
Knockdown of the Ribosomal Protein eL38 in HEK293 Cells Changes the Translational Efficiency of Specific Genes

 , ,
, ,
Abstract
:1. Introduction
2. Results
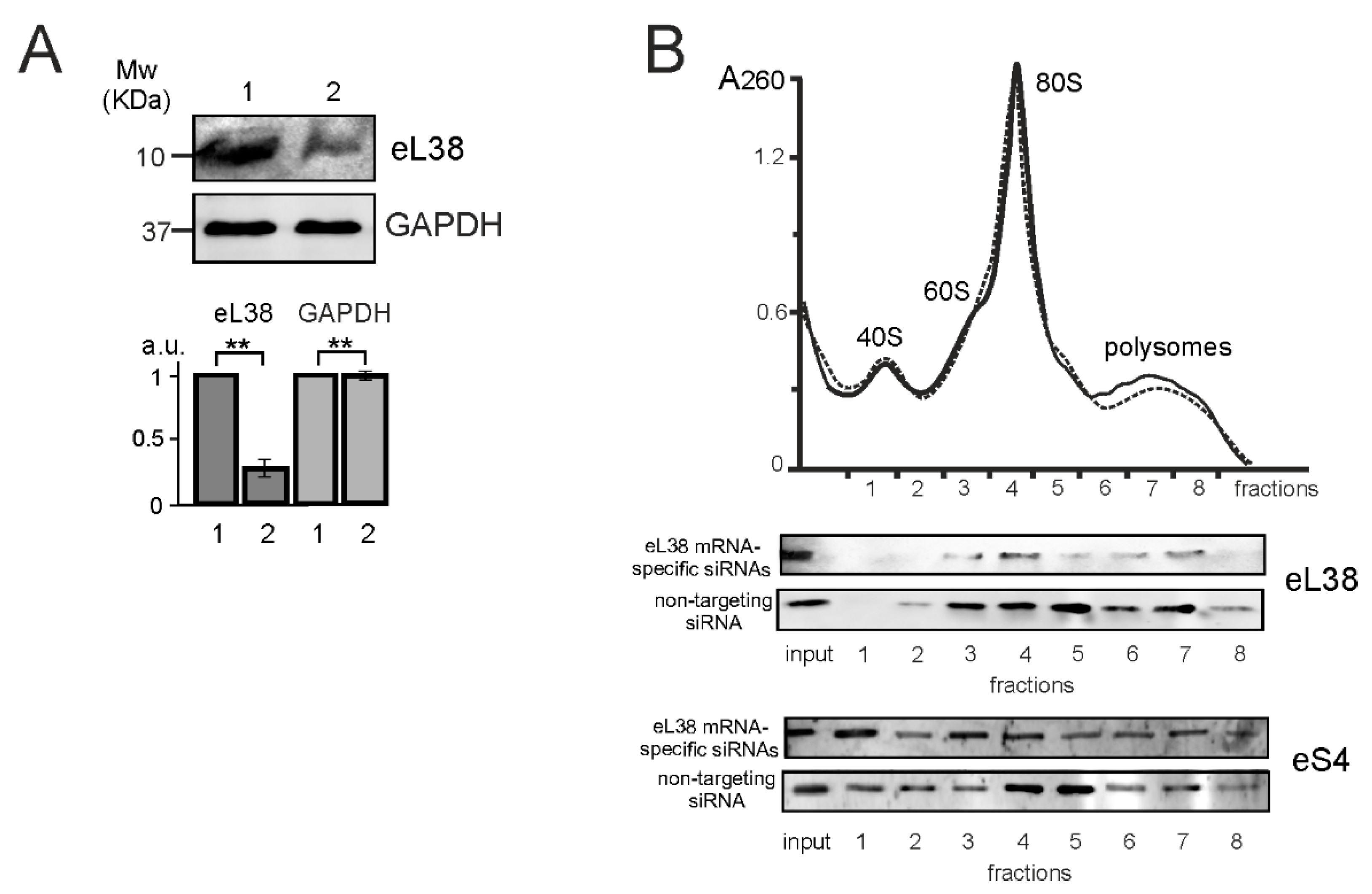
2.1. Ribo-seq of eL38 Knocked down HEK293 Cells
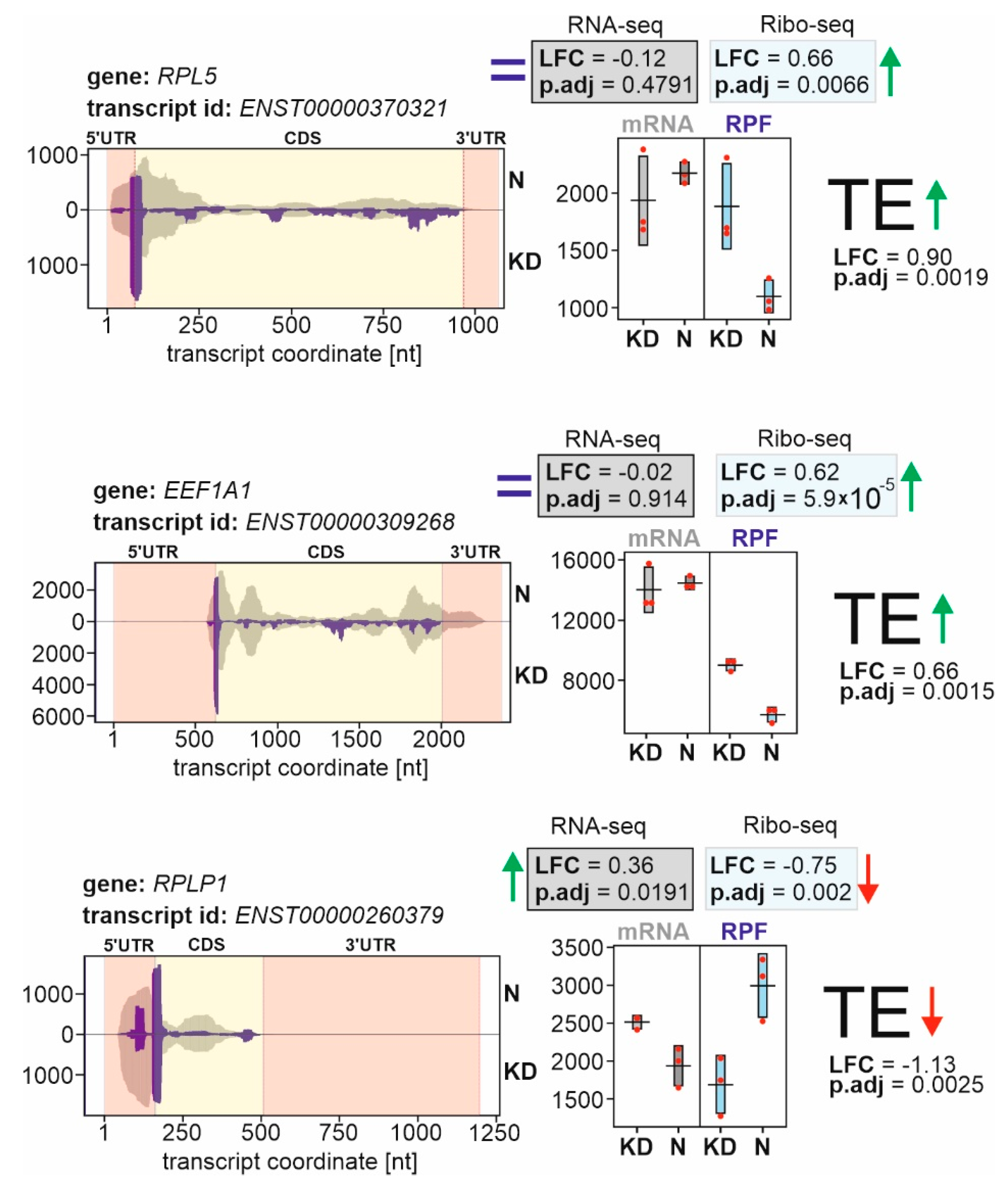
2.2. Identification of Genes with Differential Translational Efficiencies
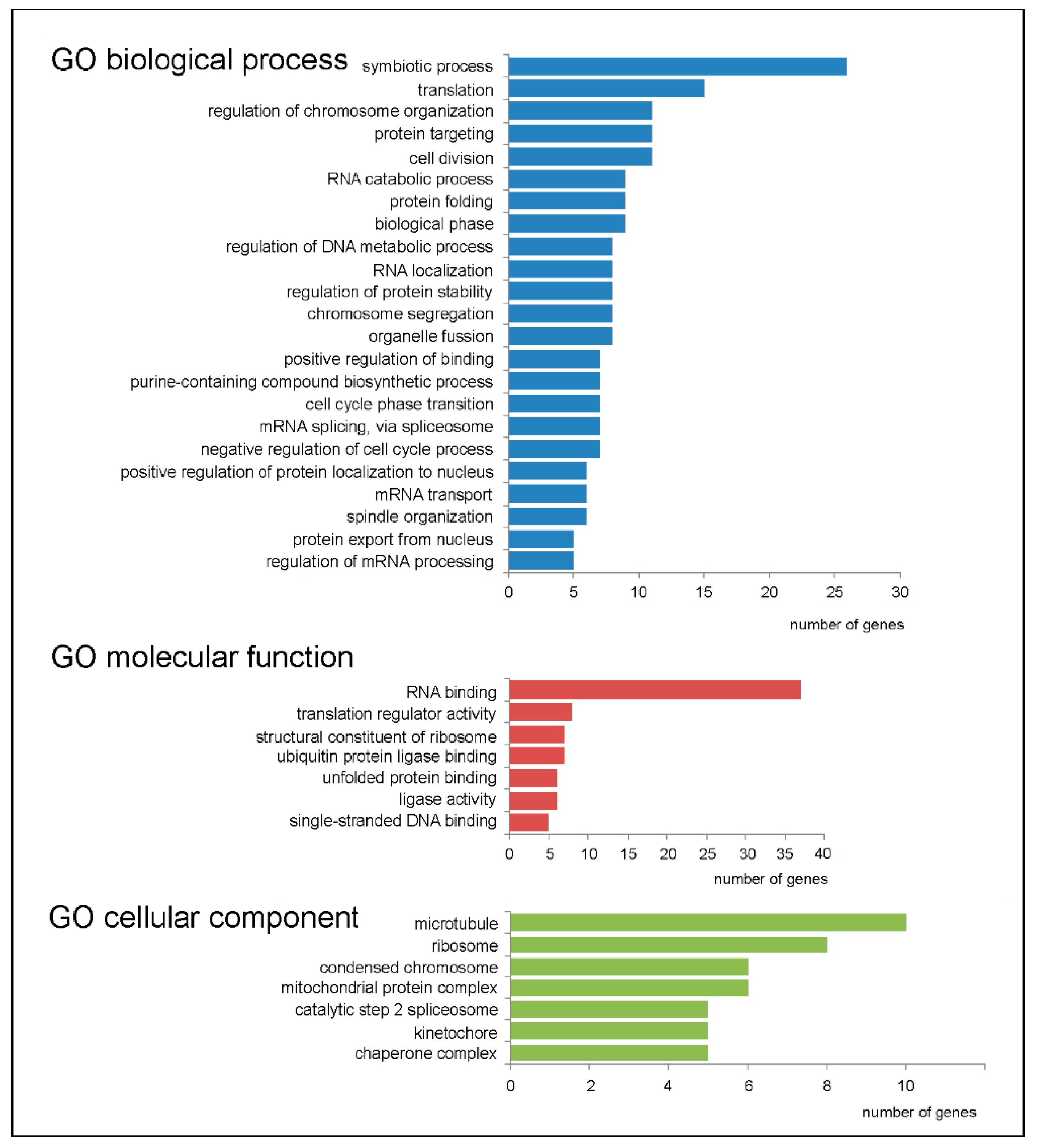
2.3. Genes with Increased TEs and Associated Cellular Processes
2.4. Genes with Decreased TEs and Associated Cellular Processes
3. Discussion
4. Materials and Methods
4.1. Preparation of siRNAs, Cell Culturing, eL38 mRNA Knockdown and Determination of eL38 and Its mRNA Levels
4.2. Ribo-seq Procedures
4.3. NGS Data Analysis
4.4. Determining the GDTEs-Associated Processes
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CDS | coding sequence |
| GDTE | gene with differential translational efficiency |
| GO | Gene Ontology |
| NGS | next generation sequencing |
| TE | translational efficiency |
References
- Anger, A.M.; Armache, J.P.; Berninghausen, O.; Habeck, M.; Subklewe, M.; Wilson, D.N.; Beckmann, R. Structures of the human and Drosophila 80S ribosome. Nature 2013, 497, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Khatter, H.; Myasnikov, A.G.; Natchiar, S.K.; Klaholz, B.P. Structure of the human 80S ribosome. Nature 2015, 520, 640–645. [Google Scholar] [CrossRef] [PubMed]
- Kampen, K.R.; Sulima, S.O.; Vereecke, S.; Keersmaecker, K.D. Hallmarks of ribosomopathies. Nucleic Acids Res. 2020, 48, 1013–1028. [Google Scholar] [CrossRef]
- Slavov, N.; Semrau, S.; Airoldi, E.; Budnik, B.; van Oudenaarden, A. Differential stoichiometry among core ribosomal proteins. Cell Rep. 2015, 13, 865–873. [Google Scholar] [CrossRef] [Green Version]
- Shi, Z.; Fujii, K.; Kovary, K.M.; Genuth, N.R.; Röst, H.L.; Teruel, M.N.; Barna, M. Heterogeneous ribosomes preferentially translate distinct subpools of mRNAs genome-wide. Mol. Cell. 2017, 67, 71–83.e7. [Google Scholar] [CrossRef] [Green Version]
- Xue, S.; Barna, M. Specialized ribosomes: A new frontier in gene regulation and organismal biology. Nat. Rev. Mol. Cell. Biol. 2012, 13, 355–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mauro, V.P.; Edelman, G.M. The ribosome filter hypothesis. Proc. Natl. Acad. Sci. USA 2002, 99, 12031–12036. [Google Scholar] [CrossRef] [Green Version]
- Morgan, W.C. A new tail-short mutation in the mouse whose lethal effects are conditioned by the residual genotypes. J. Hered. 1950, 41, 208–215. [Google Scholar] [CrossRef]
