
Application of Nitrate, Ammonium, or Urea Changes the Concentrations of Ureides, Urea, Amino Acids and Other Metabolites in Xylem Sap and in the Organs of Soybean Plants (Glycine max (L.) Merr.)

Abstract
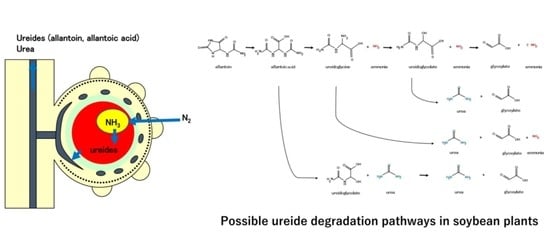
:
1. Introduction
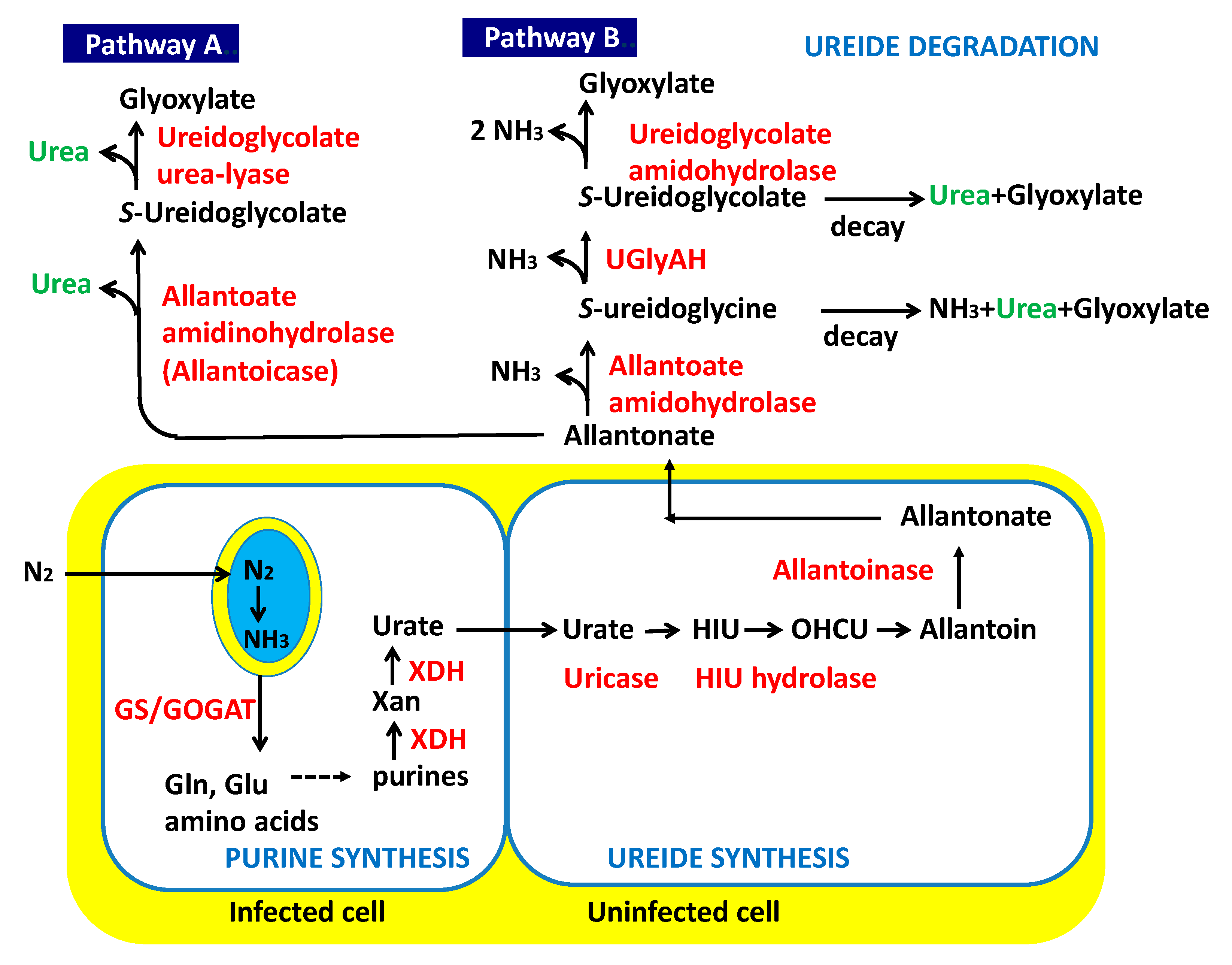
1.1. Ureide Biosynthesis in Soybean Plants
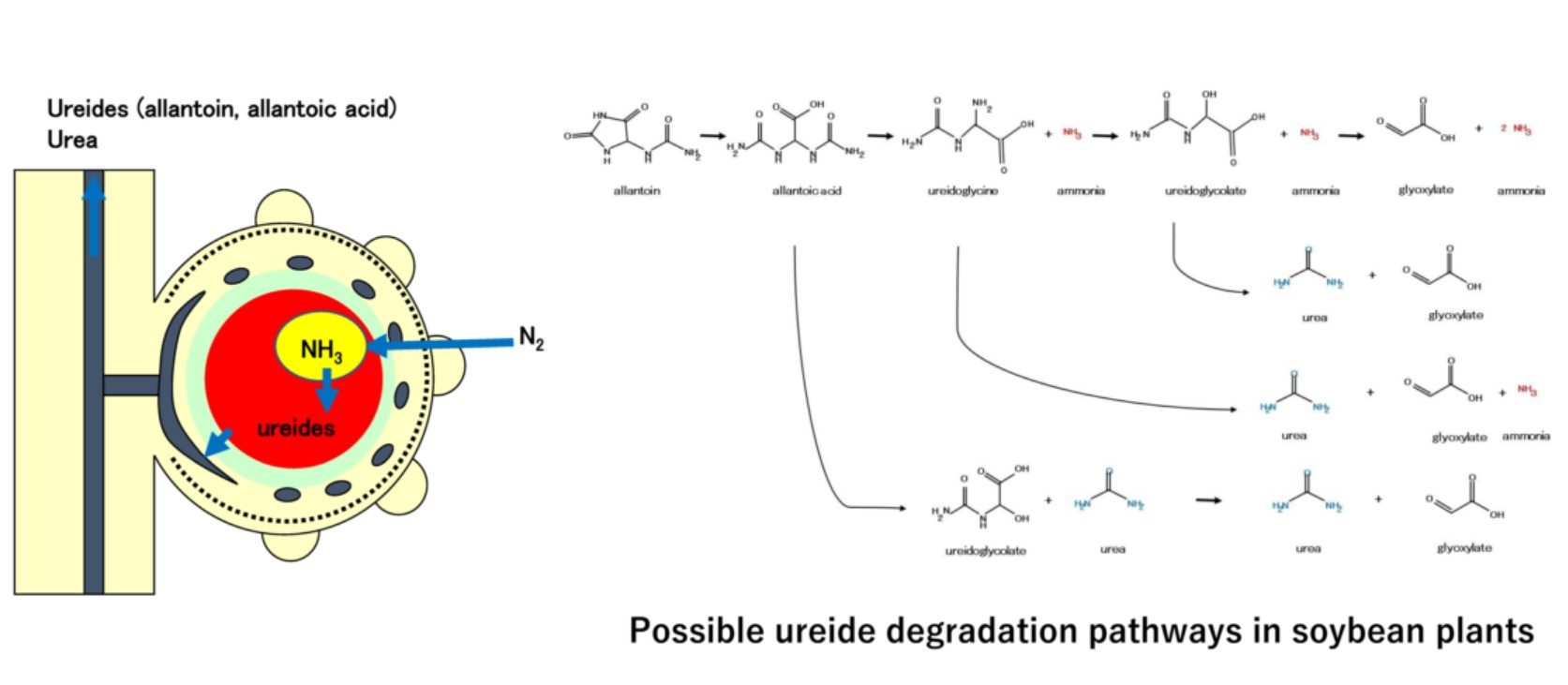
1.2. Degradation of Ureides
2. Results
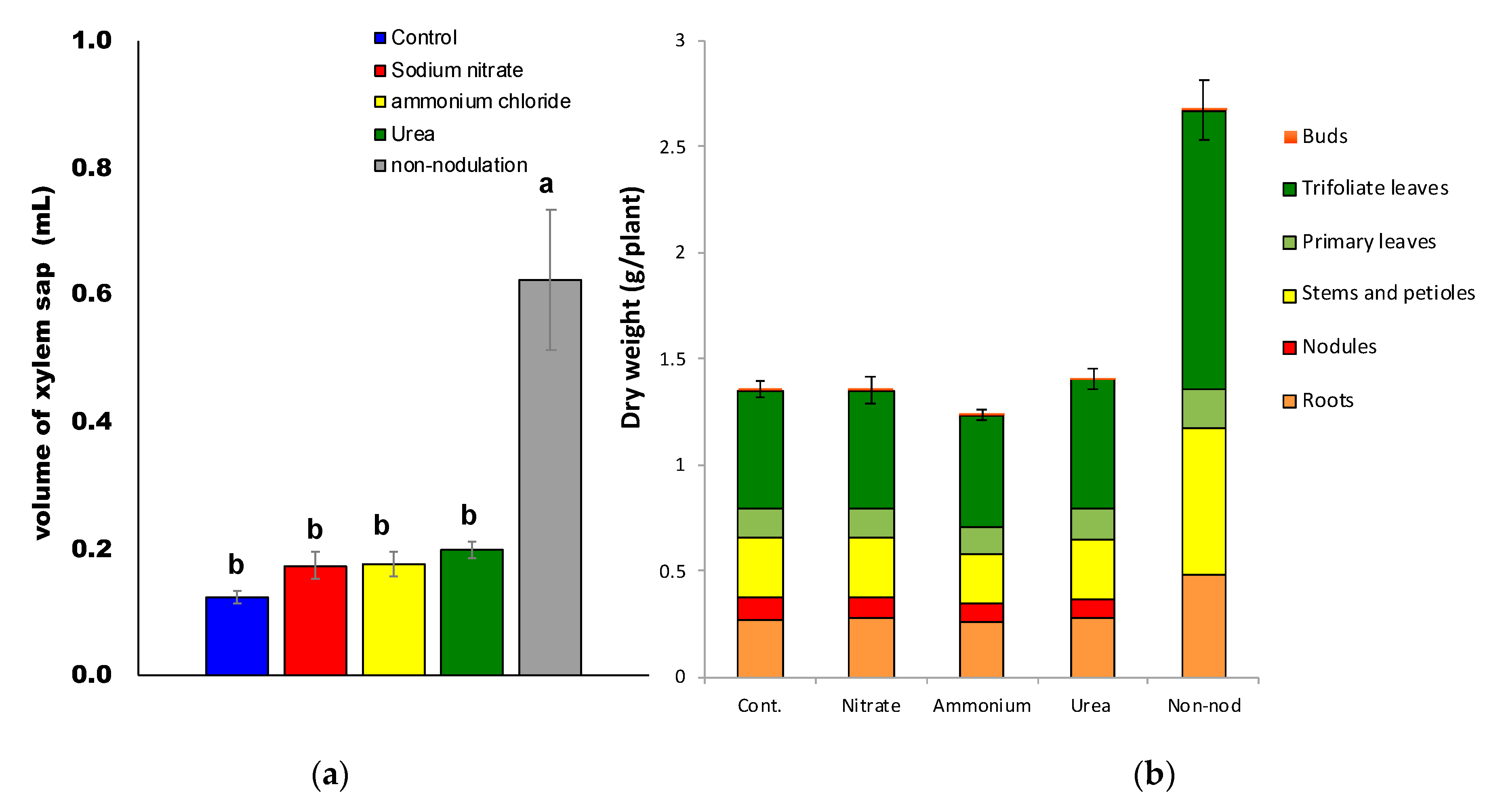
2.1. Effects of Supplying Various Nitrogen Compounds on Plant Growth
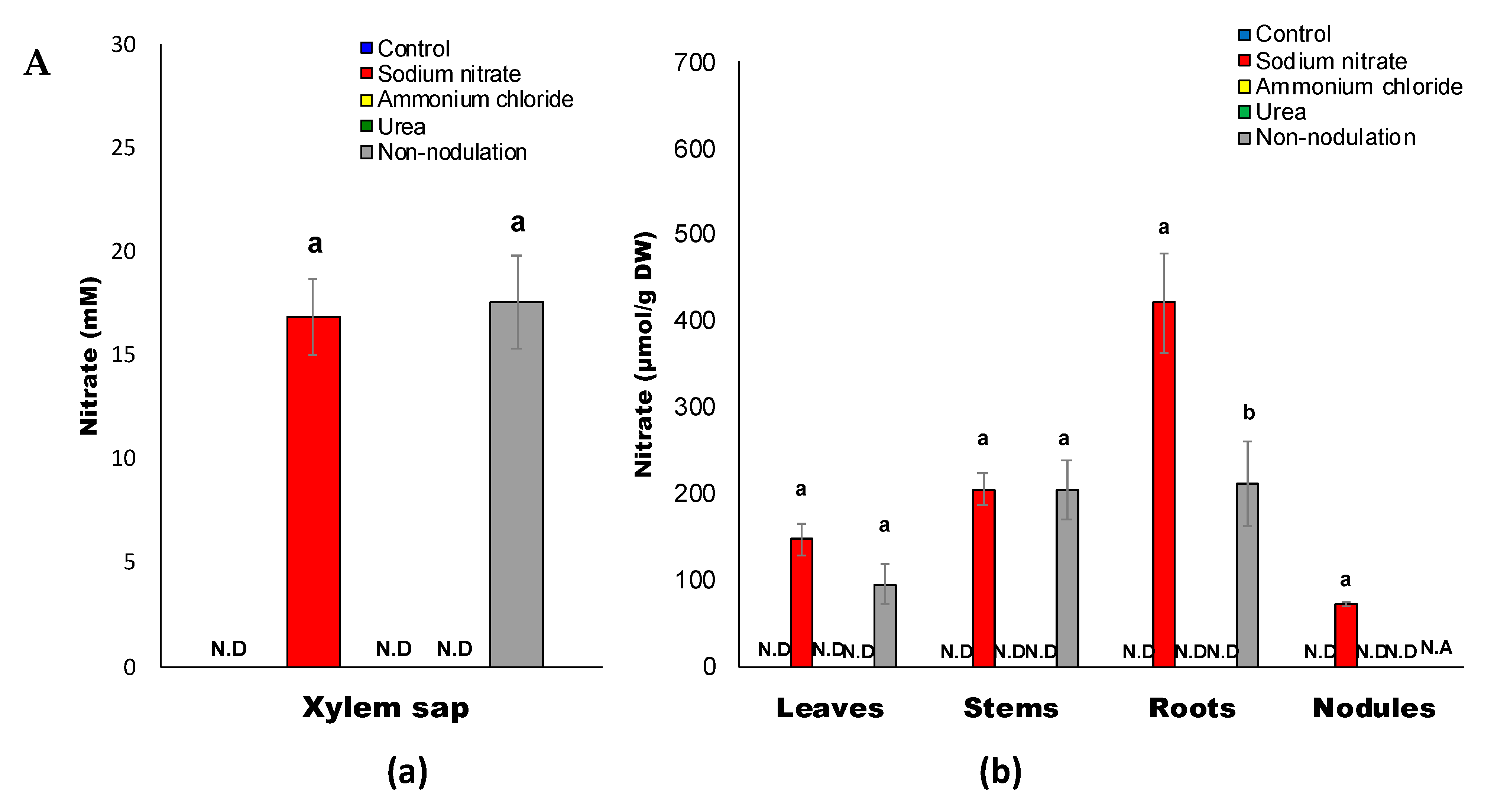
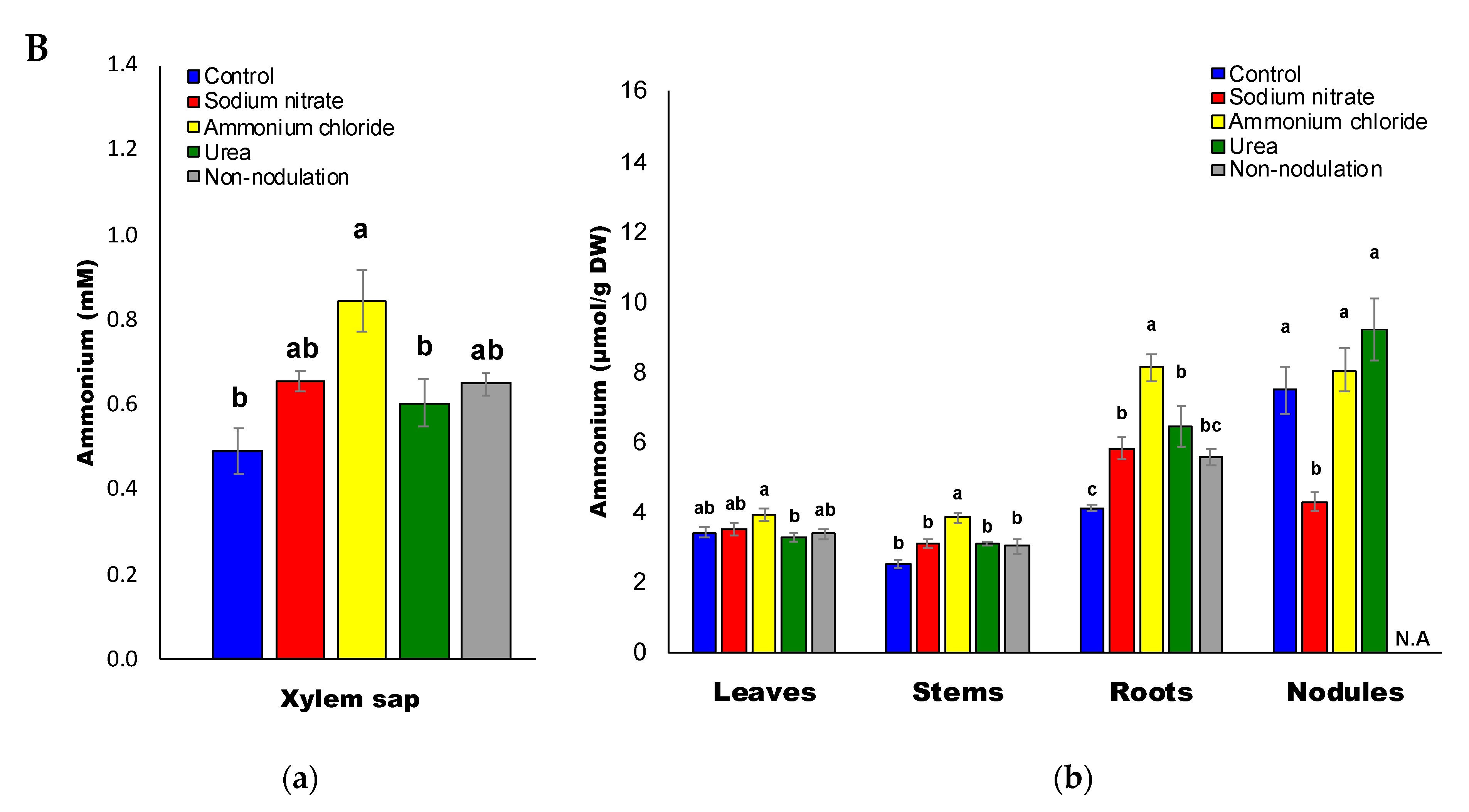
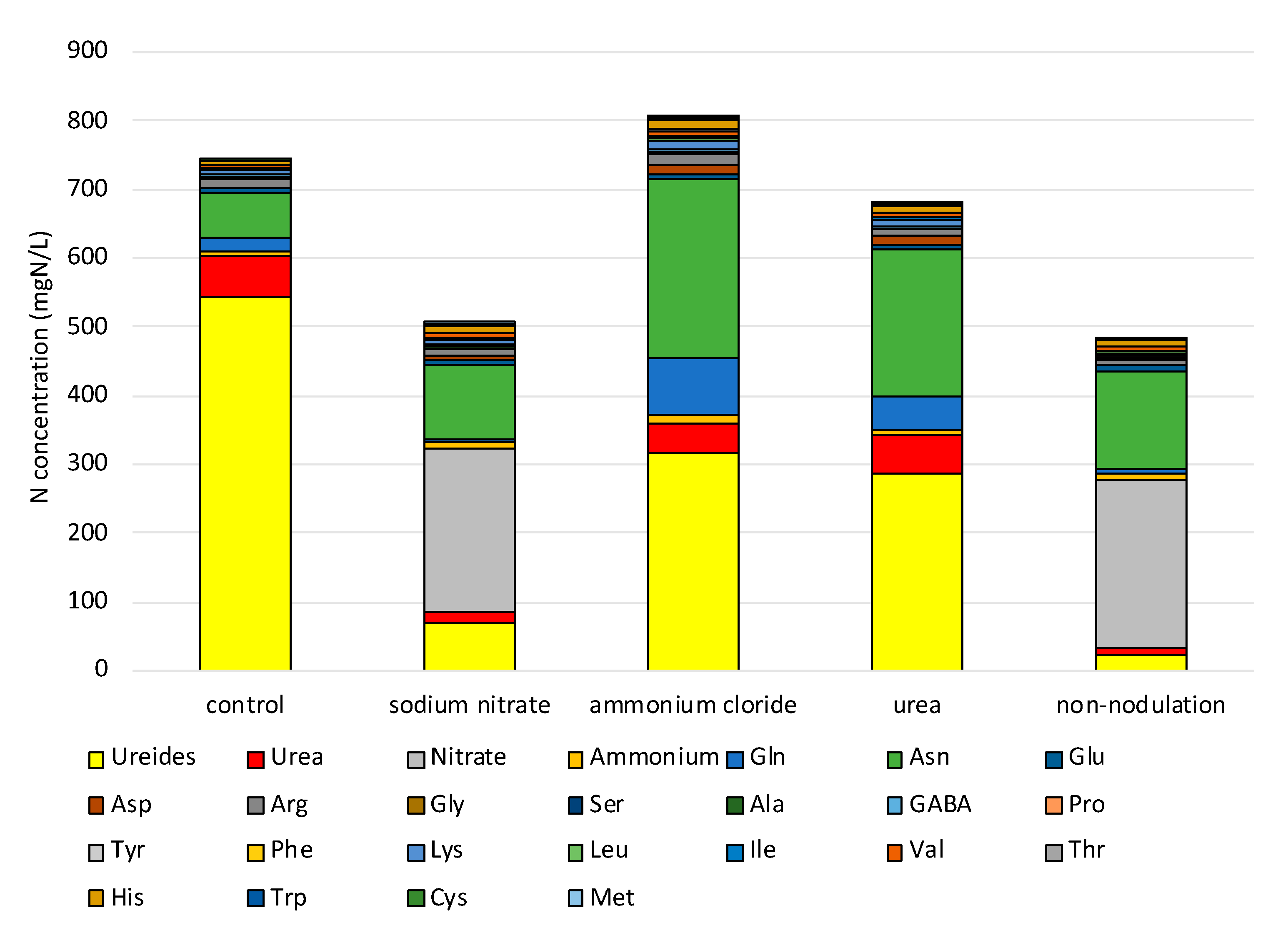
2.2. Inorganic Compositions after Application of Various Chemical Forms
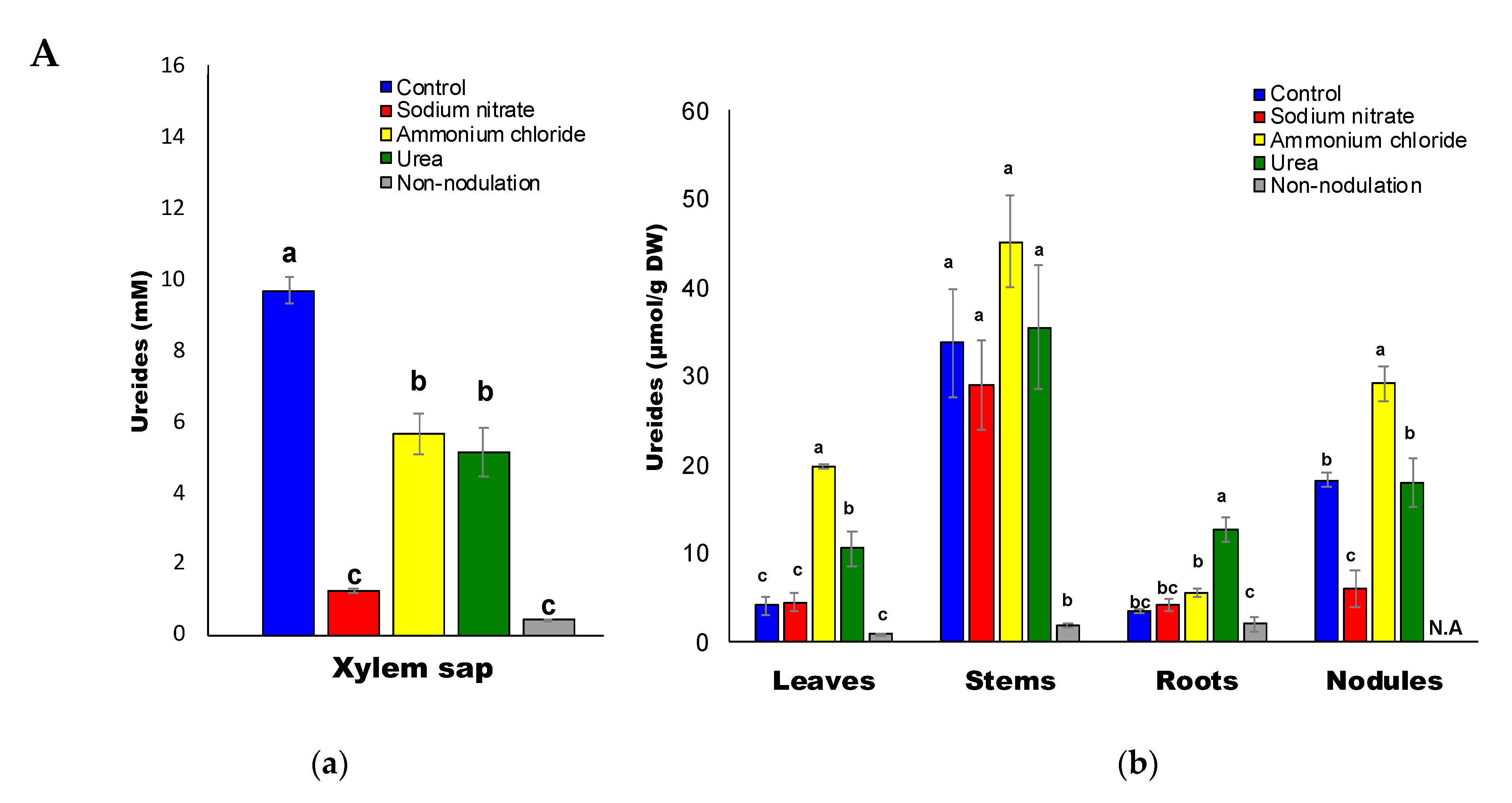
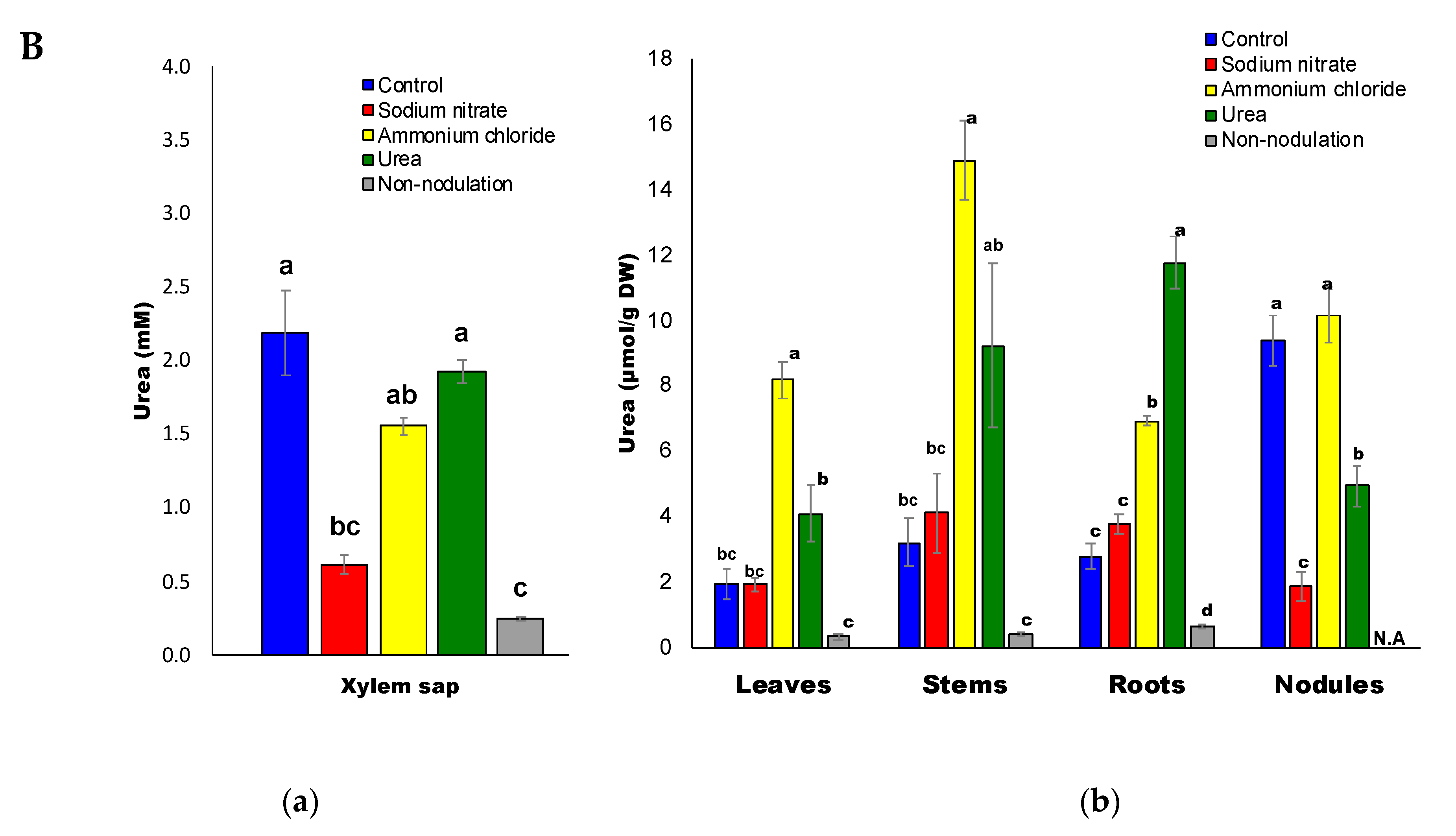
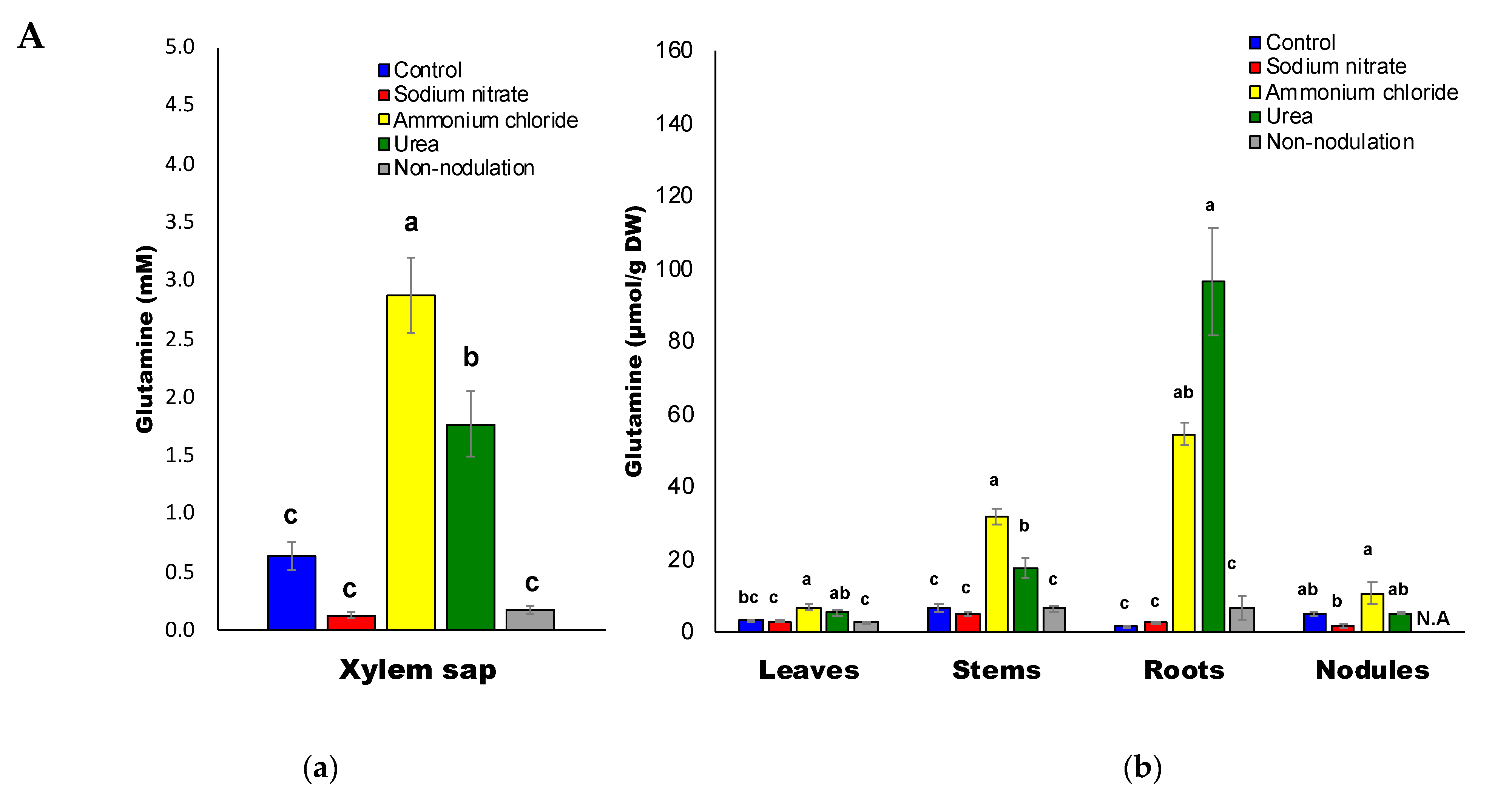
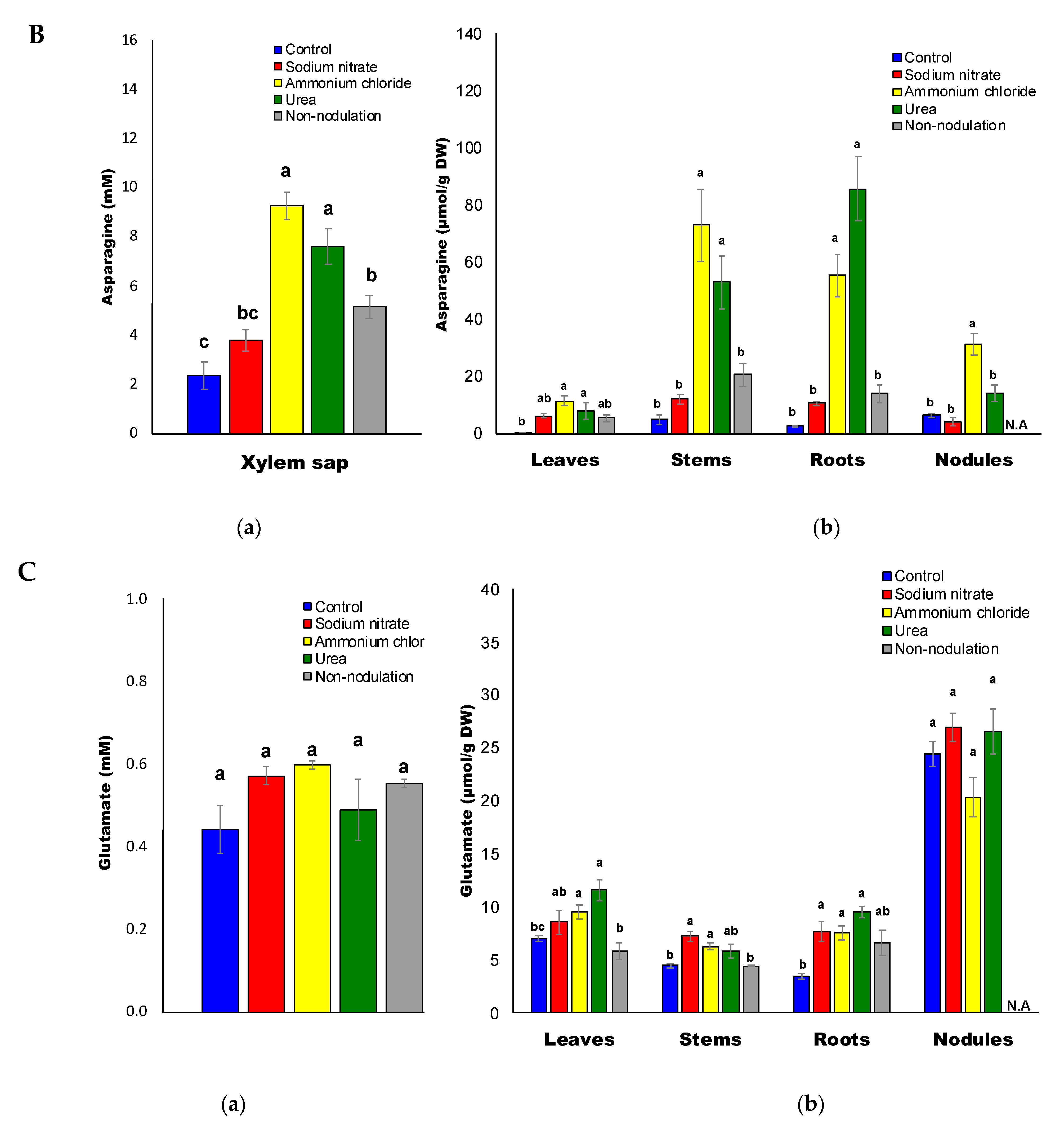
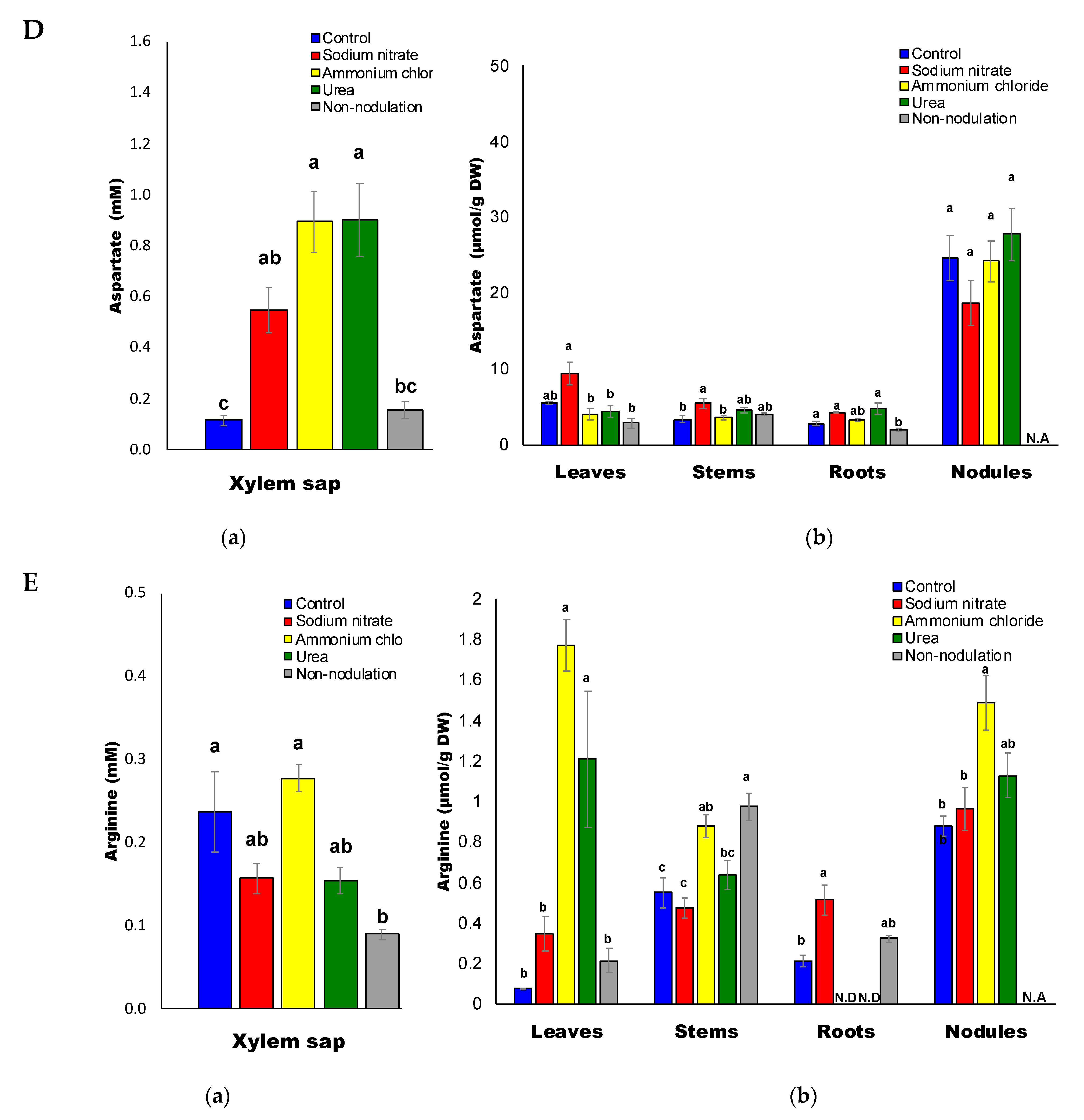
2.3. Ureides and Urea Composition
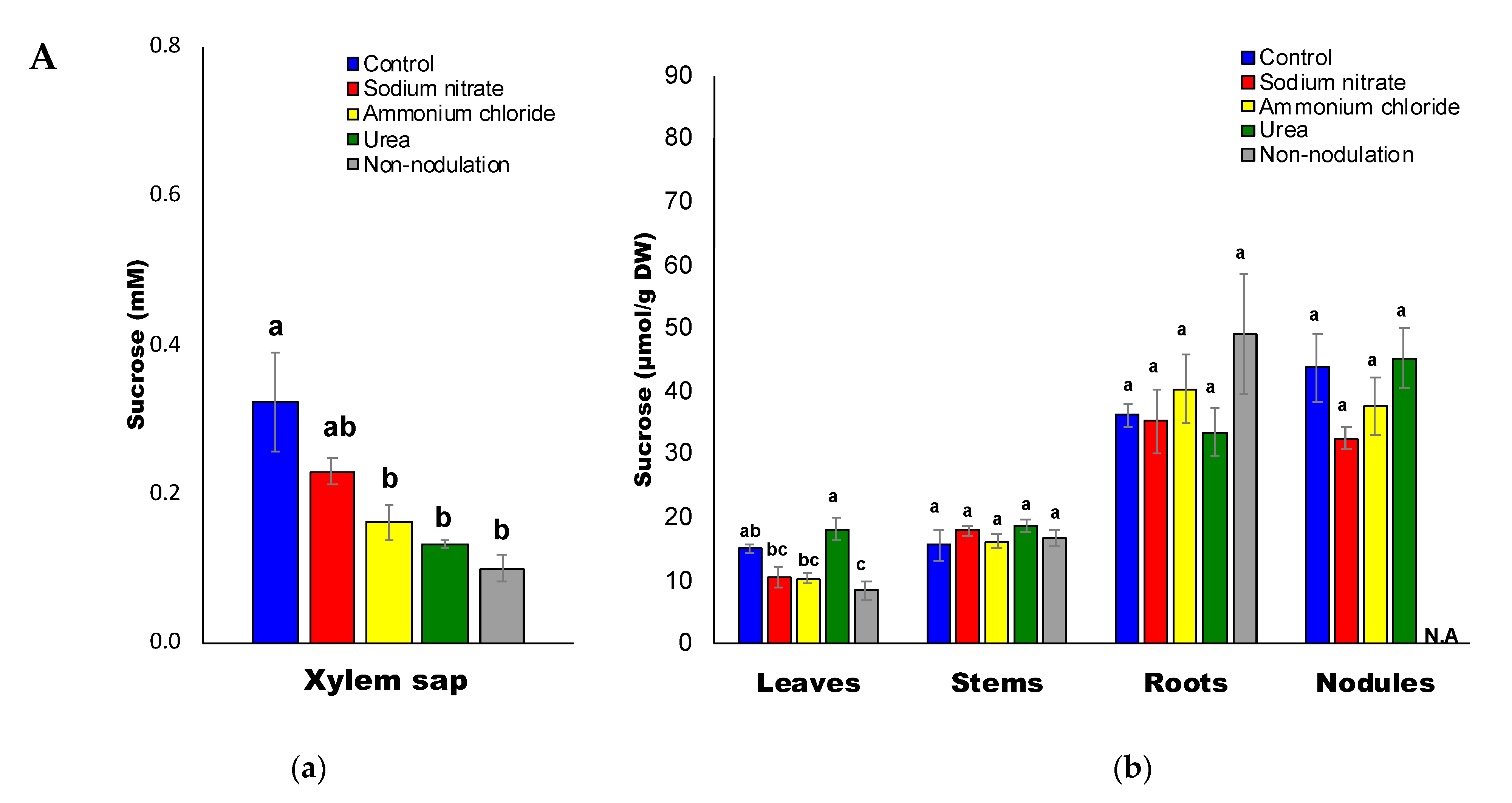
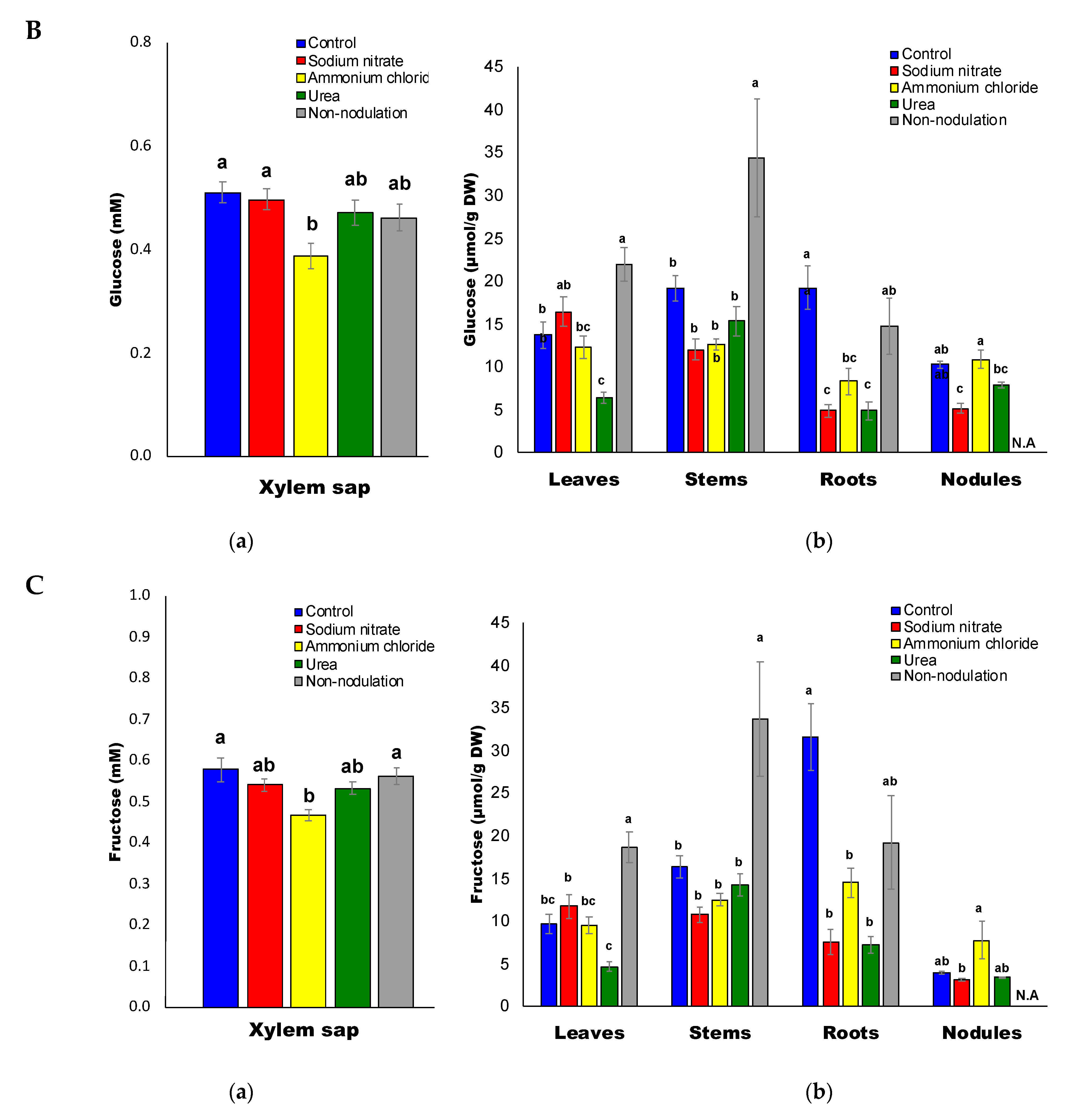
2.4. Sugar Composition
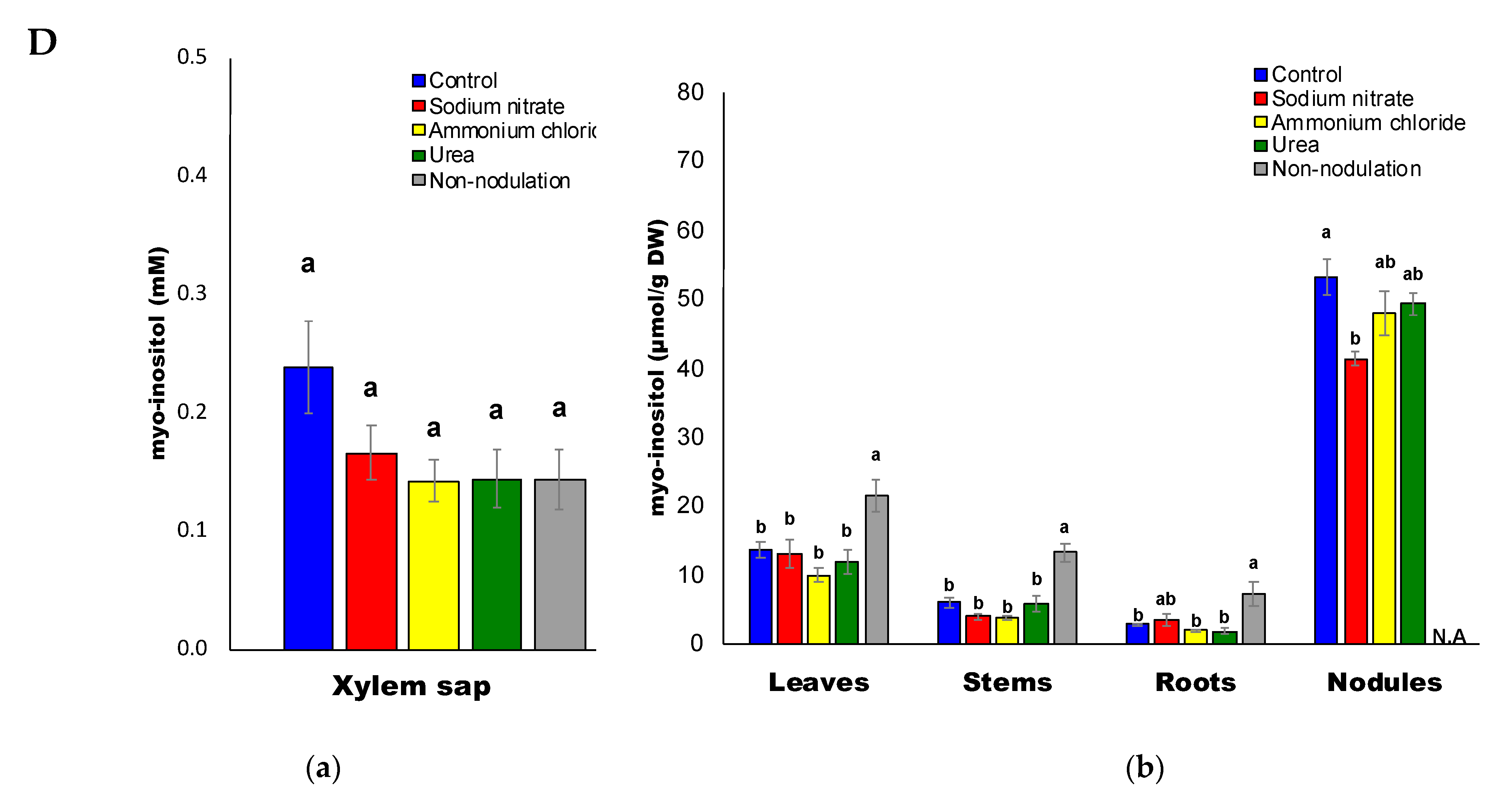
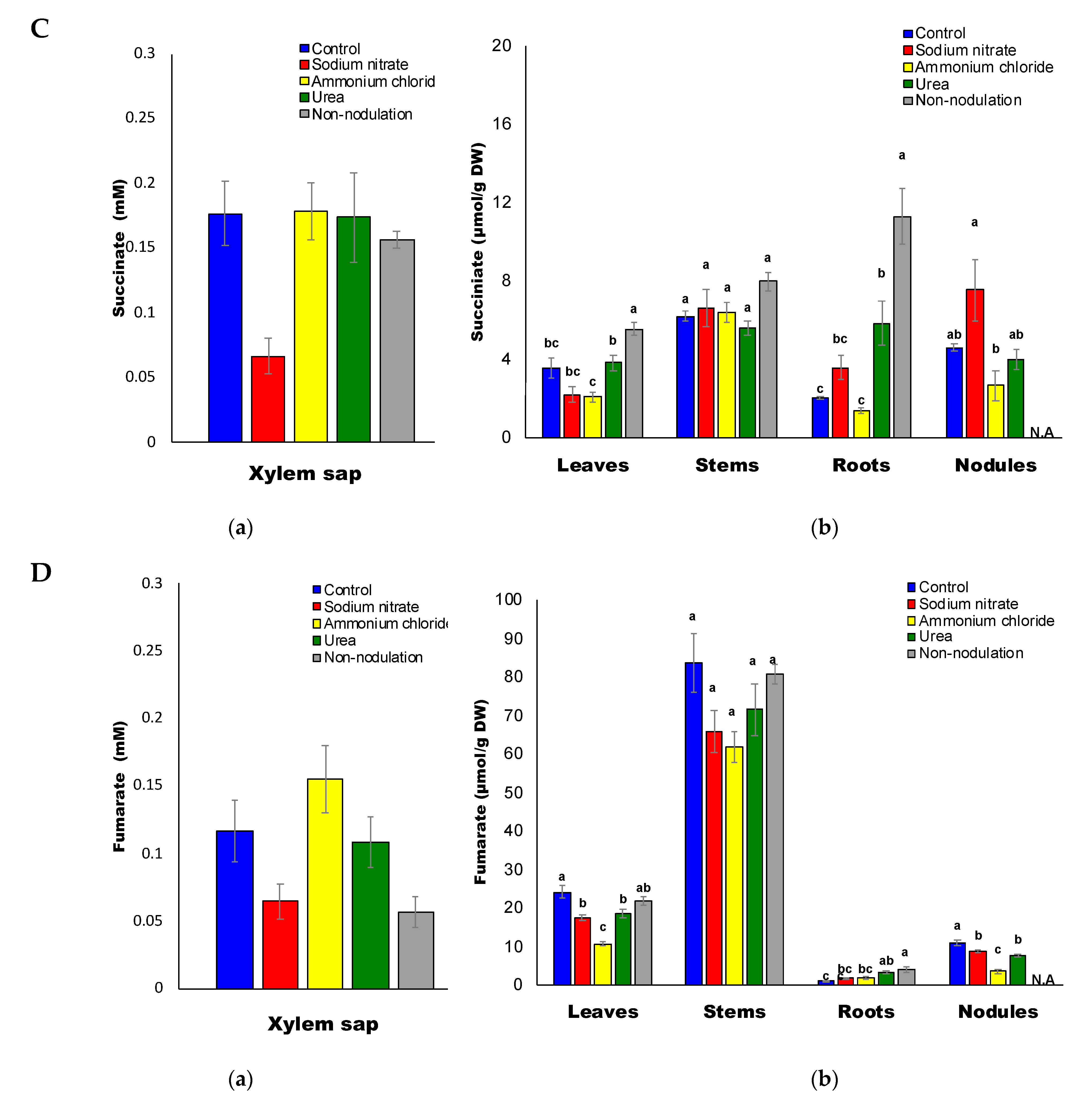
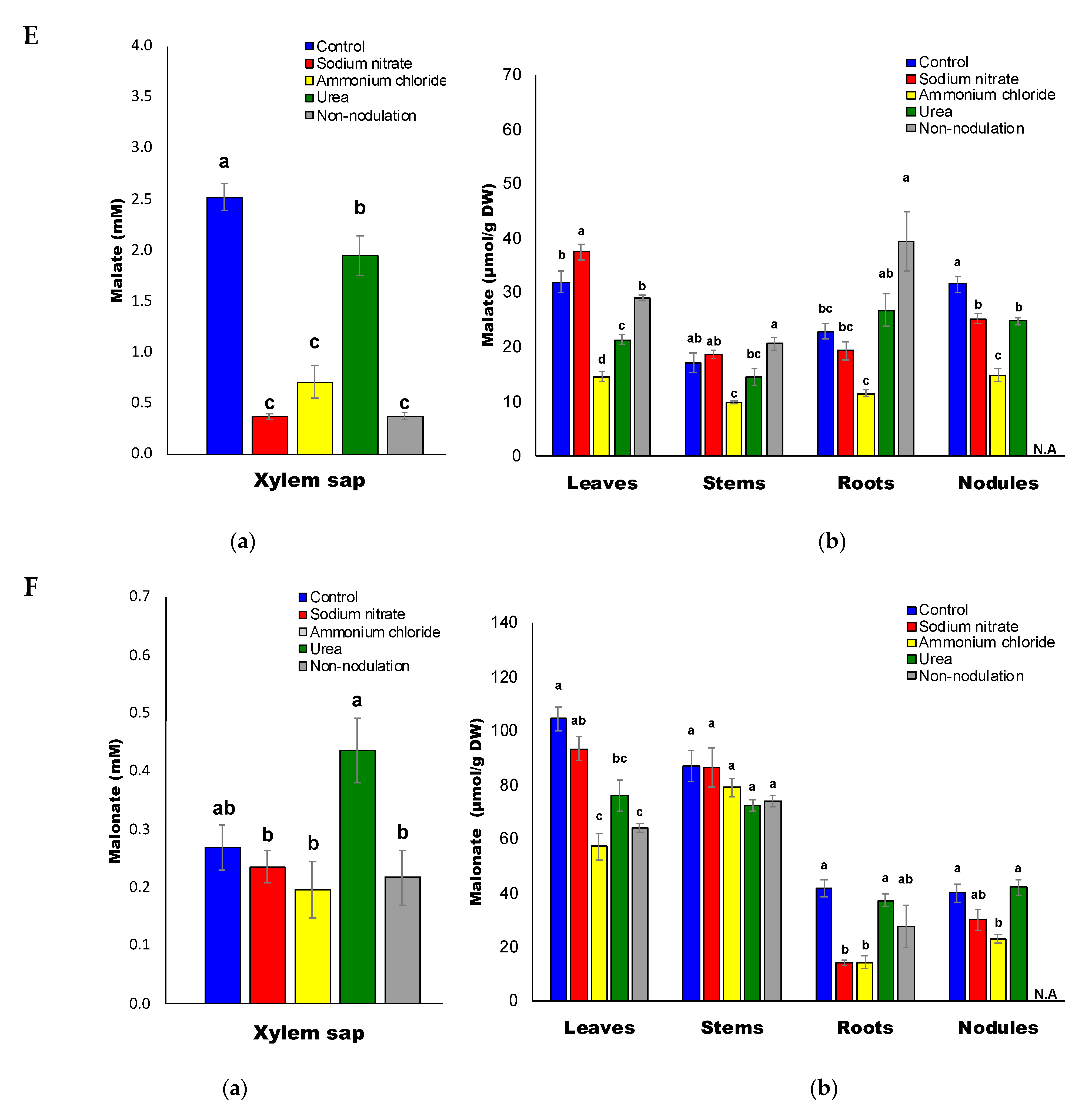
2.5. Organic Acid Composition
2.6. Ratios of the Metabolite Concentration in the Treated Plants to That of the Control Plants
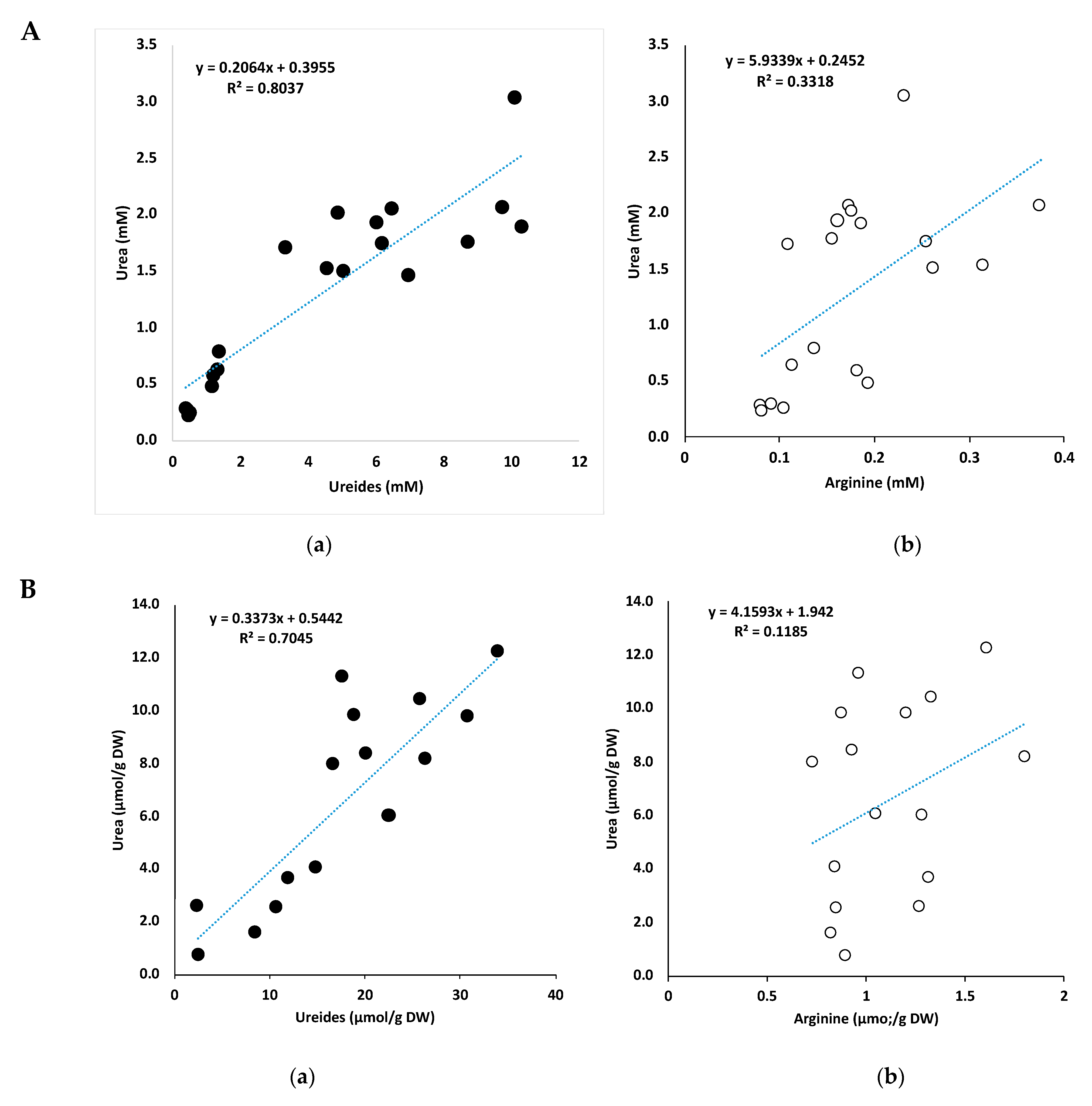
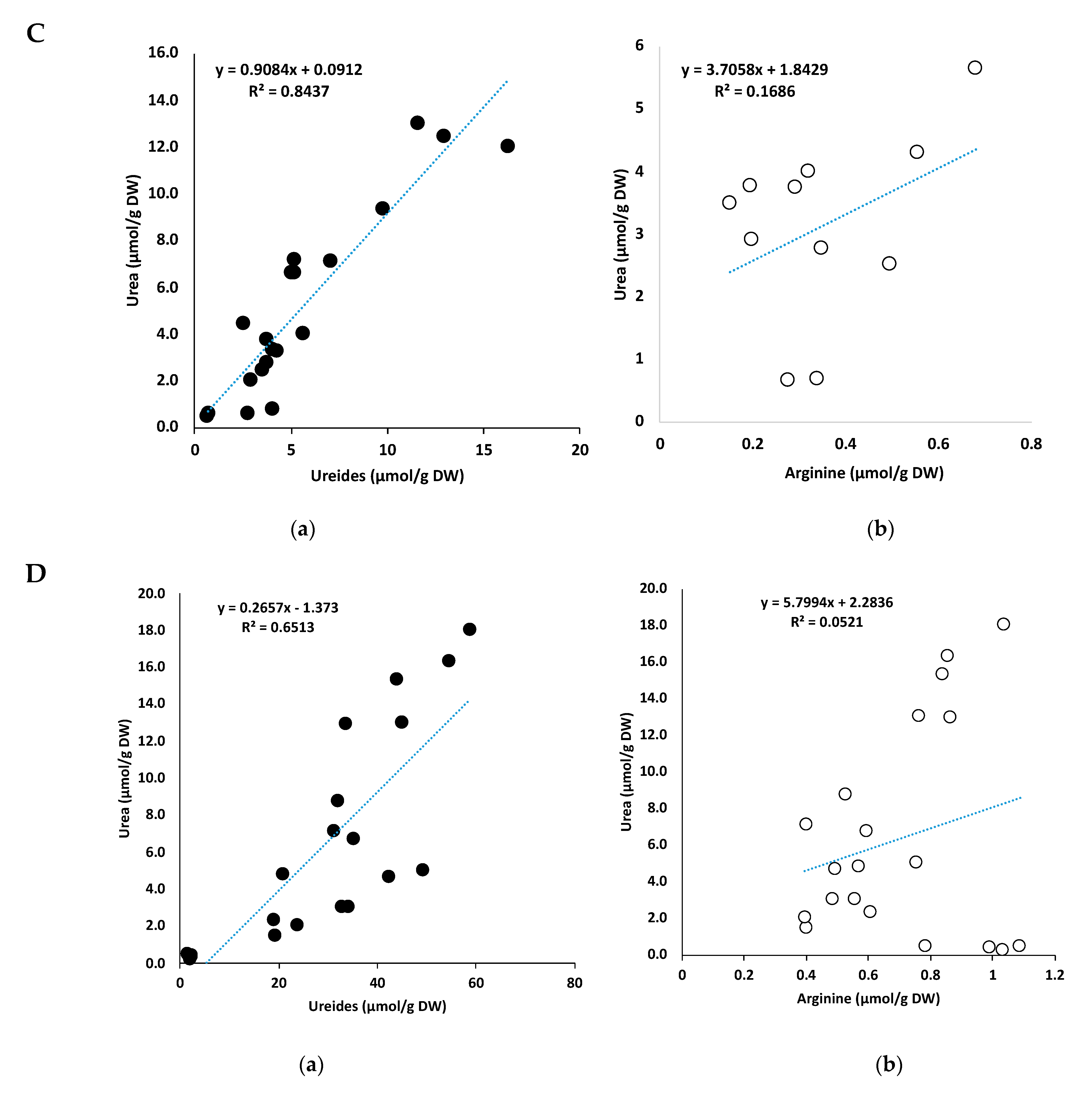
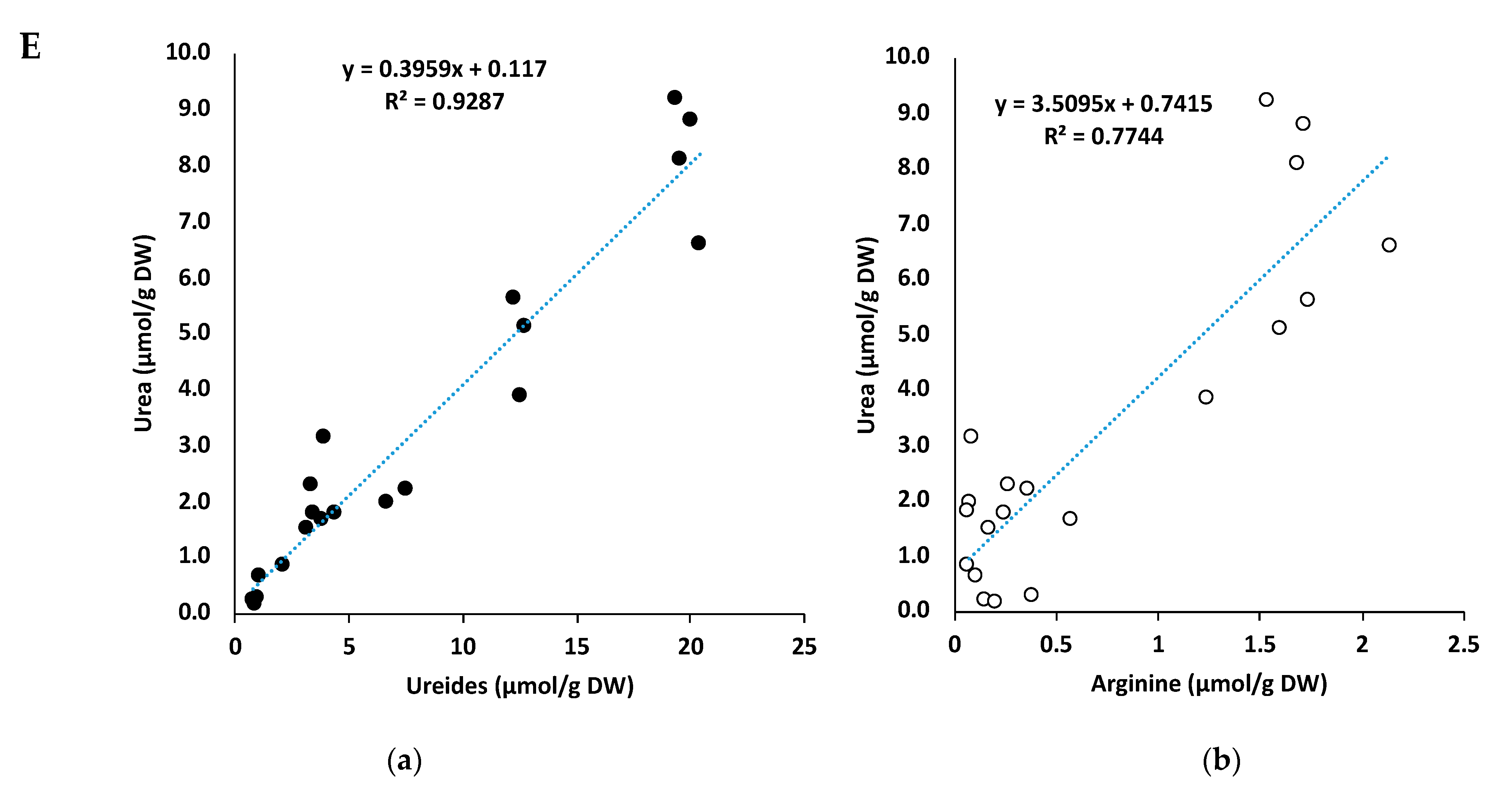
2.7. Correlation between Urea and Ureides
2.8. Confirmation of Urea in Soybean
2.9. Ratios of Urea-N/Ureide-N
3. Discussion
3.1. Characteristics of Nitrogen Transport Compounds in the Xylem
3.2. Urea Formation
3.3. High Accumulation of Asn and Gln with Ammonium and Urea Treatments
3.4. Decrease in Sugars and Organic Acids Concentrations by N Treatments
