
2.4. XH and Brewing
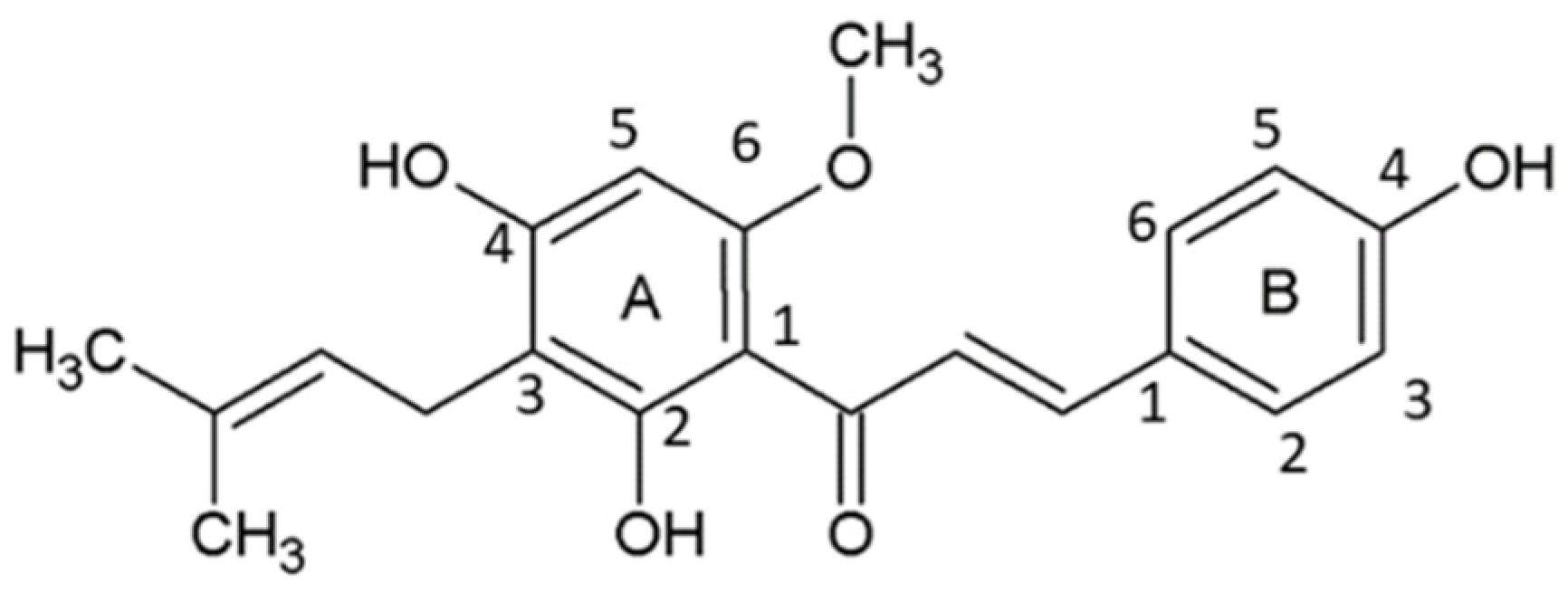
3. Chemistry and SAR of XH
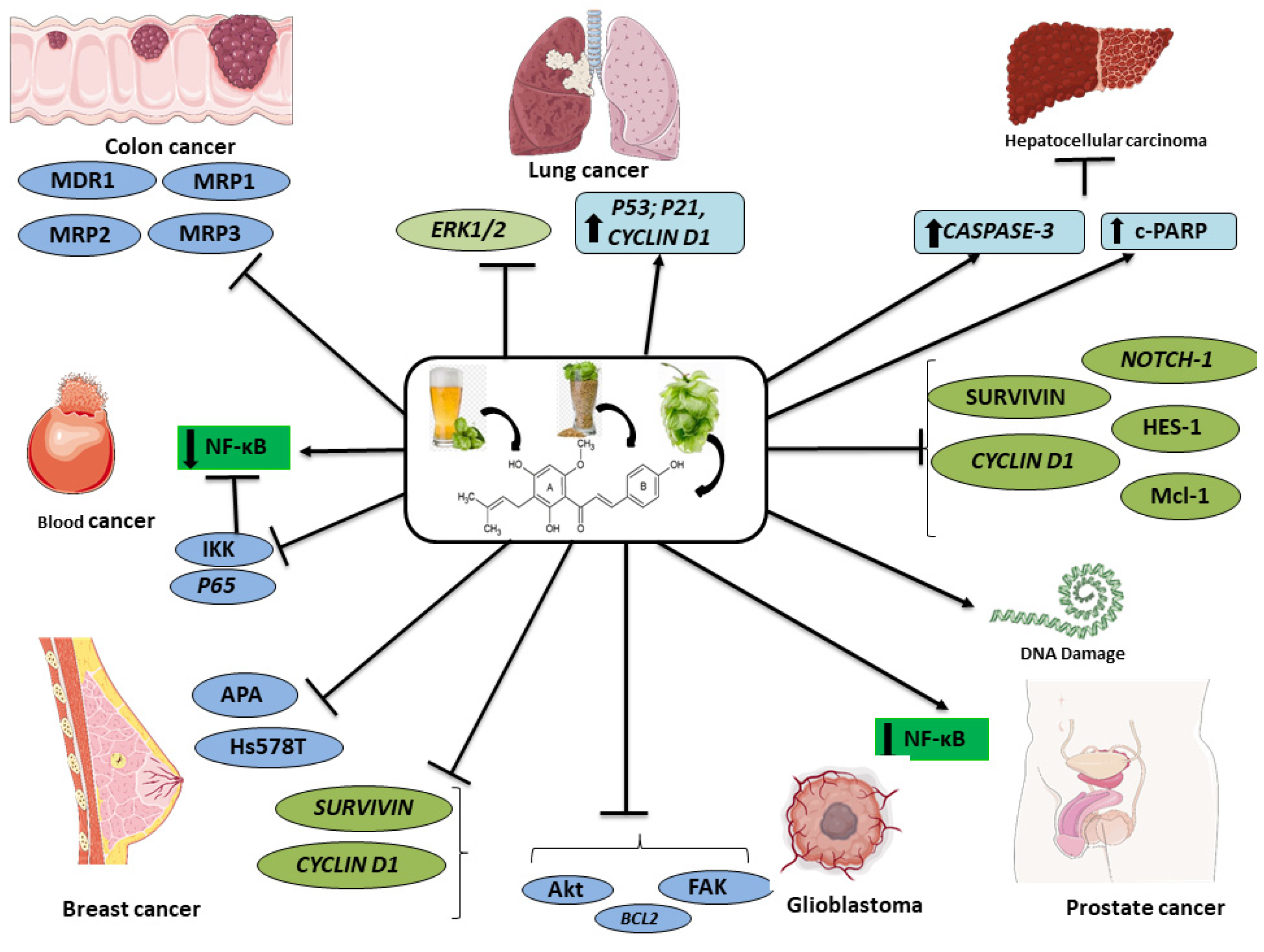
4. Anticancer Potential of XH Based on Preclinical Evidence
4.1. Breast Cancer
4.2. Cervical Cancer
4.3. Cholangiocarcinoma
4.4. Colon Cancer
4.5. Esophageal Cancer
4.6. Glioblastoma
4.7. Hematological Cancers
4.8. Laryngeal Cancer
4.9. Liver Cancer
4.10. Lung Cancer
4.11. Melanoma
4.12. Oral Cancer
4.13. Ovarian Cancer
4.14. Pancreatic Cancer
4.15. Prostate Cancer
4.16. Thyroid Cancer
5. Biotransformation and Pharmacokinetics of XH
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Stompor, M.; Żarowska, B. Antimicrobial Activity of Xanthohumol and Its Selected Structural Analogues. Molecules 2016, 21, 608. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-L.; Zhang, Y.-D.; Wang, T.; Guo, H.-Y.; Liu, Q.-M.; Su, H.-X. Evaluation on Antioxidant Effect of Xanthohumol by Different Antioxidant Capacity Analytical Methods. J. Chem. 2014, 2014, 249485. [Google Scholar] [CrossRef]
- Gao, X.; Deeb, D.; Liu, Y.; Gautam, S.; Dulchavsky, S.A.; Gautam, S.C. Immunomodulatory activity of xanthohumol: Inhibition of T cell proliferation, cell-mediated cytotoxicity and Th1 cytokine production through suppression of NF-kappaB. Immunopharmacol. Immunotoxicol. 2009, 31, 477–484. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, P.J.; Carvalho, D.O.; Cruz, J.M.; Guido, L.F.; Barros, A.A. Fundamentals and health benefits of xanthohumol, a natural product derived from hops and beer. Nat. Prod. Commun. 2009, 4, 591–610. [Google Scholar] [CrossRef] [Green Version]
- Plazar, J.; Filipic, M.; Groothuis, G.M.M. Antigenotoxic effect of Xanthohumol in rat liver slices. Toxicol. In Vitro 2008, 22, 318–327. [Google Scholar] [CrossRef]
- Dell’Eva, R.; Ambrosini, C.; Vannini, N.; Piaggio, G.; Albini, A.; Ferrari, N. AKT/NF-kappaB inhibitor xanthohumol targets cell growth and angiogenesis in hematologic malignancies. Cancer 2007, 110, 2007–2011. [Google Scholar] [CrossRef] [PubMed]
- Frölich, S.; Schubert, C.; Bienzle, U.; Jenett-Siems, K. In vitro antiplasmodial activity of prenylated chalcone derivatives of hops (Humulus lupulus) and their interaction with haemin. J. Antimicrob. Chemother. 2005, 55, 883–887. [Google Scholar] [CrossRef] [PubMed]
- Goto, K.; Asai, T.; Hara, S.; Namatame, I.; Tomoda, H.; Ikemoto, M.; Oku, N. Enhanced antitumor activity of xanthohumol, a diacylglycerol acyltransferase inhibitor, under hypoxia. Cancer Lett. 2005, 219, 215–222. [Google Scholar] [CrossRef]
- Karabín, M. Biologically Active Compounds from Hops and Prospects for Their Use. Compr. Rev. Food Sci. Food Saf. 2016, 15, 542–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Yin, H.; Liu, G.; Dong, J.; Qian, Z.; Miao, J. Xanthohumol, a prenylated chalcone from beer hops, acts as an α-glucosidase inhibitor in vitro. J. Agric. Food Chem. 2014, 62, 5548–5554. [Google Scholar] [CrossRef]
- Jiang, C.-H.; Sun, T.-L.; Xiang, D.-X.; Wei, S.-S.; Li, W.-Q. Anticancer Activity and Mechanism of Xanthohumol: A Prenylated Flavonoid From Hops (Humulus lupulus L.). Front. Pharmacol. 2018, 9, 530. [Google Scholar] [CrossRef] [PubMed]
- Ambrož, M.; Lněničková, K.; Matoušková, P.; Skálová, L.; Boušová, I. Antiproliferative Effects of Hop-derived Prenylflavonoids and Their Influence on the Efficacy of Oxaliplatine, 5-fluorouracil and Irinotecan in Human ColorectalC Cells. Nutrients 2019, 11, 879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verzele, M.; Stockx, J.; Fontijn, F.; Anteunis, M. Xanthohumol, a new natural chalkone. Bull. Sociétés Chim. Belg. 1957, 66, 452–475. [Google Scholar] [CrossRef]
- Stevens, J.F.; Page, J.E. Xanthohumol and related prenylflavonoids from hops and beer: To your good health! Phytochemistry 2004, 65, 1317–1330. [Google Scholar] [CrossRef] [PubMed]
- Zanoli, P.; Zavatti, M. Pharmacognostic and pharmacological profile of Humulus lupulus L. J. Ethnopharmacol. 2008, 116, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Vázquez Loureiro, P.; Hernández Jiménez, I.; Sendón, R.; Rodriguez-Bernaldo de Quirós, A.; Barbosa-Pereira, L. Determination of Xanthohumol in Hops, Food Supplements and Beers by HPLC. Foods 2019, 8, 435. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Uto-Kondo, H.; Ogura, M.; Ayaori, M.; Shiotani, K.; Ota, A.; Tsuchiya, Y.; Ikewaki, K. Xanthohumol, a hop-derived prenylated flavonoid, promotes macrophage reverse cholesterol transport. J. Nutr. Biochem. 2017, 47, 29–34. [Google Scholar] [CrossRef]
- Krajka-Kuźniak, V.; Cykowiak, M.; Szaefer, H.; Kleszcz, R.; Baer-Dubowska, W. Combination of xanthohumol and phenethyl isothiocyanate inhibits NF-κB and activates Nrf2 in pancreatic cancer cells. Toxicol. In Vitro 2020, 65, 104799. [Google Scholar] [CrossRef]