- Kondrashov, N.; Pusic, A.; Stumpf, C.R.; Shimizu, K.; Hsieh, A.C.; Ishijima, J.; Shiroishi, T.; Barna, M. Ribosome-mediated specificity in Hox mRNA translation and vertebrate tissue patterning. Cell 2011, 145, 383–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, S.; Tian, S.; Fujii, K.; Kladwang, W.; Das, R.; Barna, M. RNA regulons in Hox 5′ UTRs confer ribosome specificity to gene regulation. Nature 2015, 517, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Gopanenko, A.V.; Kolobova, A.V.; Meschaninova, M.I.; Venyaminova, A.G.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the mRNA encoding the ribosomal protein eL38 in mammalian cells causes a substantial reorganization of genomic transcription. Biochimie 2021, 184, 132–142. [Google Scholar] [CrossRef]
- Ingolia, N.T.; Ghaemmaghami, S.; Newman, J.R.; Weissman, J.S. Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science 2009, 324, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Piovesan, A.; Caracausi, M.; Antonaros, F.; Pelleri, M.C.; Vitale, L. GeneBase 1.1: A tool to summarize data from NCBI gene datasets and its application to an update of human gene statistics. Database 2016, 2016, baw153. [Google Scholar] [CrossRef] [PubMed]
- Aulehla, A.; Pourquie, O. More than patterning—Hox genes and the control of posterior axial elongation. Dev. Cell. 2009, 17, 439–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellik, D.M. Hox genes and vertebrate axial pattern. Curr. Top. Dev. Biol. 2009, 88, 257–278. [Google Scholar] [CrossRef] [PubMed]
- Mallo, M.; Alonso, C.R. The regulation of Hox gene expression during animal development. Development 2013, 140, 3951–3963. [Google Scholar] [CrossRef] [Green Version]
- Kiniry, S.J.; O’Connor, P.B.F.; Michel, A.M.; Baranov, P.V. Trips-Viz: A transcriptome browser for exploring Ribo-Seq data. Nucleic Acids Res. 2019, 47, D847–D852. [Google Scholar] [CrossRef] [Green Version]
- Zankl, A.; Zabel, B.; Hilbert, K.; Wildhardt, G.; Cuenot, S.; Xavier, B.; Ha-Vinh, R.; Bonafé, L.; Spranger, J.; Superti-Furga, A. Spondyloperipheral dysplasia is caused by truncating mutations in the C-propeptide of COL2A1. Am. J. Med. Genet. A 2004, 129A, 144–148. [Google Scholar] [CrossRef]
- Bogaert, R.; Wilkin, D.; Wilcox, W.R.; Lachman, R.; Rimoin, D.; Cohn, D.H.; Eyre, D.R. Expression, in cartilage, of a 7-amino-acid deletion in type II collagen from two unrelated individuals with Kniest dysplasia. Am. J. Hum. Genet. 1994, 55, 1128–1136. [Google Scholar]