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Application of Nitrogen Compounds and Plant Sampling
4.3. Analysis of Free Amino Acids and Ammonium
4.4. Analysis of Anion
4.5. Analysis of Urea, Sugars, and Organic Acids
4.6. Analysis of Urea by Gas Chromatograph/Mass Spectrum
4.7. Analysis of Ureides
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gibson, A.H.; Harper, J.E. Nitrate effect on nodulation of soybean by Bradyrhizobium japonicum. Crop Sci. 1985, 25, 497–501. [Google Scholar] [CrossRef]
- Streeter, J.G. Inhibition of legume nodule formation and N2 fixation by nitrate. CRC Crit. Rev. Plant Sci. 1988, 7, 1–23. [Google Scholar] [CrossRef]
- Imsande, J. Inhibition of nodule development in soybean by nitrate or reduced nitrogen. J. Exp. Bot. 1986, 37, 348–355. [Google Scholar] [CrossRef]
- Ohyama, T.; Fujikake, H.; Yashima, H.; Tanabata, S.; Ishikawa, S.; Sato, T. Effect of nitrate on nodulation and nitrogen fixation of soybean. In Soybean Physiology and Biochemistry; El-Shemy, H.A., Ed.; InTech: Rijeka, Croatia, 2012; pp. 333–363. ISBN 978-953-307-534-1. [Google Scholar]
- Harper, J.E. Nitrogen metabolism. In Soybeans: Improvement, Production and Uses, 2nd ed.; Agronomy Monograph: Madison, WI, USA, 1987; pp. 497–533. ISBN 0-989118-090-7. [Google Scholar]
- Bergersen, F.J. Ammonia-an early stable product of nitrogen fixation by soybean root nodules. Aust. J. Biol. Sci. 1965, 18, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, T.; Kumazawa, K. Incorporation of 15N into various nitrogenous compounds in intact soybean nodules after exposure to 15N2 gas. Soil Sci. Plant Nutr. 1978, 24, 525–533. [Google Scholar] [CrossRef] [Green Version]