- Lin, M.; Xiang, D.; Chen, X.; Huo, H. Role of Characteristic Components of Humulus lupulus in Promoting Human Health. J. Agric. Food Chem. 2019, 67, 8291–8302. [Google Scholar] [CrossRef]
- Iniguez, A.B.; Zhu, M.-J. Hop bioactive compounds in prevention of nutrition-related noncommunicable diseases. Crit. Rev. Food Sci. Nutr. 2020, 1–14. [Google Scholar] [CrossRef]
- Gerhäuser, C. Beer constituents as potential cancer chemopreventive agents. Eur. J. Cancer 2005, 41, 1941–1954. [Google Scholar] [CrossRef]
- Nookandeh, A.; Frank, N.; Steiner, F.; Ellinger, R.; Schneider, B.; Gerhäuser, C.; Becker, H. Xanthohumol metabolites in faeces of rats. Phytochemistry 2004, 65, 561–570. [Google Scholar] [CrossRef]
- Pang, Y.; Nikolic, D.; Zhu, D.; Chadwick, L.R.; Pauli, G.F.; Farnsworth, N.R.; van Breemen, R.B. Binding of the hop (Humulus lupulus L.) chalcone xanthohumol to cytosolic proteins in Caco-2 intestinal epithelial cells. Mol. Nutr. Food Res. 2007, 51, 872–879. [Google Scholar] [CrossRef]
- Venturelli, S.; Burkard, M.; Biendl, M.; Lauer, U.M.; Frank, J.; Busch, C. Prenylated chalcones and flavonoids for the prevention and treatment of cancer. Nutrition 2016, 32, 1171–1178. [Google Scholar] [CrossRef]
- Small, E. The relationships of hop cultivars and wild variants of Humulus lupulus. Can. J. Bot. 1980, 58, 676–686. [Google Scholar] [CrossRef]
- Royle, D.J. Hops. By RA Neve. London: Chapman and Hall, (1991), pp. 266, £32.50, ISBN 0-412-30330-2. Exp. Agric. 1992, 28, 123–124. [Google Scholar] [CrossRef]
- Murakami, A.; Darby, P.; Javornik, B.; Pais, M.S.S.; Seigner, E.; Lutz, A.; Svoboda, P. Microsatellite DNA analysis of wild hops, Humulus lupulus L. Genet. Resour. Crop Evol. 2006, 53, 1553–1562. [Google Scholar] [CrossRef]
- Olsovska, J.; Bostikova, V.; Dusek, M.; Jandovska, V.; Bogdanova, K.; Cermak, P.; Bostik, P.; Mikyska, A.; Kolar, M. Humulus lupulus L.(hops)–a valuable source of compounds with bioactive effects for future therapies. Mil. Med. Sci. Lett. 2016, 85, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Moir, M. Hops—A millennium review. J. Am. Soc. Brew. Chem. 2000, 58, 131–146. [Google Scholar] [CrossRef]
- Edwardson, J.R. Hops—Their botany, history, production and utilization. Econ. Bot. 1952, 6, 160–175. [Google Scholar] [CrossRef]
- The Angiosperm Phylogeny Group. An update of the Angiosperm Phylogeny Group classification for the orders and families of flowering plants: APG II. Bot. J. Linn. Soc. 2003, 141, 399–436. [Google Scholar] [CrossRef] [Green Version]
- Stevens, R. The chemistry of hop constituents. Chem. Rev. 1967, 67, 19–71. [Google Scholar] [CrossRef]
- Chapman, A.C. The Hop and Its Constituents: A Monograph on the Hop Plant; The Brewing Trade Review: London, UK, 1905. [Google Scholar]
- Small, E. A numerical and nomenclatural analysis of morpho-geographic taxa of Humulus. Syst. Bot. 1978, 3, 37–76. [Google Scholar] [CrossRef]
- Robert, T.R.; Wilson, R.J.H. Hops. In Handbook of Brewing; Taylor & Francis Group: Boca Raton, FL, USA, 2006. [Google Scholar]
- Šrédl, K.; Prášilová, M.; Svoboda, R.; Severová, L. Hop production in the Czech Republic and its international aspects. Heliyon 2020, 6, e04371. [Google Scholar] [CrossRef]
- Brissart, R.; Brauninger, U.; Haydon, S.; Morand, R.; Palmer, G.H.O.; Sauvage, R.; Seward, B. Malting Technology: Manual of Good Practice; Frachverlag Hans Carl: Wolverhampton, UK, 2000; ISBN 3418007538. [Google Scholar]
- Goldhammer, T. The Brewers Handbook: The Complete Book to Brewing Beer; Apex: Centreville, VA, USA, 2008; ISBN 0967521238. [Google Scholar]
- De Keukeleire, D. Fundamentals of beer and hop chemistry. Quim. Nova 2000, 23, 108–112. [Google Scholar] [CrossRef]
- Wunderlich, S.; Zürcher, A.; Back, W. Enrichment of xanthohumol in the brewing process. Mol. Nutr. Food Res. 2005, 49, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Zamzow, D.R.; Elias, V.; Legette, L.L.; Choi, J.; Stevens, J.F.; Magnusson, K.R. Xanthohumol improved cognitive flexibility in young mice. Behav. Brain Res. 2014, 275, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Nuti, E.; Bassani, B.; Camodeca, C.; Rosalia, L.; Cantelmo, A.; Gallo, C.; Baci, D.; Bruno, A.; Orlandini, E.; Nencetti, S.; et al. Synthesis and antiangiogenic activity study of new hop chalcone Xanthohumol analogues. Eur. J. Med. Chem. 2017, 138, 890–899. [Google Scholar] [CrossRef]
- Teng, Y.; Li, X.; Yang, K.; Li, X.; Zhang, Z.; Wang, L.; Deng, Z.; Song, B.; Yan, Z.; Zhang, Y.; et al. Synthesis and antioxidant evaluation of desmethylxanthohumol analogs and their dimers. Eur. J. Med. Chem. 2017, 125, 335–345. [Google Scholar] [CrossRef]