- Eyre, D.R.; Weis, M.A.; Moskowitz, R.W. Cartilage expression of a type II collagen mutation in an inherited form of osteoarthritis associated with a mild chondrodysplasia. J. Clin. Investig. 1991, 87, 357–361. [Google Scholar] [CrossRef] [Green Version]
- Robledo, S.; Idol, R.A.; Crimmins, D.L.; Ladenson, J.H.; Mason, P.J.; Bessler, M. The role of human ribosomal proteins in the maturation of rRNA and ribosome production. RNA 2008, 14, 1918–1929. [Google Scholar] [CrossRef] [Green Version]
- Görlich, D.; Kutay, U. Transport between the cell nucleus and the cytoplasm. Annu. Rev. Cell Dev. Biol. 1999, 15, 607–660. [Google Scholar] [CrossRef]
- Klinge, S.; Voigts-Hoffmann, F.; Leibundgut, M.; Arpagaus, S.; Ban, N. Crystal structure of the eukaryotic 60S ribosomal subunit in complex with initiation factor 6. Science 2011, 334, 941–948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gartmann, M.; Blau, M.; Armache, J.-P.; Mielke, T.; Topf, M.; Beckmann, R. Mechanism of eIF6-mediated inhibition of ribosomal subunit joining. J. Biol. Chem. 2010, 285, 14848–14851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babaylova, E.S.; Gopanenko, A.V.; Bulygin, K.N.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. mRNA regions where 80S ribosomes pause during translation elongation in vivo interact with protein uS19, a component of the decoding site. Nucleic Acids Res. 2020, 48, 912–923. [Google Scholar] [CrossRef] [PubMed]
- Gopanenko, A.V.; Kolobova, A.V.; Meschaninova, M.I.; Venyaminova, A.G.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the ribosomal protein eL29 in mammalian cells leads to significant changes in gene expression at the transcription level. Cells 2020, 9, 1228. [Google Scholar] [CrossRef]
- Gopanenko, A.V.; Malygin, A.A.; Tupikin, A.E.; Laktionov, P.P.; Kabilov, M.R.; Karpova, G.G. Human ribosomal protein eS1 is engaged in cellular events related to processing and functioning of U11 snRNA. Nucleic Acids Res. 2017, 45, 9121–9137. [Google Scholar] [CrossRef] [Green Version]
- Backman, T.W.H.; Girke, T. bioassayR: Cross-target analysis of small molecule bioactivity. J. Chem. Inf. Model. 2016, 56, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Gene Ontology Consortium. The Gene Ontology resource: Enriching a GOld mine. Nucleic Acids Res. 2021, 49, D325–D334. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor package for reactome pathway analysis and visualization. Mol. Biosyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Gene Symbol | Description | p.adj | Shrunken LFC |
|---|---|---|---|---|
| 1 | ATP5MG | ATP synthase membrane subunit g | 0.012694951 | 4.312883344 |
| 2 | TPM4 | tropomyosin 4 | 0.012694951 | 2.407078538 |
