
Metabolic Profiling of VOCs Emitted by Bacteria Isolated from Pressure Ulcers and Treated with Different Concentrations of Bio-AgNPs

 ,
,  , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
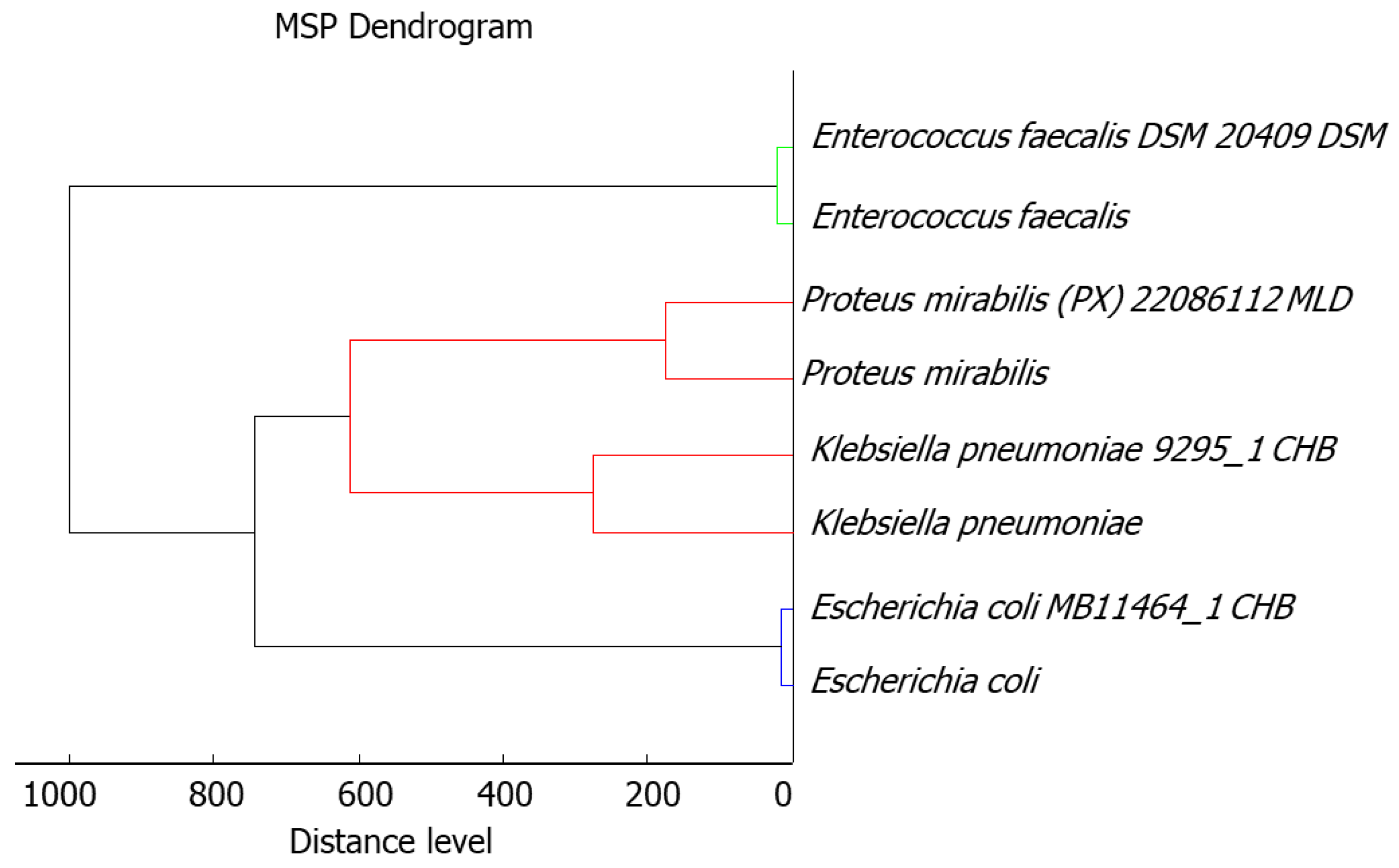
2.1. Bacteria Identification
2.2. Evaluation of