- Guo, H.; Wu, F.; Wang, Y. Overexpressed ubiquitin ligase Cullin7 in breast cancer promotes cell proliferation and invasion via down-regulating p53. Biochem. Biophys. Res. Commun. 2014, 450, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Keyaerts, M.; Xavier, C.; Heemskerk, J.; Devoogdt, N.; Everaert, H.; Ackaert, C.; Vanhoeij, M.; Duhoux, F.P.; Gevaert, T.; Simon, P. Phase I study of 68Ga-HER2-nanobody for PET/CT assessment of HER2 expression in breast carcinoma. J. Nucl. Med. 2016, 57, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Al-Hussaini, H.; Subramanyam, D.; Reedijk, M.; Sridhar, S.S. Notch signaling pathway as a therapeutic target in breast cancer. Mol. Cancer Ther. 2011, 10, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, X.; Zhang, M.; Wu, H.; Xu, H.; Han, N.; Chu, Q.; Yu, S.; Chen, Y.; Wu, K. Expression of Notch1 correlates with breast cancer progression and prognosis. PLoS ONE 2015, 10, e0131689. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Pear, W.S.; Aster, J.C. The multifaceted role of Notch in cancer. Curr. Opin. Genet. Dev. 2007, 17, 52–59. [Google Scholar] [CrossRef]
- Robinson, D.R.; Kalyana-Sundaram, S.; Wu, Y.-M.; Shankar, S.; Cao, X.; Ateeq, B.; Asangani, I.A.; Iyer, M.; Maher, C.A.; Grasso, C.S. Functionally recurrent rearrangements of the MAST kinase and Notch gene families in breast cancer. Nat. Med. 2011, 17, 1646. [Google Scholar] [CrossRef] [Green Version]
- Guerreiro, S.; Monteiro, R.; Martins, M.J.; Calhau, C.; Azevedo, I.; Soares, R. Distinct modulation of alkaline phosphatase isoenzymes by 17beta-estradiol and xanthohumol in breast cancer MCF-7 cells. Clin. Biochem. 2007, 40, 268–273. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Zhou, C.; Liu, F.; Zhang, W.; Chen, J.; Pan, Y.; Ma, L.; Liu, Q.; Du, Y.; Yang, J.; et al. Inhibition of breast cancer cell survival by Xanthohumol via modulation of the Notch signaling pathway in vivo and in vitro. Oncol. Lett. 2018, 15, 908–916. [Google Scholar] [CrossRef] [Green Version]
- Yin, L.; Sun, Z.; Ren, Q.; Su, X.; Zhang, D. Methyl eugenol induces potent anticancer effects in RB355 human retinoblastoma cells by inducing autophagy, cell cycle arrest and inhibition of PI3K/mTOR/Akt signalling pathway. J. BUON 2018, 23, 1174–1178. [Google Scholar] [PubMed]
- Monteiro, R.; Calhau, C.; Silva, A.O.E.; Pinheiro-Silva, S.; Guerreiro, S.; Gärtner, F.; Azevedo, I.; Soares, R. Xanthohumol inhibits inflammatory factor production and angiogenesis in breast cancer xenografts. J. Cell. Biochem. 2008, 104, 1699–1707. [Google Scholar] [CrossRef] [PubMed]
- Jitariu, A.-A.; Cîmpean, A.M.; Ribatti, D.; Raica, M. Triple negative breast cancer: The kiss of death. Oncotarget 2017, 8, 46652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassey-Archibong, B.I.; Kwiecien, J.M.; Milosavljevic, S.B.; Hallett, R.M.; Rayner, L.G.A.; Erb, M.J.; Crawford-Brown, C.J.; Stephenson, K.B.; Bédard, P.A.; Hassell, J.A. Kaiso depletion attenuates transforming growth factor-β signaling and metastatic activity of triple-negative breast cancer cells. Oncogenesis 2016, 5, e208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, Y.B.; Park, K.S.; Kim, J.B.; Kang, H.J.; Yang, J.H.; Lee, E.K.; Kim, H.Y. Xanthohumol inhibits cellular proliferation in a breast cancer cell line (MDA-MB231) through an intrinsic mitochondrial-dependent pathway. Indian J. Cancer 2014, 51, 518–523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.Y.; Lee, I.-S.; Moon, A. 2-Hydroxychalcone and xanthohumol inhibit invasion of triple negative breast cancer cells. Chem. Biol. Interact. 2013, 203, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Fujisaki, M.; Ishii, S.; Ikeda, T.; Kitajima, M. Evaluation of bone metabolic markers in breast cancer with bone metastasis. Breast Cancer 2001, 8, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Park, M.A.; Heo, S.-W.; Park, S.-Y.; Kang, K.W.; Park, P.-H.; Kim, J.-A. The radio-sensitizing effect of xanthohumol is mediated by STAT3 and EGFR suppression in doxorubicin-resistant MCF-7 human breast cancer cells. Biochim. Biophys. Acta 2013, 1830, 2638–2648. [Google Scholar] [CrossRef] [PubMed]
- Marquina, G.; Manzano, A.; Casado, A. Targeted agents in cervical cancer: Beyond bevacizumab. Curr. Oncol. Rep. 2018, 20, 40. [Google Scholar] [CrossRef] [PubMed]
- Yong, W.K.; Abd Malek, S.N. Xanthohumol induces growth inhibition and apoptosis in ca ski human cervical cancer cells. Evid. Based Complement. Altern. Med. 2015, 2015, 921306. [Google Scholar] [CrossRef] [PubMed]
- Walden, D.; Kunnimalaiyaan, S.; Sokolowski, K.; Clark, T.G.; Kunnimalaiyaan, M. Antiproliferative and apoptotic effects of xanthohumol in cholangiocarcinoma. Oncotarget 2017, 8, 88069–88078. [Google Scholar] [CrossRef]
- Dokduang, H.; Yongvanit, P.; Namwat, N.; Pairojkul, C.; Sangkhamanon, S.; Yageta, M.S.; Murakami, Y.; Loilome, W. Xanthohumol inhibits STAT3 activation pathway leading to growth suppression and apoptosis induction in human cholangiocarcinoma cells. Oncol. Rep. 2016, 35, 2065–2072. [Google Scholar] [CrossRef] [Green Version]