| 3 | TUBA1A | tubulin alpha 1a | 0.028524939 | 2.303007559 |
| 4 | NME1 | NME/NM23 nucleoside diphosphate kinase 1 | 0.010574469 | 2.189657665 |
| 5 | CADM1 | cell adhesion molecule 1 | 0.007530612 | 2.027829748 |
| 6 | LAMP2 | lysosomal associated membrane protein 2 | 0.00925901 | 1.908361642 |
| 7 | CDK1 | cyclin dependent kinase 1 | 0.01847032 | 1.803062543 |
| 8 | RCC1 | regulator of chromosome condensation 1 | 0.007265371 | 1.727042562 |
| 9 | RPL26 | ribosomal protein L26 | 0.011706077 | 1.636955215 |
| 10 | RPL10 | ribosomal protein L10 | 0.000538004 | 1.635221166 |
| 11 | NUP43 | nucleoporin 43 | 0.008195855 | 1.632862465 |
| 12 | FUBP3 | far upstream element binding protein 3 | 0.029189622 | 1.617421008 |
| 13 | FBXW11 | F-box and WD repeat domain containing 11 | 0.076761322 | 1.57234785 |
| 14 | PRPS2 | phosphoribosyl pyrophosphate synthetase 2 | 0.029868456 | 1.570681683 |
| 15 | RARS | arginyl-tRNA synthetase | 0.007265371 | 1.510380471 |
| 16 | HNRNPA3 | heterogeneous nuclear ribonucleoprotein A3 | 0.009556945 | 1.429504559 |
| 17 | SLC25A5 | solute carrier family 25 member 5 | 0.000250642 | 1.426609786 |
| 18 | EIF4A2 | eukaryotic translation initiation factor 4A2 | 0.032395672 | 1.387976198 |
| 19 | NCBP2 | nuclear cap binding protein subunit 2 | 0.054499316 | 1.366077171 |
| 20 | BCCIP | BRCA2 and CDKN1A interacting protein | 0.040935557 | 1.33229145 |
| 21 | FAU | FAU, ubiquitin like and ribosomal protein S30 fusion | 0.074353133 | 1.292640825 |
| 22 | UQCRB | ubiquinol-cytochrome c reductase binding protein | 0.044358822 | 1.283025687 |
| 23 | PMPCB | peptidase, mitochondrial processing beta subunit | 0.010250639 | 1.27205479 |
| 24 | PDHA1 | pyruvate dehydrogenase E1 alpha 1 subunit | 0.00218117 | 1.269166906 |
| 25 | MAPRE1 | microtubule associated protein RP/EB family member 1 | 0.049156453 | 1.266782661 |
| 26 | SKP1 | S-phase kinase associated protein 1 | 0.071604825 | 1.254078271 |
| 27 | HMGB1 | high mobility group box 1 | 0.012694951 | 1.230704281 |
| 28 | ACAT1 | acetyl-CoA acetyltransferase 1 | 0.084656497 | 1.229396761 |
| 29 | SHMT2 | serine hydroxymethyltransferase 2 | 0.029915865 | 1.229351493 |
| 30 | COPZ1 | coatomer protein complex subunit zeta 1 | 0.074353133 | 1.215072132 |
| 31 | USP14 | ubiquitin specific peptidase 14 | 0.010498702 | 1.178426319 |
| 32 | RPS15A | ribosomal protein S15a | 0.069434337 | 1.171489242 |
| 33 | SDHC | succinate dehydrogenase complex subunit | 0.076718561 | 1.153724537 |
| 34 | GLYR1 | glyoxylate reductase 1 homolog | 0.075169122 | 1.148655113 |