Minimum Inhibitory Concentration (MIC)
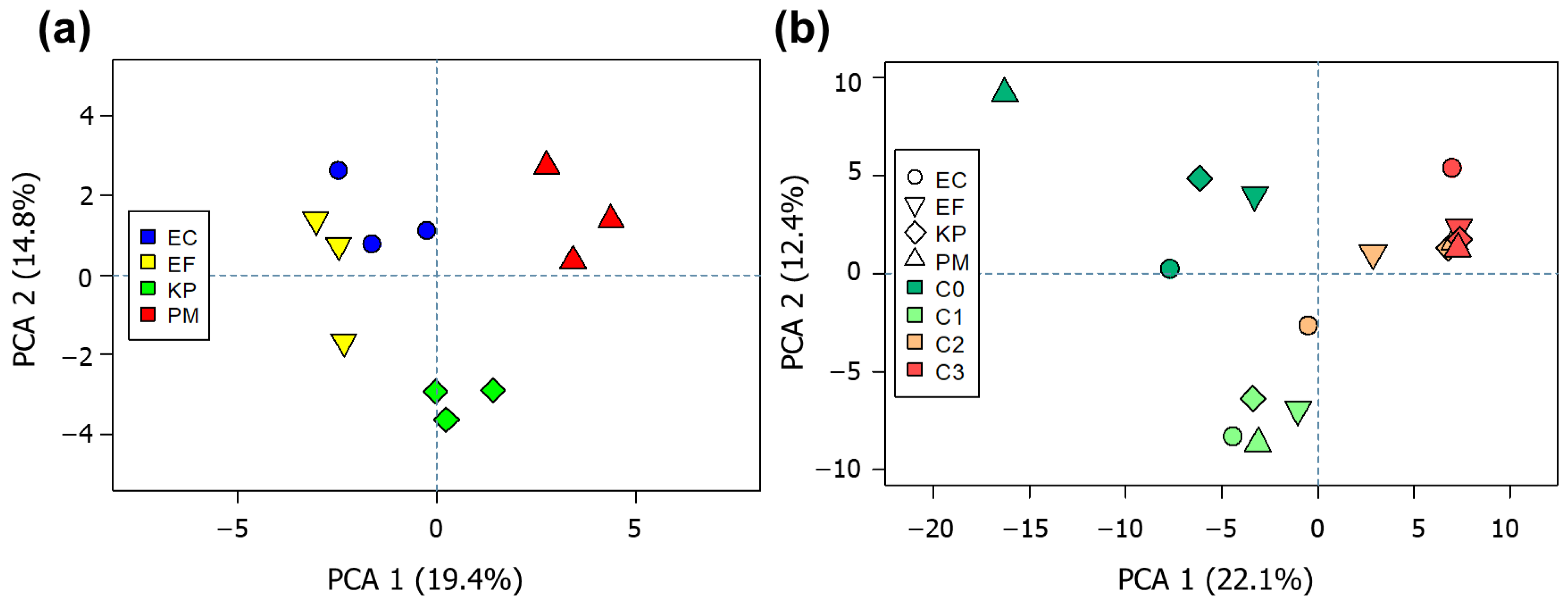