- Thongchot, S.; Thanee, M.; Loilome, W.; Techasen, A.; Boonmars, T.; Sa-Ngiamwibool, P.; Titapun, A.; Yongvanit, P.; Isidoro, C.; Namwat, N. Curative effect of xanthohumol supplementation during liver fluke-associated cholangiocarcinogenesis: Potential involvement of autophagy. J. Tradit. Complement. Med. 2020, 10, 230–235. [Google Scholar] [CrossRef]
- Pan, L.; Becker, H.; Gerhäuser, C. Xanthohumol induces apoptosis in cultured 40-16 human colon cancer cells by activation of the death receptor- and mitochondrial pathway. Mol. Nutr. Food Res. 2005, 49, 837–843. [Google Scholar] [CrossRef]
- Lee, S.H.; Kim, H.J.; Lee, J.S.; Lee, I.-S.; Kang, B.Y. Inhibition of topoisomerase I activity and efflux drug transporters’ expression by xanthohumol. from hops. Arch. Pharm. Res. 2007, 30, 1435–1439. [Google Scholar] [CrossRef]
- Scagliarini, A.; Mathey, A.; Aires, V.; Delmas, D. Xanthohumol, a Prenylated Flavonoid from Hops, Induces DNA Damages in Colorectal Cancer Cells and Sensitizes SW480 Cells to the SN38 Chemotherapeutic Agent. Cells 2020, 9, 932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, M.; Yin, H.; Qian, X.; Dong, J.; Qian, Z.; Miao, J. Xanthohumol, a Prenylated Chalcone from Hops, Inhibits the Viability and Stemness of Doxorubicin-Resistant MCF-7/ADR Cells. Molecules 2016, 22, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; An, L.J.; Li, Y.; Wang, Y.; Zhao, L.; Lv, X.; Guo, J.; Song, A.L. Xanthohumol chalcone acts as a powerful inhibitor of carcinogenesis in drug-resistant human colon carcinoma and these effects are mediated via G2/M phase cell cycle arrest, activation of apoptotic pathways, caspase activation and targeting Ras /MEK/ERK p. J. BUON 2019, 24, 2442–2447. [Google Scholar]
- Miranda, C.L.; Stevens, J.F.; Helmrich, A.; Henderson, M.C.; Rodriguez, R.J.; Yang, Y.H.; Deinzer, M.L.; Barnes, D.W.; Buhler, D.R. Antiproliferative and cytotoxic effects of prenylated flavonoids from hops (Humulus lupulus) in human cancer cell lines. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 1999, 37, 271–285. [Google Scholar] [CrossRef]
- Liu, W.; Li, W.; Liu, H.; Yu, X. Xanthohumol inhibits colorectal cancer cells via downregulation of Hexokinases II-mediated glycolysis. Int. J. Biol. Sci. 2019, 15, 2497–2508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, S.; Song, M.; Zhao, R.; Liu, X.; Kang, W.K.; Lee, J.M.; Kim, Y.E.; Zhang, C.; Shim, J.-H.; Liu, K.; et al. Xanthohumol Inhibits the Growth of Keratin 18-Overexpressed Esophageal Squamous Cell Carcinoma in vitro and in vivo. Front. Cell Dev. Biol. 2020, 8, 366. [Google Scholar] [CrossRef]
- Chen, P.-H.; Chang, C.-K.; Shih, C.-M.; Cheng, C.-H.; Lin, C.-W.; Lee, C.-C.; Liu, A.-J.; Ho, K.-H.; Chen, K.-C. The miR-204-3p-targeted IGFBP2 pathway is involved in xanthohumol-induced glioma cell apoptotic death. Neuropharmacology 2016, 110, 362–375. [Google Scholar] [CrossRef]
- Festa, M.; Capasso, A.; D’Acunto, C.W.; Masullo, M.; Rossi, A.G.; Pizza, C.; Piacente, S. Xanthohumol induces apoptosis in human malignant glioblastoma cells by increasing reactive oxygen species and activating MAPK pathways. J. Nat. Prod. 2011, 74, 2505–2513. [Google Scholar] [CrossRef]
- Yuan, J.; Peng, G.; Xiao, G.; Yang, Z.; Huang, J.; Liu, Q.; Yang, Z.; Liu, D. Xanthohumol suppresses glioblastoma via modulation of Hexokinase 2 -mediated glycolysis. J. Cancer 2020, 11, 4047–4058. [Google Scholar] [CrossRef]
- Benelli, R. The AKT/NF-κB inhibitor xanthohumol is a potent anti-lymphocytic leukemia drug overcoming chemoresistance and cell infiltration. Biochem. Pharmacol. 2012, 83, 1634–1642. [Google Scholar] [CrossRef]
- Harikumar, K.B.; Kunnumakkara, A.B.; Ahn, K.S.; Anand, P.; Krishnan, S.; Guha, S.; Aggarwal, B.B. Modification of the cysteine residues in IkappaBalpha kinase and NF-kappaB (p65) by xanthohumol leads to suppression of NF-kappaB-regulated gene products and potentiation of apoptosis in leukemia cells. Blood 2009, 113, 2003–2013. [Google Scholar] [CrossRef] [Green Version]
- Monteghirfo, S.; Tosetti, F.; Ambrosini, C.; Stigliani, S.; Pozzi, S.; Frassoni, F.; Fassina, G.; Soverini, S.; Albini, A.; Ferrari, N. Antileukemia effects of xanthohumol in Bcr/Abl-transformed cells involve nuclear factor-kappaB and p53 modulation. Mol. Cancer Ther. 2008, 7, 2692–2702. [Google Scholar] [CrossRef] [Green Version]
- Sławińska-Brych, A.; Król, S.K.; Dmoszyńska-Graniczka, M.; Zdzisińska, B.; Stepulak, A.; Gagoś, M. Xanthohumol inhibits cell cycle progression and proliferation of larynx cancer cells in vitro. Chem. Biol. Interact. 2015, 240, 110–118. [Google Scholar] [CrossRef]
- Li, Y.; Wang, K.; Yin, S.; Zheng, H.; Min, D. Xanthohumol inhibits proliferation of laryngeal squamous cell carcinoma. Oncol. Lett. 2016, 12, 5289–5294. [Google Scholar] [CrossRef] [Green Version]