| 35 | OAT | ornithine aminotransferase | 0.005882567 | 1.134150228 |
| 36 | VPS29 | VPS29, retromer complex component | 0.088864364 | 1.122312686 |
| 37 | DNAJC21 | DnaJ heat shock protein family (Hsp40) member C21 | 0.096374154 | 1.120757635 |
| 38 | B2M | beta-2-microglobulin | 0.083259285 | 1.108883017 |
| 39 | FOXC1 | forkhead box C1 | 0.06145435 | 1.108340457 |
| 40 | SKI | SKI proto-oncogene | 0.088864364 | 1.102900252 |
| 41 | TGOLN2 | trans-golgi network protein 2 | 0.037609589 | 1.101849221 |
| 42 | FBL | fibrillarin | 0.044358822 | 1.097579167 |
| 43 | TFAP2A | transcription factor AP-2 alpha | 0.049183194 | 1.09566041 |
| 44 | PRKAR1A | protein kinase cAMP-dependent type I regulatory subunit alpha | 0.055044332 | 1.09362372 |
| 45 | NEFL | neurofilament light | 0.063550637 | 1.075315115 |
| 46 | PTTG1 | pituitary tumor-transforming 1 | 0.03996427 | 1.073127863 |
| 47 | PREP | prolyl endopeptidase | 0.099910871 | 1.06731661 |
| 48 | EIF3L | eukaryotic translation initiation factor 3 subunit L | 0.065101199 | 1.062215988 |
| 49 | HSPA9 | heat shock protein family A (Hsp70) member 9 | 0.013159468 | 1.049985495 |
| 50 | SRSF1 | serine and arginine rich splicing factor 1 | 0.081526317 | 1.034177714 |
| 51 | CDC23 | cell division cycle 23 | 0.035728724 | 1.033849332 |
| 52 | NAE1 | NEDD8 activating enzyme E1 subunit 1 | 0.054499316 | 1.0329236 |
| 53 | CDC37 | cell division cycle 37 | 0.076761322 | 1.005715312 |
| 54 | NCAPG2 | non-SMC condensin II complex subunit G2 | 0.048942052 | 1.000694023 |
| 55 | MTREX | Mtr4 exosome RNA helicase | 0.054604073 | 0.99292805 |
| 56 | CCT7 | chaperonin containing TCP1 subunit 7 | 0.002282515 | 0.977419566 |
| 57 | KAT7 | lysine acetyltransferase 7 | 0.045918547 | 0.974711617 |
| 58 | RPS24 | ribosomal protein S24 | 0.007265371 | 0.970173253 |
| 59 | HES1 | hes family bHLH transcription factor 1 | 0.068638154 | 0.959729472 |
| 60 | RBM17 | RNA binding motif protein 17 | 0.076761322 | 0.946115122 |
| 61 | RPS3A | ribosomal protein S3A | 0.098865311 | 0.921705513 |
| 62 | AUP1 | AUP1. lipid droplet regulating VLDL assembly factor | 0.067871964 | 0.904287613 |
| 63 | HNRNPA1 | heterogeneous nuclear ribonucleoprotein A1 | 0.000431183 | 0.895925461 |
| 64 | RPL5 | ribosomal protein L5 | 0.001910138 | 0.895501706 |
| 65 | SUCLG1 | succinate-CoA ligase alpha subunit | 0.065734544 | 0.868052522 |
| 66 | BUB1 | BUB1 mitotic checkpoint serine/threonine kinase | 0.056010491 | 0.849315094 |
| 67 | INTS14 | integrator complex subunit 14 | 0.069509177 | 0.843657405 |
| 68 | CCT4 | chaperonin containing TCP1 subunit | 0.002282515 | 0.835602103 |