2.3. Unsupervised Inspection of Bacterial VOC Profiles
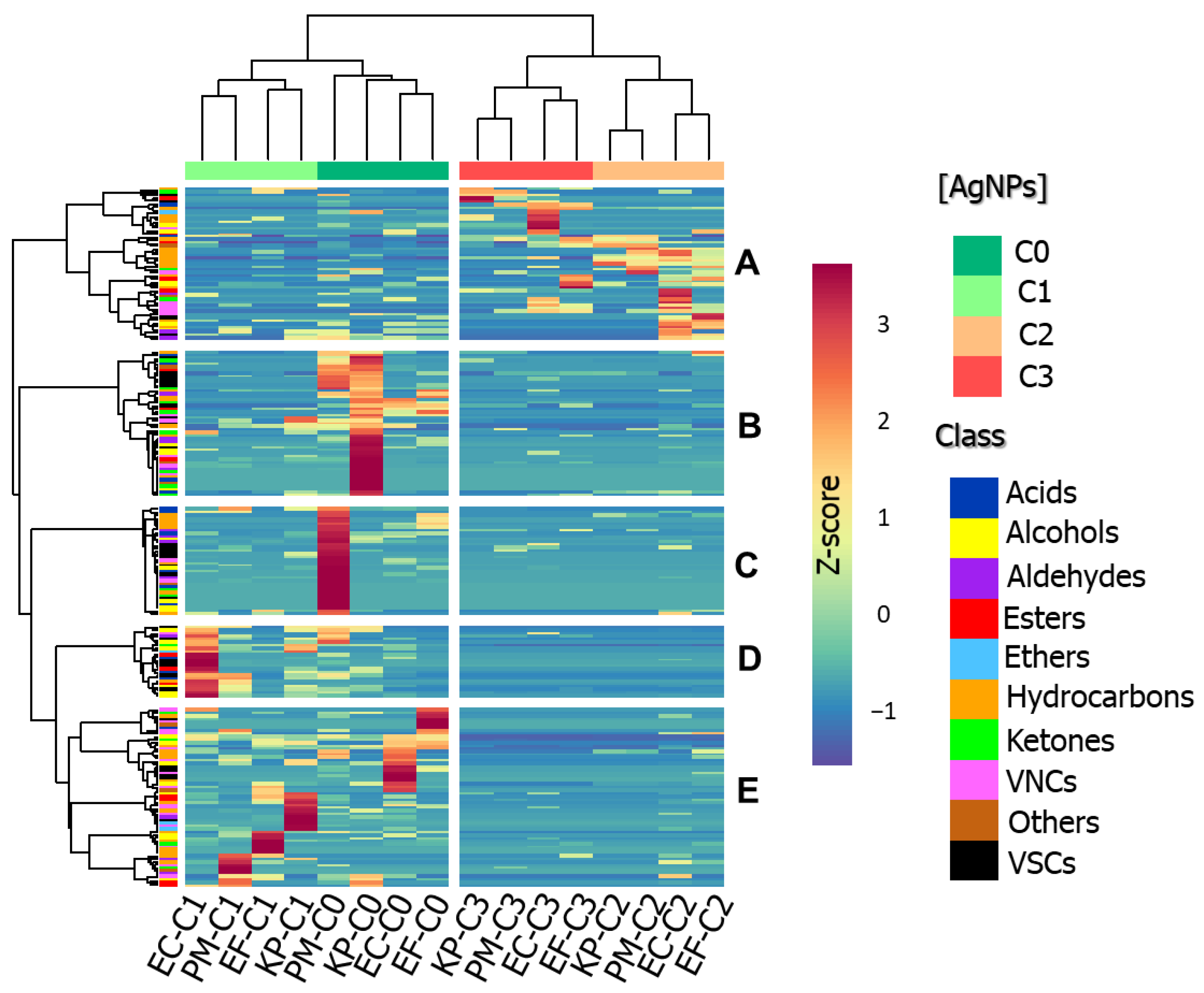