- Plazar, J.; Zegura, B.; Lah, T.T.; Filipic, M. Protective effects of xanthohumol against the genotoxicity of benzo(a)pyrene (BaP), 2-amino-3-methylimidazo[4,5-f]quinoline (IQ) and tert-butyl hydroperoxide (t-BOOH) in HepG2 human hepatoma cells. Mutat. Res. 2007, 632, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kunnimalaiyaan, S.; Sokolowski, K.M.; Balamurugan, M.; Gamblin, T.C.; Kunnimalaiyaan, M. Xanthohumol inhibits Notch signaling and induces apoptosis in hepatocellular carcinoma. PLoS ONE 2015, 10, e0127464. [Google Scholar] [CrossRef] [PubMed]
- Ho, Y.-C.; Liu, C.-H.; Chen, C.-N.; Duan, K.-J.; Lin, M.-T. Inhibitory effects of xanthohumol from hops (Humulus lupulus L.) on human hepatocellular carcinoma cell lines. Phytother. Res. 2008, 22, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Koo, J.-H.; Kim, H.T.; Yoon, H.-Y.; Kwon, K.-B.; Choi, I.-W.; Jung, S.H.; Kim, H.-U.; Park, B.-H.; Park, J.-W. Effect of xanthohumol on melanogenesis in B16 melanoma cells. Exp. Mol. Med. 2008, 40, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Drenzek, J.G.; Seiler, N.L.; Jaskula-Sztul, R.; Rausch, M.M.; Rose, S.L. Xanthohumol decreases Notch1 expression and cell growth by cell cycle arrest and induction of apoptosis in epithelial ovarian cancer cell lines. Gynecol. Oncol. 2011, 122, 396–401. [Google Scholar] [CrossRef]
- Li, M.; Gao, F.; Yu, X.; Zhao, Q.; Zhou, L.; Liu, W.; Li, W. Promotion of ubiquitination-dependent survivin destruction contributes to xanthohumol-mediated tumor suppression and overcomes radioresistance in human oral squamous cell carcinoma. J. Exp. Clin. Cancer Res. 2020, 39, 88. [Google Scholar] [CrossRef]
- Jiang, W.; Zhao, S.; Xu, L.; Lu, Y.; Lu, Z.; Chen, C.; Ni, J.; Wan, R.; Yang, L. The inhibitory effects of xanthohumol, a prenylated chalcone derived from hops, on cell growth and tumorigenesis in human pancreatic cancer. Biomed. Pharmacother. 2015, 73, 40–47. [Google Scholar] [CrossRef]
- Saito, K.; Matsuo, Y.; Imafuji, H.; Okubo, T.; Maeda, Y.; Sato, T.; Shamoto, T.; Tsuboi, K.; Morimoto, M.; Takahashi, H.; et al. Xanthohumol inhibits angiogenesis by suppressing nuclear factor-κB activation in pancreatic cancer. Cancer Sci. 2018, 109, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colgate, E.C.; Miranda, C.L.; Stevens, J.F.; Bray, T.M.; Ho, E. Xanthohumol, a prenylflavonoid derived from hops induces apoptosis and inhibits NF-kappaB activation in prostate epithelial cells. Cancer Lett. 2007, 246, 201–209. [Google Scholar] [CrossRef] [PubMed]
- Deeb, D.; Gao, X.; Jiang, H.; Arbab, A.S.; Dulchavsky, S.A.; Gautam, S.C. Growth inhibitory and apoptosis-inducing effects of xanthohumol, a prenylated chalone present in hops, in human prostate cancer cells. Anticancer Res. 2010, 30, 3333–3339. [Google Scholar] [PubMed]
- Venè, R.; Benelli, R.; Minghelli, S.; Astigiano, S.; Tosetti, F.; Ferrari, N. Xanthohumol impairs human prostate cancer cell growth and invasion and diminishes the incidence and progression of advanced tumors in TRAMP mice. Mol. Med. 2012, 18, 1292–1302. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.R.; Luo, J.; Ndiaye, M.; Chen, H.; Kunnimalaiyaan, M. Xanthohumol inhibits the neuroendocrine transcription factor achaete-scute complex-like 1, suppresses proliferation, and induces phosphorylated ERK1/2 in medullary thyroid cancer. Am. J. Surg. 2010, 199, 315–318; discussion 318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; Zhang, L.; Li, G.; Gao, Z. Xanthohumol protects against Azoxymethane-induced colorectal cancer in Sprague-Dawley rats. Environ. Toxicol. 2020, 35, 136–144. [Google Scholar] [CrossRef]
- Gao, F.; Li, M.; Zhou, L.; Liu, W.; Zuo, H.; Li, W. Xanthohumol targets the ERK1/2‑Fra1 signaling axis to reduce cyclin D1 expression and inhibit non‑small cell lung cancer. Oncol. Rep. 2020, 44, 1365–1374. [Google Scholar] [CrossRef]
- Liu, X.; Song, M.; Wang, P.; Zhao, R.; Chen, H.; Zhang, M.; Shi, Y.; Liu, K.; Liu, F.; Yang, R.; et al. Targeted therapy of the AKT kinase inhibits esophageal squamous cell carcinoma growth in vitro and in vivo. Int. J. Cancer 2019, 145, 1007–1019. [Google Scholar] [CrossRef] [Green Version]
- Reitman, Z.J.; Winkler, F.; Elia, A.E.H. New Directions in the Treatment of Glioblastoma. Semin. Neurol. 2018, 38, 50–61. [Google Scholar] [CrossRef] [Green Version]