| 69 | INTS13 | integrator complex subunit 13 | 0.099231448 | 0.828717986 |
| 70 | PLS3 | plastin 3 | 0.040594481 | 0.8130572 |
| 71 | EIF5 | eukaryotic translation initiation factor 5 | 0.03403791 | 0.812449142 |
| 72 | NARS | asparaginyl-tRNA synthetase | 0.037609589 | 0.809858958 |
| 73 | CCT3 | chaperonin containing TCP1 subunit 3 | 0.057566318 | 0.806374214 |
| 74 | KIF22 | kinesin family member 22 | 0.076761322 | 0.792676124 |
| 75 | CCT6A | chaperonin containing TCP1 subunit 6A | 0.028782703 | 0.786750634 |
| 76 | SEPTIN7 | septin 7 | 0.062370622 | 0.77110045 |
| 77 | PAICS | phosphoribosylaminoimidazole carboxylase and phosphoribosylaminoimidazolesuccinocarboxamide synthase | 0.09282529 | 0.762744164 |
| 78 | ACBD3 | acyl-CoA binding domain containing 3 | 0.096199008 | 0.738281486 |
| 79 | EIF4H | eukaryotic translation initiation factor 4H | 0.05417421 | 0.718503713 |
| 80 | EIF2S3 | eukaryotic translation initiation factor 2 subunit gamma | 0.052916229 | 0.704489885 |
| 81 | FKBP4 | FK506 binding protein 4 | 0.048451346 | 0.697700308 |
| 82 | EEF1A1 | eukaryotic translation elongation factor 1 alpha 1 | 0.001499086 | 0.663548054 |
| 83 | XRCC6 | X-ray repair cross complementing 6 | 0.017242388 | 0.644500151 |
| 84 | HNRNPA2B1 | heterogeneous nuclear ribonucleoprotein A2/B1 | 0.052916229 | 0.611694961 |
| # | Gene Symbol | Description | p.adj | Shrunken LFC |
|---|---|---|---|---|
| 1 | ATN1 | atrophin 1 | 0.000250642 | −2,383,968,333 |
| 2 | HHIPL1 | HHIP like 1 | 0.099910871 | −2,108,283,211 |
| 3 | LAMA5 | laminin subunit alpha 5 | 1.04737E-07 | −2,099,435,635 |
| 4 | NOMO3 | NODAL modulator 3 | 0.034998577 | −1,772,216,098 |
| 5 | GSK3A | glycogen synthase kinase 3 alpha | 0.086893975 | −1,657,765,364 |
| 6 | OCIAD2 | OCIA domain containing 2 | 0.041499491 | −1,602,956,347 |
| 7 | AUTS2 | AUTS2, activator of transcription and developmental regulator | 0.007265371 | −1,450,228,921 |
| 8 | CAMSAP1 | calmodulin regulated spectrin associated protein 1 | 0.020568057 | −1,399,881,818 |
| 9 | PRELID1 | PRELI domain containing 1 | 0.022809358 | −1,365,693,298 |
| 10 | ZNF274 | zinc finger protein 274 | 0.01422397 | −1,355,991,291 |
| 11 | GAK | cyclin G associated kinase | 0.010498702 | −1,351,541,744 |
| 12 | LTBP1 | latent transforming growth factor beta binding protein 1 | 0.000250642 | −1,336,690,238 |
| 13 | MAX | MYC associated factor X | 0.040594481 | −1,320,705,011 |
| 14 | HIST1H2BC | histone cluster 1 H2B family member c | 0.064581433 | −1,314,815,037 |