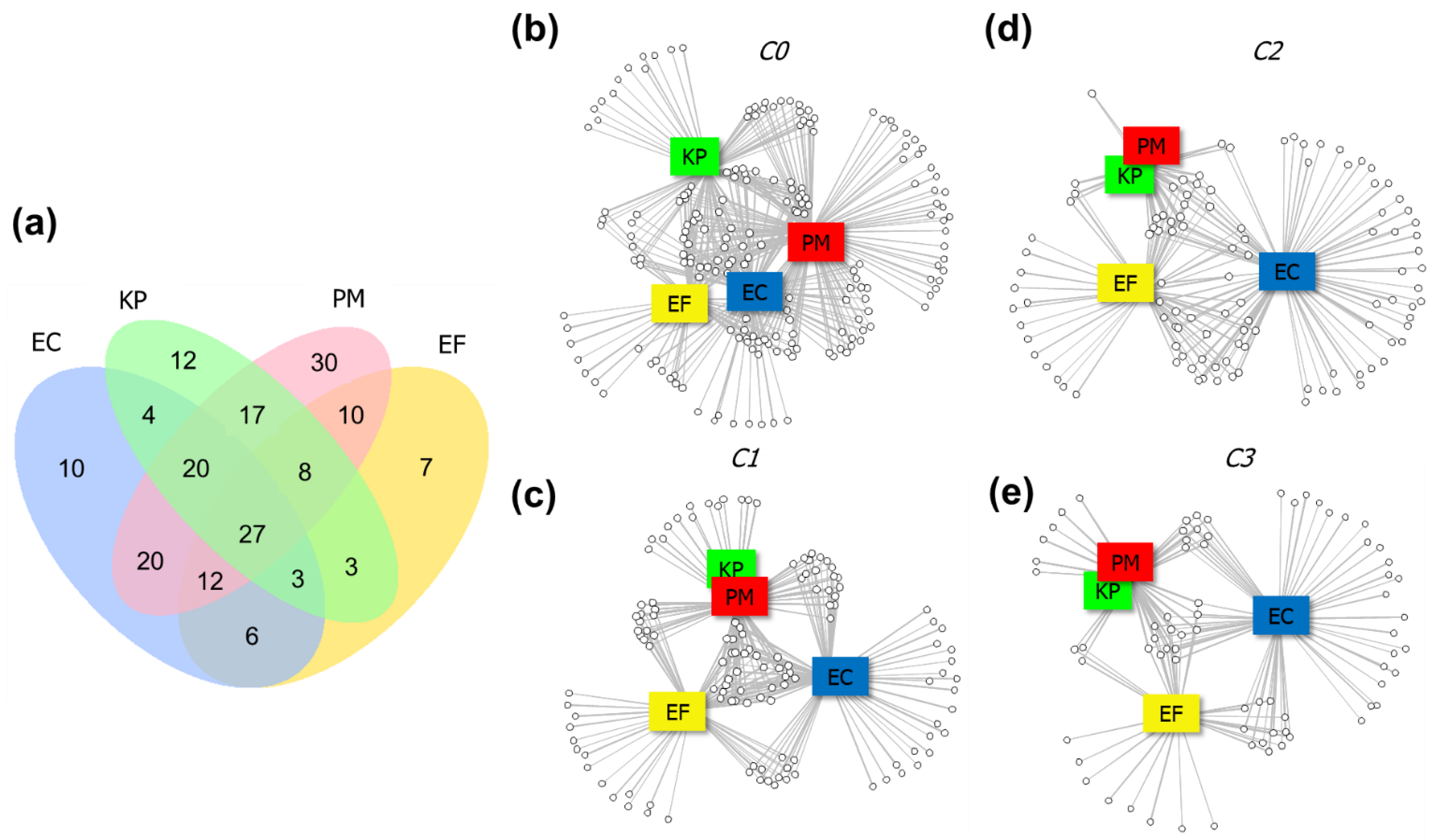
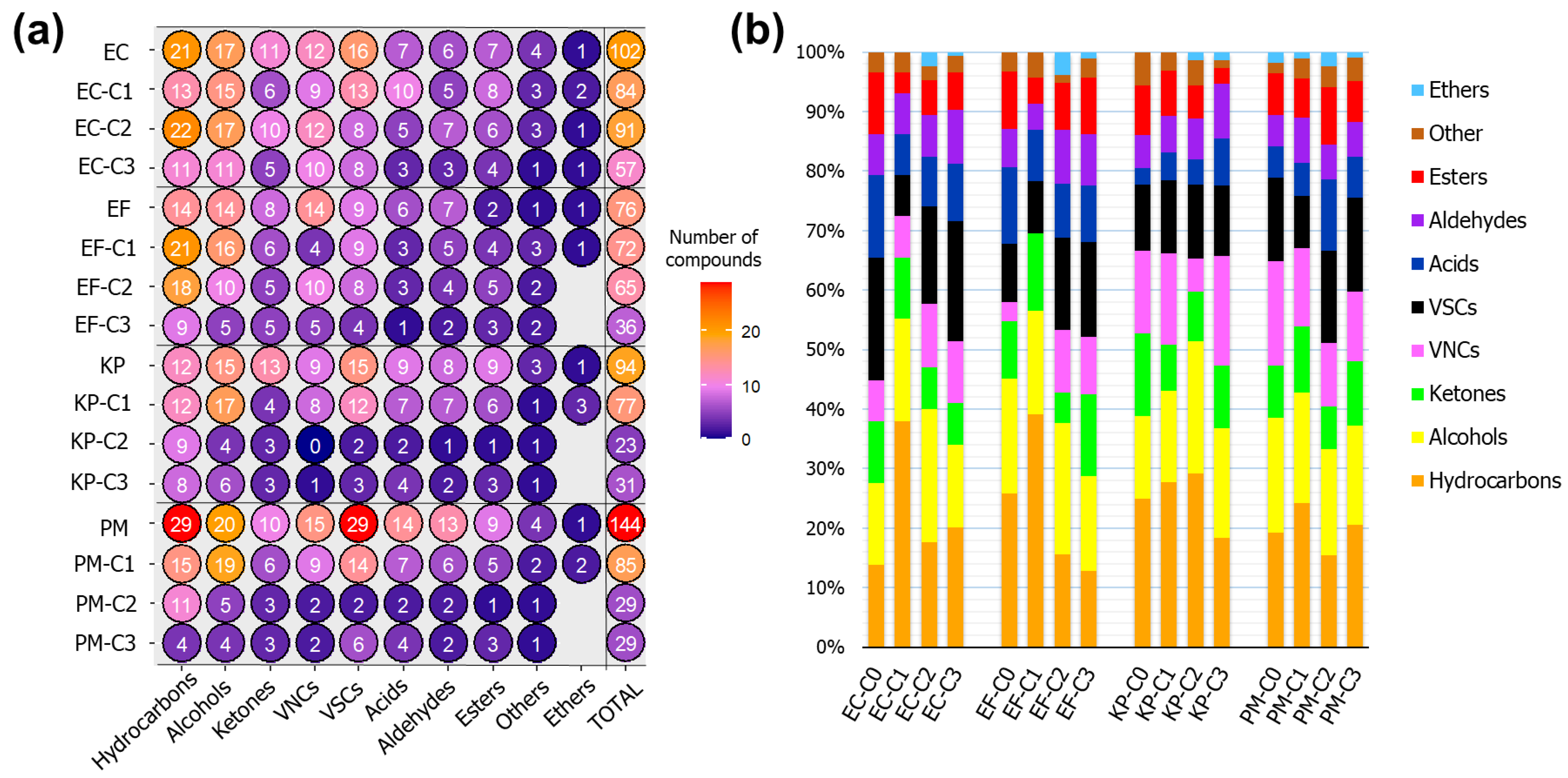
2.4. Differences in VOC