- Ohyama, T.; Kumazawa, K. Assimilation and transport of nitrogenenous compounds originated from 15N2 fixation and 15NO3 absorption. Soil Sci. Plant Nutr. 1979, 25, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Kouchi, H.; Fukai, K.; Katagiri, H.; Minamisawa, K.; Tajima, S. Isolation and enzymological characterization of infected and uninfected cell protoplasts from root nodules of Glycine max. Physiol. Plant. 1988, 73, 327–334. [Google Scholar] [CrossRef]
- Kouchi, H.; Tsukamoto, M.; Tajima, S. Differential expression of nodule specific (nodulin) genes in the infected, uninfected and cortical cells of soybean (Glycine max.) root nodules. J. Plant Physiol. 1989, 135, 608–617. [Google Scholar] [CrossRef]
- Tajima, S.; Nomura, M.; Kouchi, H. Ureide Biosynthesis in legume nodules. Front. BioSci. 2004, 9, 1374–1381. [Google Scholar] [CrossRef] [Green Version]
- Reinbothe, H.; Mothes, K. Urea, udeides, and guanidines in plants. Annu. Rev. Plant Physiol. 1962, 13, 129–149. [Google Scholar] [CrossRef]
- Bollard, E.G. Nitrogenous compounds in plant xylem sap. Nature 1956, 178, 1189–1190. [Google Scholar] [CrossRef]
- Tomas, R.J.; Schrader, L.S. Ureide Metabolism in higher plants. Phytochemistry 1981, 20, 361–371. [Google Scholar] [CrossRef]
- Kushizaki, M.; Ishizuka, J.; Akamatsu, F. Physiological studies on the nutrition of soybean plants. 2. Effect of nodule formation on nitrogenous constituents of soybeans. J. Sci. Soil Manure Jpn. 1964, 35, 323–327. [Google Scholar]
- Ishizuka, J. Physiological roles of soluble nitrogenous compounds on vegetative growth and seed protein formation of soybean plants in Hokkaido. Res. Bull. Hokkaido Natl. Agric. Exp. Stn. 1970, 101, 51–121. [Google Scholar]
- Matsumoto, T.; Yatazawa, M.; Yamamoto, Y. Incorporation of 15N into allantoin in nodulated soybean plants supplied with 15N2. Plant Cell Physiol. 1977, 18, 459–462. [Google Scholar] [CrossRef]
- Fujihara, S.; Yamaguchi, M. Nitrogen Fixation and Allantoin Formation in Soybean Plants. Agric. Biol. Chem. 1980, 44, 2569–2573. [Google Scholar] [CrossRef]