- Festa, M.; Caputo, M.; Cipolla, C.; D’Acunto, C.; Rossi, A.; Tecce, M.; Capasso, A. The involvement of xanthohumol in the expression of annexin in human malignant glioblastoma cells. Open Biochem. J. 2013, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Cosetti, M.; Yu, G.-P.; Schantz, S.P. Five-year survival rates and time trends of laryngeal cancer in the US population. Arch. Otolaryngol. Head Neck Surg. 2008, 134, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Weiskirchen, R.; Mahli, A.; Weiskirchen, S.; Hellerbrand, C. The hop constituent xanthohumol exhibits hepatoprotective effects and inhibits the activation of hepatic stellate cells at different levels. Front. Physiol. 2015, 6, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, I.E.; Miranda, C.L.; Lowry, M.B.; Maier, C.S.; Stevens, J.F.; Gombart, A.F. Antiproliferative and Cytotoxic Activity of Xanthohumol and Its Non-Estrogenic Derivatives in Colon and Hepatocellular Carcinoma Cell Lines. Int. J. Mol. Sci. 2019, 20, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorn, C.; Weiss, T.S.; Heilmann, J.; Hellerbrand, C. Xanthohumol, a prenylated chalcone derived from hops, inhibits proliferation, migration and interleukin-8 expression of hepatocellular carcinoma cells. Int. J. Oncol. 2010, 36, 435–441. [Google Scholar] [CrossRef] [Green Version]
- Krajka-Kuźniak, V.; Paluszczak, J.; Baer-Dubowska, W. Xanthohumol induces phase II enzymes via Nrf2 in human hepatocytes in vitro. Toxicol. In Vitro 2013, 27, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Ferk, F.; Huber, W.W.; Filipic, M.; Bichler, J.; Haslinger, E.; Misík, M.; Nersesyan, A.; Grasl-Kraupp, B.; Zegura, B.; Knasmüller, S. Xanthohumol, a prenylated flavonoid contained in beer, prevents the induction of preneoplastic lesions and DNA damage in liver and colon induced by the heterocyclic aromatic amine amino-3-methyl-imidazo[4,5-f]quinoline (IQ). Mutat. Res. 2010, 691, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Jiang, K.; Liang, B.; Huang, X. Anticancer effect of xanthohumol induces growth inhibition and apoptosis of human liver cancer through NF-κB/p53-apoptosis signaling pathway. Oncol. Rep. 2016, 35, 669–675. [Google Scholar] [CrossRef] [Green Version]
- Sławińska-Brych, A.; Zdzisińska, B.; Dmoszyńska-Graniczka, M.; Jeleniewicz, W.; Kurzepa, J.; Gagoś, M.; Stepulak, A. Xanthohumol inhibits the extracellular signal regulated kinase (ERK) signalling pathway and suppresses cell growth of lung adenocarcinoma cells. Toxicology 2016, 357–358, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Seitz, T.; Hackl, C.; Freese, K.; Dietrich, P.; Mahli, A.; Thasler, R.M.; Thasler, W.E.; Lang, S.A.; Bosserhoff, A.K.; Hellerbrand, C. Xanthohumol, a Prenylated Chalcone Derived from Hops, Inhibits Growth and Metastasis of Melanoma Cells. Cancers 2021, 13, 511. [Google Scholar] [CrossRef] [PubMed]
- Goenka, S.; Simon, S.R. Depigmenting effect of Xanthohumol from hop extract in MNT-1 human melanoma cells and normal human melanocytes. Biochem. Biophys. Rep. 2021, 26, 100955. [Google Scholar] [CrossRef] [PubMed]
- Musella, A.; Bardhi, E.; Marchetti, C.; Vertechy, L.; Santangelo, G.; Sassu, C.; Tomao, F.; Rech, F.; D’Amelio, R.; Monti, M. Rucaparib: An emerging parp inhibitor for treatment of recurrent ovarian cancer. Cancer Treat. Rev. 2018, 66, 7–14. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer Statistics, 2017. CA. Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [Green Version]
- LeComte, M.D.; Spees, J.L. Notch1-STAT3-ETBR signaling in brain injury and cancer. Cytokine 2016, 80, 64–65. [Google Scholar] [CrossRef] [Green Version]
- Dinesh, T.A.; Nair, P.; Abhijath, V.; Jha, V.; Aarthy, K. Economics of cancer care: A community-based cross-sectional study in Kerala, India. South Asian J. Cancer 2020, 9, 7. [Google Scholar]
- Steinestel, J.; Luedeke, M.; Arndt, A.; Schnoeller, T.J.; Lennerz, J.K.; Wurm, C.; Maier, C.; Cronauer, M.V.; Steinestel, K.; Schrader, A.J. Detecting predictive androgen receptor modifications in circulating prostate cancer cells. Oncotarget 2019, 10, 4213. [Google Scholar] [CrossRef] [Green Version]
- Kłósek, M.; Mertas, A.; Król, W.; Jaworska, D.; Szymszal, J.; Szliszka, E. Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand-Induced Apoptosis in Prostate Cancer Cells after Treatment with Xanthohumol-A Natural Compound Present in Humulus lupulus L. Int. J. Mol. Sci. 2016, 17, 837. [Google Scholar] [CrossRef] [Green Version]
- Legette, L.; Karnpracha, C.; Reed, R.L.; Choi, J.; Bobe, G.; Christensen, J.M.; Rodriguez-Proteau, R.; Purnell, J.Q.; Stevens, J.F. Human pharmacokinetics of xanthohumol, an antihyperglycemic flavonoid from hops. Mol. Nutr. Food Res. 2014, 58, 248–255. [Google Scholar] [CrossRef]
- Carvalho, D.O.; Freitas, J.; Nogueira, P.; Henriques, S.N.; Carmo, A.M.; Castro, M.A.; Guido, L.F. Xanthohumol inhibits cell proliferation and induces apoptosis in human thyroid cells. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2018, 121, 450–457. [Google Scholar] [CrossRef]
- Radović, B.; Schmutzler, C.; Köhrle, J. Xanthohumol stimulates iodide uptake in rat thyroid-derived FRTL-5 cells. Mol. Nutr. Food Res. 2005, 49, 832–836. [Google Scholar] [CrossRef]