Profiles Composition
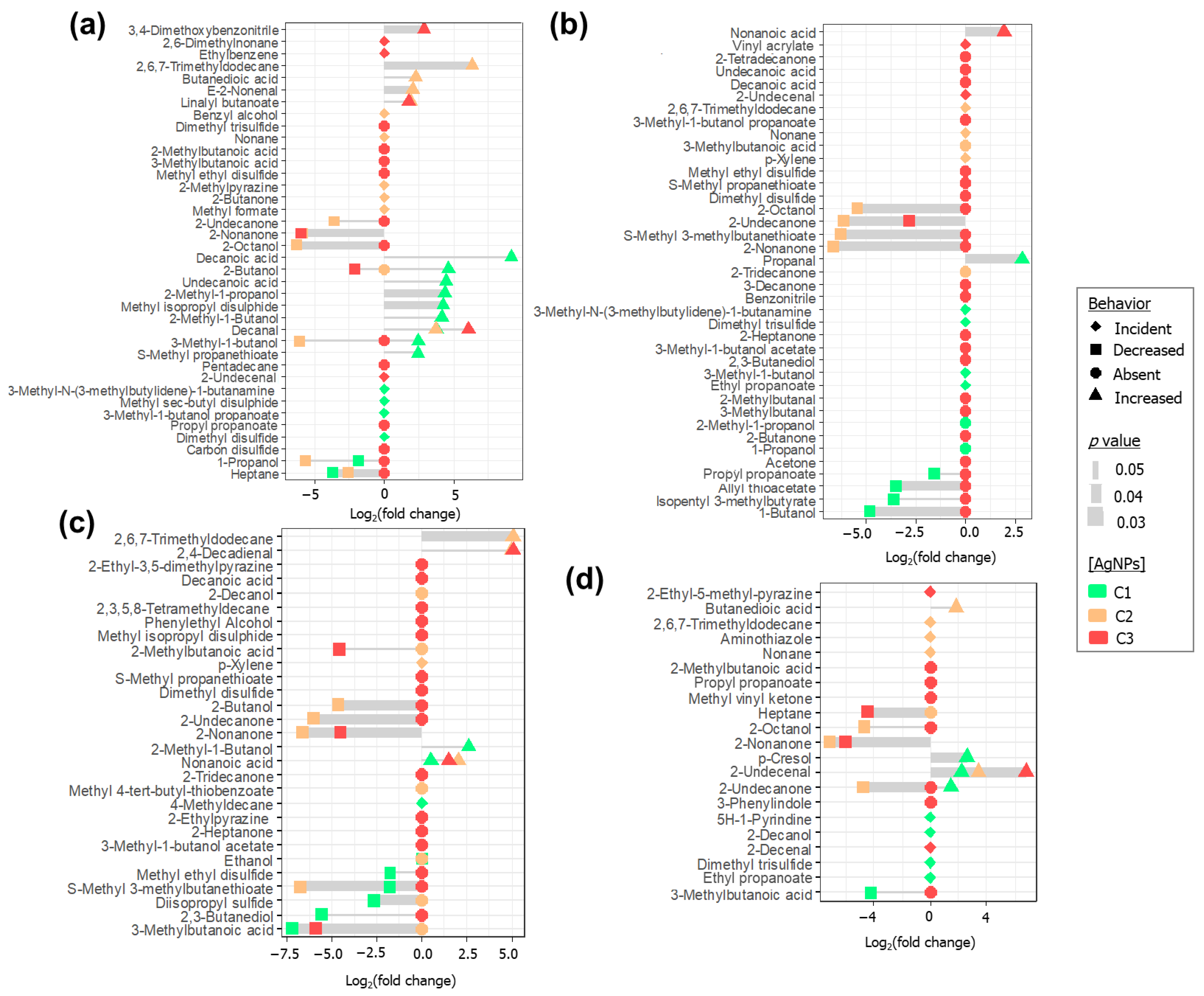
2.5. Investigation of Discriminating Features