- McClurem, P.R.; Israel, D.W. Transport of nitrogen in the xylem of soybean plants. Plant Physiol. 1979, 64, 411–416. [Google Scholar] [CrossRef]
- Tomas, R.J.; Schrader, L.S. The assimilaton of ureides in shoot tissues of soybeans. Plant Physiol. 1981, 67, 973–976. [Google Scholar] [CrossRef] [Green Version]
- Cheng, X.-G.; Nomura, M.; Sato, T.; Fujikake, H.; Ohyama, T.; Tajima, S. Effect of exogenous NH4+-N supply on distribution of ureide content in various tissues of alfalfa plants. Medicago sativa. Soil Sci. Plant Nutr. 1999, 45, 921–927. [Google Scholar] [CrossRef] [Green Version]
- Raychaudhuri, A.; Tipton, P.A. Cloning and Expression of the Gene for Soybean Hydroxyisourate Hydrolase. Localization and Implications for Function and Mechanism. Plant Physiol. 2002, 130, 2061–2068. [Google Scholar] [CrossRef] [Green Version]
- Shelp, B.J.; Ireland, R.J. Ureide Metabolism in Leaves of Nitrogen-Fixing Soybean Plants. Plant Physiol. 1985, 77, 779–783. [Google Scholar] [CrossRef] [Green Version]
- Vogels, G.D.; Van der Drift, C. Degradation of purines and pyrimidines by microorganisms. Bacteriol. Rev. 1976, 40, 403–468. [Google Scholar] [CrossRef] [PubMed]
- Atkins, C.A.; Pate, J.S.; Ritchie, A.; Peoples, M.B. Metabolism and translocation of allantoin in ureide producing grain legumes. Plant Physiol. 1982, 70, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winkler, R.G.; Polacco, J.C.; Blevins, D.G.; Randall, D.D. Enzymatic degradation of allantoate in developing soybeans. Plant Physiol. 1985, 79, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Winkler, R.G.; Blevins, D.G.; Polacco, J.C.; Randall, D.D. Ureide catabolism of soybeans. II. Pathway of catabolism in intact leaf tissue. Plant Physiol. 1987, 83, 585–591. [Google Scholar] [CrossRef] [Green Version]