- Yilmazer, M.; Stevens, J.F.; Deinzer, M.L.; Buhler, D.R. In vitro biotransformation of xanthohumol, a flavonoid from hops (Humulus lupulus), by rat liver microsomes. Drug Metab. Dispos. 2001, 29, 223–231. [Google Scholar]
- Possemiers, S.; Heyerick, A.; Robbens, V.; De Keukeleire, D.; Verstraete, W. Activation of proestrogens from hops (Humulus lupulus L.) by intestinal microbiota; conversion of isoxanthohumol into 8-prenylnaringenin. J. Agric. Food Chem. 2005, 53, 6281–6288. [Google Scholar] [CrossRef] [Green Version]
- Hanske, L.; Loh, G.; Sczesny, S.; Blaut, M.; Braune, A. Recovery and metabolism of xanthohumol in germ-free and human microbiota-associated rats. Mol. Nutr. Food Res. 2010, 54, 1405–1413. [Google Scholar] [CrossRef] [PubMed]
- Jirásko, R.; Holcapek, M.; Vrublová, E.; Ulrichová, J.; Simánek, V. Identification of new phase II metabolites of xanthohumol in rat in vivo biotransformation of hop extracts using high-performance liquid chromatography electrospray ionization tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 4100–4108. [Google Scholar] [CrossRef] [PubMed]
- Bartmanska, A.; Tronina, T.; Poplonski, J.; Huszcza, E. Biotransformations of prenylated hop flavonoids for drug discovery and production. Curr. Drug Metab. 2013, 14, 1083–1097. [Google Scholar] [CrossRef] [PubMed]
- Van Breemen, R.B.; Yuan, Y.; Banuvar, S.; Shulman, L.P.; Qiu, X.; Alvarenga, R.F.R.; Chen, S.-N.; Dietz, B.M.; Bolton, J.L.; Pauli, G.F.; et al. Pharmacokinetics of prenylated hop phenols in women following oral administration of a standardized extract of hops. Mol. Nutr. Food Res. 2014, 58, 1962–1969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolff, H.; Motyl, M.; Hellerbrand, C.; Heilmann, J.; Kraus, B. Xanthohumol uptake and intracellular kinetics in hepatocytes, hepatic stellate cells, and intestinal cells. J. Agric. Food Chem. 2011, 59, 12893–12901. [Google Scholar] [CrossRef]
- Wesołowska, O.; Gąsiorowska, J.; Petrus, J.; Czarnik-Matusewicz, B.; Michalak, K. Interaction of prenylated chalcones and flavanones from common hop with phosphatidylcholine model membranes. Biochim. Biophys. Acta 2014, 1838, 173–184. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Country | Region |
|---|---|
| Germany | Hallertau region |
| USA | Washington, Oregon, and Idaho |
| Other countries | Poland, Czech Republic, South Africa, England, Slovenia, Ukraine, China, Australia, and New Zealand |
| Cancer Type | Cell Line | Effects | Mechanisms | Conc. | Reference |
|---|---|---|---|---|---|
| Breast cancer | Hs57BT and MDA-MB-231 | Decreased cell viability, cell invasion and proliferation | None | 4.78–6.7 µM | [57] |
| MDA-MB-231 | Decreased cell viability | ↑Caspase-3; ↑caspase-9; ↓Bax | 10 and 20 µM | [56] | |
| MCF-7 | Decreased proliferation | ↓ALP isoenzymes | 10 µM | [50] | |
| Adriamycin-resistant MCF-7 | Decreased cell viability, stemness, and increased radio- and chemosensitivity | ↑Apoptosis; ↑γ-H2AX; ↓STAT3; ↓MDR1; ↓EGFR | 10 µM | [59,68] | |
| Cervical cancer | Ca Ski | Decreased proliferation | ↑Apoptosis; ↑caspase-3; ↑caspase-8; ↑caspase-9; ↑cell cycle arrest; ↑p53; ↓XIAP | 59.96 µM | [61] |
| Choliangiocarcinoma | KKU-M139 and KKU-M214 | Decreased cell growth | ↓STAT3 | 20 and 50 µM | [63] |
| Colon cancer | 40-16 colon cancer | Decreased proliferation | None | 4.1, 3.6 and 2.6 µM | [65] |
| HT-29 and CDD-18Co | Decreased cell viability | ↑Apoptosis; ↑caspase-3; ↑caspase-9; ↓cyclin B1; ↓MEK/ERK; ↓G2/M phase of cell cycle | 10 and 100 µM | [69] | |
| HT-29 | Decreased cell viability | None | 48 and 72 µM | [70] | |
| HCT115 | Decreased proliferation | ↓ABCC 1,2,3; ↓ABCB1 | 10.2 µM | [66] | |
| Colorectal cancer | FHC, CCD841, CoN, HT29, SW480, LOVO, HCT116 and SW620 | Decreased cell proliferation, cell viability, and colony formation | ↑Apoptosis; ↓HK2; ↓glycolysis; ↓EGFR-Akt | 25 µM | [71] |
| Esophageal cancer | KYSE30, KYSE70, KYSE410, and KYSE450 | Suppressed proliferation, foci formation, and anchorage-independent colony growth | ↓Apoptosis; ↑cell cycle arrest (G1 phase); ↓Bax; ↓cyclin D1; ↓cyt. c; ↓cleaved-PARP; ↓Bcl-2; ↓cyclin D3; ↓KRT18 | 0.3, 0.6, 1.25, and 2.5 µM | [72] |
| Glioblastoma | U87 glioblastoma | Decreased cell viability | ↑Apoptosis; ↓IGFBP2/Akt/Bcl‑2; ↑mIR-204-3p; ↑ERK/c-Fos | 25 µM | [73] |
| T98G | Decreased cell viability | ↑Apoptosis; ↑ROS; ↑p-p38; ↓p‑ERK1/2; ↑cleavage of PARP ↓caspase-3; ↓caspase-9 | 20 µM | [74] | |
| LN229, T98G and U87-MG | Inhibited proliferation, viability, and colony formation | ↓Akt-GSK3β-FBW7-c-Myc protein, ↓HK2 protein | 2, 5, and 10 µM | [75] | |
| Hematological cancers | Acute lymphoblastic leukemia L1210 and adriamycin-resistant L1210 | Decreased cell viability, invasion and migration | ↑Apoptosis; ↓Akt; ↓FAK; ↓NF-κB | 2.5, 5, and 10 µM | [76] |