| 15 | EDC4 | enhancer of mRNA decapping 4 | 0.042249906 | −1,307,590,139 |
| 16 | ASCC2 | activating signal cointegrator 1 complex subunit 2 | 0.029868456 | −1,256,837,357 |
| 17 | ENOSF1 | enolase superfamily member 1 | 0.081515108 | −1,250,428,661 |
| 18 | CTCF | CCCTC-binding factor | 0.007366565 | −1,239,742,061 |
| 19 | IQGAP3 | IQ motif containing GTPase activating protein 3 | 0.033902778 | −119,141,635 |
| 20 | RPLP1 | ribosomal protein lateral stalk subunit P1 | 0.002534611 | −1,125,473,816 |
| 21 | C6orf106 | chromosome 6 open reading frame 106 | 0.052916229 | −1,106,301,158 |
| 22 | AEN | apoptosis enhancing nuclease | 0.010498702 | −1,105,573,856 |
| 23 | CDK5RAP2 | CDK5 regulatory subunit associated protein 2 | 0.055629954 | −1,096,112,547 |
| 24 | ATP13A2 | ATPase cation transporting 13A2 | 0.015260908 | −1,083,453,368 |
| 25 | ITPR3 | inositol 1,4,5-trisphosphate receptor type 3 | 0.052068978 | −1,046,481,817 |
| 26 | COL6A1 | collagen type VI alpha 1 chain | 0.059954251 | −1,046,201,279 |
| 27 | CYB5B | cytochrome b5 type B | 0.061939362 | −1,045,026,726 |
| 28 | FASN | fatty acid synthase | 0.020429889 | −1,006,556,157 |
| 29 | TBRG4 | transforming growth factor beta regulator 4 | 0.031530019 | −1,000,246,535 |
| 30 | SERP1 | stress associated endoplasmic reticulum protein 1 | 0.098408591 | −0.99348297 |
| 31 | SBF1 | SET binding factor 1 | 0.020568057 | −0.991430768 |
| 32 | HIST1H4E | histone cluster 1 H4 family member e | 0.029649385 | −0.985384473 |
| 33 | DDT | D-dopachrome tautomerase | 0.054499316 | −0.98064313 |
| 34 | FBRS | fibrosin | 0.035039202 | −0.969773425 |
| 35 | STRN4 | striatin 4 | 0.089949483 | −0.943078161 |
| 36 | WDR62 | WD repeat domain 62 | 0.035792707 | −0.923170831 |
| 37 | URB2 | URB2 ribosome biogenesis 2 homolog (S. cerevisiae) | 0.085713486 | −0.89953143 |
| 38 | PIEZO1 | piezo type mechanosensitive ion channel component 1 | 0.026333758 | −0.891075548 |
| 39 | KMT2D | lysine methyltransferase 2D | 0.052916229 | −0.889992224 |
| 40 | SDCBP | syndecan binding protein | 0.05742347 | −0.882057197 |
| 41 | SMARCC1 | SWI/SNF related, matrix associated, actin dependent regulator of chromatin subfamily c member 1 | 0.035728724 | −0.870997243 |
| 42 | HECTD4 | HECT domain E3 ubiquitin protein ligase 4 | 0.04412228 | −0.868499831 |
| 43 | ARHGAP35 | Rho GTPase activating protein 35 | 0.072048538 | −0.86688062 |
| 44 | SYMPK | symplekin | 0.019920301 | −0.855418984 |
| 45 | ABCA3 | ATP binding cassette subfamily A member 3 | 0.096753082 | −0.841808007 |
| 46 | HIST1H2AC | histone cluster 1 H2A family member c | 0.038804021 | −0.834943336 |