- Todd, C.D.; Polacco, J.C. Soybean cultivars ‘Williams 82’ and ‘Maple Arrow’ produce both urea and ammonia during ureide degradation. J. Exp. Bot. 2004, 55, 867–877. [Google Scholar] [CrossRef]
- Stebbins, N.E.; Polacco, J.C. Urease is not essential for ureide degradation in soybean. Plant Physiol. 1995, 109, 169–175. [Google Scholar] [CrossRef] [Green Version]
- Werner, A.K.; Medina-Escobar, N.; Zulawski, M.; Sparkes, I.A.; Cao, F.Q.; Witte, C.P. The Ureide-Degrading Reactions of Purine Ring Catabolism Employ Three Amidohydrolases and One Aminohydrolase in Arabidopsis, Soybean, and Rice. Plant Physiol. 2013, 163, 672–681. [Google Scholar] [CrossRef]
- Muñoz, A.; Piedras, P.; Aguilar, M.; Pineda, M. Urea Is a Product of Ureidoglycolate Degradation in Chickpea. Purification and Characterization of the Ureidoglycolate Urea-Lyase. Plant Physiol. 2001, 125, 828–834. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, A.; Raso, M.J.; Pineda, M.; Piedras, P. Degradation of ureidoglycolate in French bean (Phaseolus vulgaris) is catalysed by a ubiquitous ureidoglycolate urea-lyase. Planta 2005, 224, 75–184. [Google Scholar] [CrossRef]
- Muñoz, A.; Bennenberg, G.L.; Montero, O.; Cabello-Díaz, J.M.; Piedras, P.; Pineda, M. An alternative pathway for ureide usage in legumes: Enzymatic formation of a ureidoglycolate adduct in Cicer arietinum and Phaseolus vulgaris. J. Exp. Bot. 2011, 62, 307–318. [Google Scholar] [CrossRef] [Green Version]
- Bohner, A.; Kojima, S.; Hajirezaei, M.; Melzer, M.; von Wirén, N. Urea retranslocation from senescing Arabidopsis leaves is promoted by DUR3-mediated urea retrieval from leaf apoplast. Plant J. 2015, 81, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, N.; Holland, M.A.; Cianzio, S.R.; Polacco, J.C. Genetic tests of the roles of the embryonic ureases of soybean. Plant Physiol. 1991, 97, 1004–1010. [Google Scholar] [CrossRef] [Green Version]