| Chronic myeloid leukemia KBM-5 | Suppressed invasion | ↑Apoptosis; ↓IKK activity; ↓p65 nuclear translocation; ↓IκBα degradation and phosphorylation; ↓TRAF-2; ↓cIAP-1; ↓cIAP2; ↓survivin; ↓XIAP; ↓Bcl-xL | 50 µM | [77] | |
| Bcr-Abl+ myeloid leukemia cells K562 | Decreased adhesion to endothelial cells, cell viability, and invasion | ↑Apoptosis; ↓MMP-2; ↓Bcr-Abl; ↑p21; ↑p53 | 2.5, 5, and 10 µM | [78] | |
| Laryngeal cancer | RK33 and RK45 | Decreased cell viability | ↑Apoptosis; ↑caspase-3; ↑caspase-8; ↑caspase-9; ↑p53; ↑p21; ↓cyclin D1; ↓ERK1/2 | 12.3 and 22.5 µM | [79] |
| SCC4 | Decreased proliferation | ↑Apoptosis; ↑PARP; ↑p53; ↑AIF; ↓Bcl-2; ↓Mcl-1 | 20, 30, and 40 µM | [80] | |
| Liver cancer | HepG2 | None | None | 10 µM | [81] |
| Huh7, Hep3B, SK-Hep1, and HepG2 | Decreased colony forming, cell viability and confluency ability | ↓HES1; ↓Notch1 pathway | 5 µM | [82] | |
| Hep3B and HA22T/VGH | None | None | 108 and 166 µM | [83] | |
| Melanoma | B16 | Decreased IBMX-induced melanogenesis | ↓Tyrosine enzyme activity | 0.5, 1.5, and 10 µM | [84] |
| SK-MEL-2 | Decreased proliferation | ↓DNA topoisomerase 1 | 14.4 µM | [66] | |
| Ovarian cancer | A-2780 | Decreased proliferation | None | 0.52 and 5.2 µM | [70] |
| OVCAR3 and SKOV3 | Decreased proliferation | ↓Notch1 pathway; ↑p21; ↑cell cycle arrest | 10, 20, and 30 µM | [85] | |
| Oral squamous cell carcinoma | OSCC | Decreased cell viability and reversed radioresistance | ↓Survivin; ↑mitochondrial apoptotic signaling; ↓Akt-Wee1-CDK1 | 1–5 µM | [86] |
| Pancreatic cancer | PANC1 and BxPC3 | Decreased proliferation, viability, and colony formation | ↑Apoptosis; ↓p-STAT3 | 5–100 µM | [87] |
| BxPC3, MXPaCa2, and AsPC1 | Inhibited cell proliferation | ↓NF-κB; ↓VGEF ↓IL-8; ↓mRNA | 0.5–25 µmol/L | [88] | |
| Prostate cancer | Hormone-refractory AR−PC3 | Decreased cell viability | ↑Apoptosis; ↓activation of NF-κB | 2.5–20 µM | [89] |
| Hormone-sensitive AR+, hormone-refractory AR− PC3, LNCaP and DU145 | Decreased cell viability | ↑Apoptosis; ↓NF-κB; ↓p65; ↓p-Akt; ↓p-mTOR; ↓survivin; ↓Bcl-2 | 24 and 40 µM | [90] | |
| Hormone-refractory AR− PC3, DU145 PC3, DU145 | Decreased proliferation, invasion, and migration | ↓p-FAK; ↓p-Akt | 2.5, 5, and 10 µM | [91] | |
| Thyroid cancer | MTC (medullary thyroid cancer cells) | Decreased proliferation and malignant phenotype | ↑ERK1/2 phosphorylation | 10, 20, and 30 µM | [92] |
| Cancer Type | Animal Model | Effects | Mechanisms | Dose (Route) | Duration | Reference |
|---|---|---|---|---|---|---|
| Breast cancer | BALB/c mouse tumor model by using 4T1 cell lines | Suppressed tumor growth; decreased tumor weight and size | ↓Survivin; ↑caspase cleavage, ↓Notch-1; ↓Ki-67 | 100 and 200 mg/kg | 14 days | [51] |
| Colorectal cancer | Xenograft mouse model by using FHC, SW620, LOVO, CCD841, SW480, CoN, HT29, and HCT116 | Inhibited tumor cell proliferation | ↑Apoptosis; ↑cyt. c release | 10 mg/kg (i.p.) | Every two days | [71] |
| Male Sprague Dawley rats by using SW480 CRC cells | Inhibited tumor cell proliferation | ↑Apoptosis; ↓wnt/β-catenin signaling ↓Bax; ↓ Bcl-2; ↓caspase-3; ↓iNOS; ↓COX-2 | 5 mg/kg for alternate days | 8 weeks | [93] | |
| Esophageal cancer | Patient-derived xenograft mouse model by using KYSE30 cell lines | Decreased tumor volume and weight | ↑Apoptosis; Ki-67; ↓KRT18 | 40, 80, and 160 mg/kg (p.o.) | 64 days | [72] |
| Glioblastoma | Xenograft mouse model by using LN229, U87MG, and T98G cell lines | Reduced tumor weight | ↓Akt-GSK3β-FBW7-c-Myc protein | 10 mg/kg for every three days (i.p.) | 32 days | [75] |
| Lung cancer | Xenograft mouse model by using HCC827 cells | Suppressed tumor growth | ↓Cyclin D1; ↓ERK1/2-fra1 signaling pathway | 10mg/kg (i.p.) | 32 days | [94] |
| Pancreatic cancer | Subcutaneous xenograft mouse model by using BXPC-3 cells | Inhibited tumor growth and angiogenesis | ↓NF-κB activation ↓tube formation; ↓VGEF; ↓IL8 | 10 mg/kg/week | 5 weeks | [88] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Harish, V.; Haque, E.; Śmiech, M.; Taniguchi, H.; Jamieson, S.; Tewari, D.; Bishayee, A. Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets. Int. J. Mol. Sci. 2021, 22, 4478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094478
Harish V, Haque E, Śmiech M, Taniguchi H, Jamieson S, Tewari D, Bishayee A. Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets. International Journal of Molecular Sciences. 2021; 22(9):4478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094478
Chicago/Turabian StyleHarish, Vancha, Effi Haque, Magdalena Śmiech, Hiroaki Taniguchi, Sarah Jamieson, Devesh Tewari, and Anupam Bishayee. 2021. "Xanthohumol for Human Malignancies: Chemistry, Pharmacokinetics and Molecular Targets" International Journal of Molecular Sciences 22, no. 9: 4478. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094478