| 47 | KIF1B | kinesin family member 1B | 0.057776732 | −0.834338228 |
| 48 | RETREG3 | reticulophagy regulator family member 3 | 0.089018972 | −0.826365716 |
| 49 | CLUH | clustered mitochondria homolog | 0.079547821 | −0.822733101 |
| 50 | PELP1 | proline, glutamate and leucine rich protein 1 | 0.072048538 | −0.803585995 |
| 51 | COL2A1 | collagen type II alpha 1 chain | 0.020704772 | −0.803529736 |
| 52 | CLPTM1 | CLPTM1, transmembrane protein | 0.080528426 | −0.79290716 |
| 53 | NUP210 | nucleoporin 210 | 0.020704772 | −0.78932396 |
| 54 | SFSWAP | splicing factor SWAP | 0.094719096 | −0.788876906 |
| 55 | IGF1R | insulin like growth factor 1 receptor | 0.086783977 | −0.784038911 |
| 56 | UBE2D2 | ubiquitin conjugating enzyme E2 D2 | 0.035728724 | −0.762644136 |
| 57 | MEGF8 | multiple EGF like domains 8 | 0.062032886 | −0.741762602 |
| 58 | DDB1 | damage specific DNA binding protein 1 | 0.09771499 | −0.711925724 |
| 59 | MDC1 | mediator of DNA damage checkpoint 1 | 0.042983832 | −0.711259755 |
| 60 | FLNA | filamin A | 0.007366565 | −0.701321881 |
| 61 | SUGP2 | SURP and G-patch domain containing 2 | 0.092506997 | −0.67863598 |
| 62 | MAGED1 | MAGE family member D1 | 0.09697418 | −0.662214919 |
| 63 | UPF1 | UPF1, RNA helicase and ATPase | 0.099910871 | −0.661569626 |
| 64 | CLTC | clathrin heavy chain | 0.061939362 | −0.660003656 |
| 65 | BAG6 | BCL2 associated athanogene 6 | 0.074741705 | −0.656625382 |
| 66 | HMGA1 | high mobility group AT-hook 1 | 0.076718561 | −0.626612021 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gopanenko, A.V.; Kolobova, A.V.; Tupikin, A.E.; Kabilov, M.R.; Malygin, A.A.; Karpova, G.G. Knockdown of the Ribosomal Protein eL38 in HEK293 Cells Changes the Translational Efficiency of Specific Genes. Int. J. Mol. Sci. 2021, 22, 4531. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094531
Gopanenko AV, Kolobova AV, Tupikin AE, Kabilov MR, Malygin AA, Karpova GG. Knockdown of the Ribosomal Protein eL38 in HEK293 Cells Changes the Translational Efficiency of Specific Genes. International Journal of Molecular Sciences. 2021; 22(9):4531. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094531
Chicago/Turabian StyleGopanenko, Alexander V., Alena V. Kolobova, Alexey E. Tupikin, Marsel R. Kabilov, Alexey A. Malygin, and Galina G. Karpova. 2021. "Knockdown of the Ribosomal Protein eL38 in HEK293 Cells Changes the Translational Efficiency of Specific Genes" International Journal of Molecular Sciences 22, no. 9: 4531. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094531