- Blume, C.; Ost, J.; Mühlenbruch, M.; Peterhänsel, C.; Laxa, M. Low CO2 indices urea cycle intermediate accumulation in Arabidopsis thaliana. PLoS ONE 2019, 14, e0210342. [Google Scholar] [CrossRef]
- Yamashita, N.; Tanabata, S.; Ohtake, N.; Sueyoshi, K.; Sato, T.; Higuchi, K. Effects of Different Chemical Forms of Nitrogen on the Quick and Reversible Inhibition of Soybean Nodule Growth and Nitrogen Fixation Activity. Front. Plant Sci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Witte, C.P. Urea metabolism in plants. Plant Sci. 2011, 180, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Real-Guerra, R.; Stanscuaski, F.; Carlini, C.R. Soybean Urease: Over a hundred years of knowledge. In A Comprehensive Surbey of International Soybean Research-Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; InTech: Rijecka, Croatia, 2013; pp. 317–339. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Bothling, L.E.; Polacco, J.C.; Cianzio, S.R. Pleiotropic soybean mutants defective in both urease isozymes. Mol. Gen. Genet. 1987, 209, 432–438. [Google Scholar] [CrossRef]
- Torisky, R.S.; Griffin, J.D.; Yenofsky, R.L.; Polacco, J.C. A single gene (Eu4) encodes the tissue-ubiquitous urease of soybean. Mol. Gen. Ganet. 1994, 242, 404–414. [Google Scholar] [CrossRef]
- Ishikawa, S.; Ono, Y.; Ohtake, N.; Sueyoshi, K.; Tanabata, S.; Ohyama, T. Transcriptome and metabolome analysis reveal that nitrate strongly promotes nitrogen and carbon metabolism in soybean roots, but tends to repress it in nodules. Plants 2018, 7, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, T.A.; Streeter, J.G. Asparagine biosynthesis in soybean nodules. Plant Physiol. 1984, 74, 605–610. [Google Scholar] [CrossRef]
- Minamisawa, K.; Arima, Y.; Kumazawa, K. Characteristics of asparagine pool in soybean nodules in comparison with ureide pool. Soil Sci. Plant Nutr. 1986, 32, 1–14. [Google Scholar] [CrossRef]
- Ohtake, N.; Nishiwaki, T.; Mizukoshi, K.; Minagawa, R.; Takahashi, Y.; Chinushi, T.; Ohyama, T. Amino acid composition in xylem sap of soybean related to the evaluation of N2 fixation by the relative ureide method. Soil Sci. Plant Nutr. 1995, 41, 95–102. [Google Scholar] [CrossRef]
- Sulieman, S.; Tran, L.-S.P. Phosphorus homeostasis in legume nodules as an adaptive strategy to phosphorus deficiency. Plant Sci. 2015, 239, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Rellán-Álvarez, R.; El-Jendoubi, H.; Wohlgemuth, G.; Abadía, A.; Fiehn, O.; Abadía, J.; Álvarez-Fernández, A. Metabolite profile changes in xylem sap and leaf extracts of strategy I plants in response to iron deficiency and resupply. Front. Plant Sci. 2011, 2, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujikake, H.; Yashima, H.; Sato, T.; Ohtake, N.; Sueyoshi, K.; Ohyama, T. Rapid and reversible nitrate inhibition of nodule growth and N2 fixation activity in soybean (Glycine max (L.) Merr.). Soil Sci. Plant Nutr. 2002, 48, 211–217. [Google Scholar] [CrossRef]
- Sakazume, T.; Tanaka, K.; Aida, H.; Ishikawa, S.; Nagumo, Y.; Takahashi, Y. Estimation of nitrogen fixation rate of soybean (Glycine max (L.) Merr.) by micro-scale relative ureide analysis using root bleeding xylem sap and apoplast fluid in stem. Bull. Facul. Agric. Niigata Univ. 2014, 67, 27–41. [Google Scholar]
- Ohyama, T. Analytical Methods for Nitrogen Compounds, Experimental Methods for Plant Nutrition; The Japanese Society of Soil Science and Plant Nutrition: Tokyo, Japan, 1990; pp. 174–203. ISBN 4-8268-0118-1. [Google Scholar]
- Nagumo, Y.; Tanaka, K.; Tewari, K.; Thiraporn, K.; Tsuchida, T.; Honma, T. Rapid quantification of Cyanamide by ultra-high-pressure liquid chromatography in fertilizer, soil or plant samples. J. Chromatogr. A 2009, 1216, 5614–5618. [Google Scholar] [CrossRef]




















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Xylem Sap | Nodules | Roots | Stems | Leaves |
|---|---|---|---|---|---|
| Control | 0.113 | 0.256 | 0.398 | 0.047 | 0.236 |
| Sodium nitrate | 0.246 | 0.154 | 0.458 | 0.07 | 0.212 |
| Ammonium chloride | 0.138 | 0.174 | 0.613 | 0.165 | 0.205 |
| Urea | 0.187 | 0.137 | 0.464 | 0.13 | 0.194 |
| Non-nodulation | 0.292 | 0.158 | 0.11 | 0.164 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ono, Y.; Fukasawa, M.; Sueyoshi, K.; Ohtake, N.; Sato, T.; Tanabata, S.; Toyota, R.; Higuchi, K.; Saito, A.; Ohyama, T. Application of Nitrate, Ammonium, or Urea Changes the Concentrations of Ureides, Urea, Amino Acids and Other Metabolites in Xylem Sap and in the Organs of Soybean Plants (Glycine max (L.) Merr.). Int. J. Mol. Sci. 2021, 22, 4573. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094573
Ono Y, Fukasawa M, Sueyoshi K, Ohtake N, Sato T, Tanabata S, Toyota R, Higuchi K, Saito A, Ohyama T. Application of Nitrate, Ammonium, or Urea Changes the Concentrations of Ureides, Urea, Amino Acids and Other Metabolites in Xylem Sap and in the Organs of Soybean Plants (Glycine max (L.) Merr.). International Journal of Molecular Sciences. 2021; 22(9):4573. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094573
Chicago/Turabian StyleOno, Yuki, Masashige Fukasawa, Kuni Sueyoshi, Norikuni Ohtake, Takashi Sato, Sayuri Tanabata, Ryo Toyota, Kyoko Higuchi, Akihiro Saito, and Takuji Ohyama. 2021. "Application of Nitrate, Ammonium, or Urea Changes the Concentrations of Ureides, Urea, Amino Acids and Other Metabolites in Xylem Sap and in the Organs of Soybean Plants (Glycine max (L.) Merr.)" International Journal of Molecular Sciences 22, no. 9: 4573. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094573