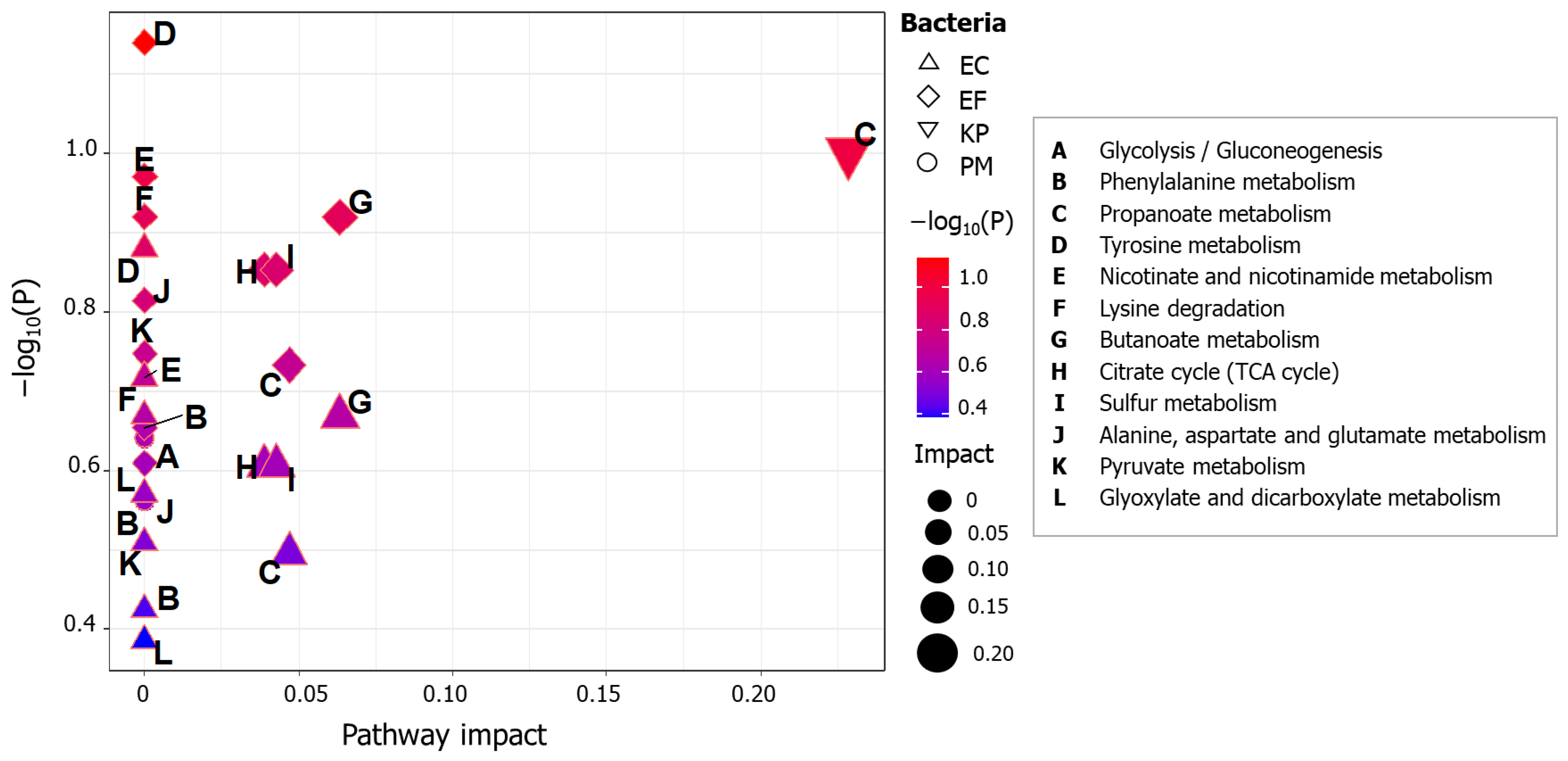
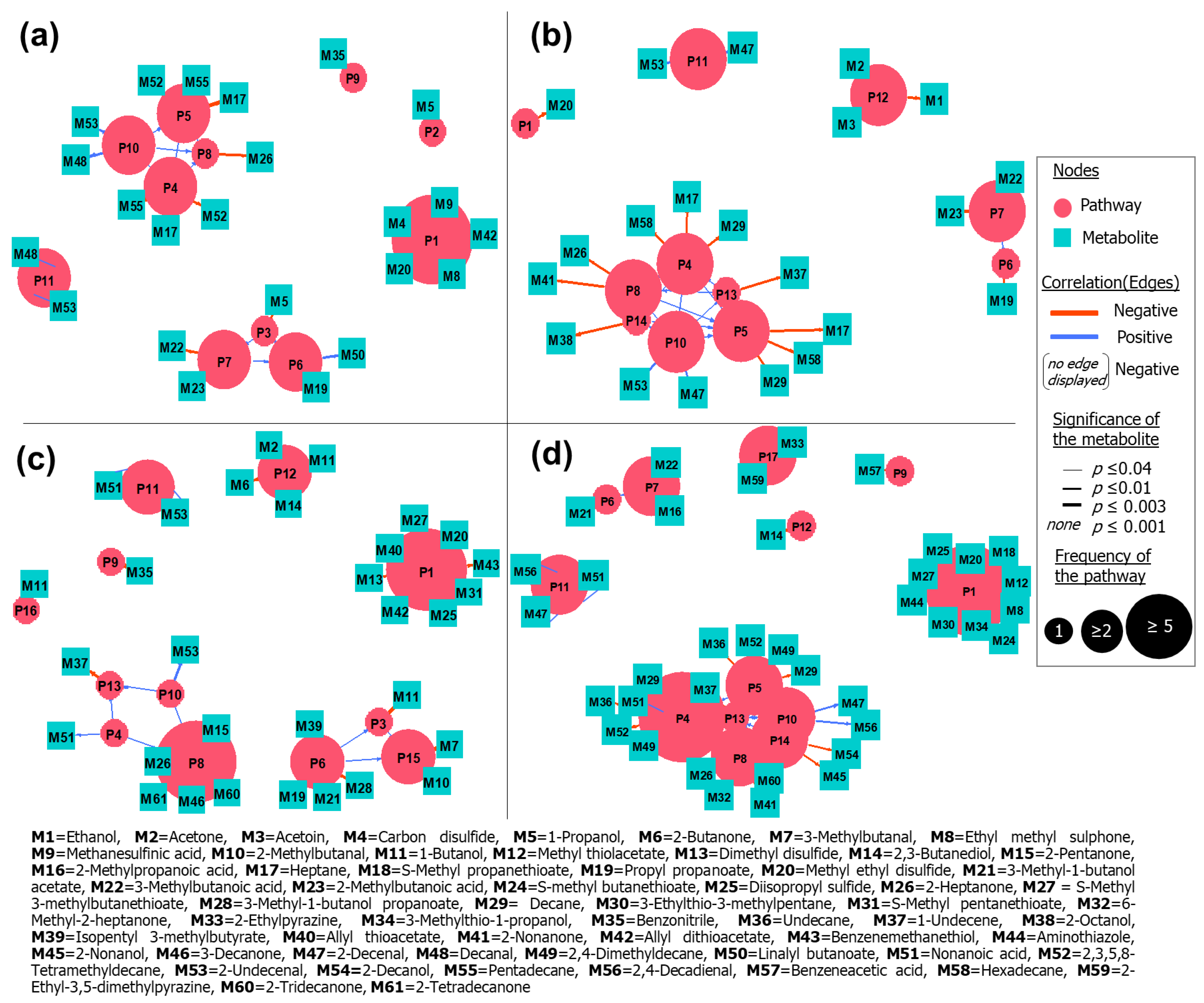
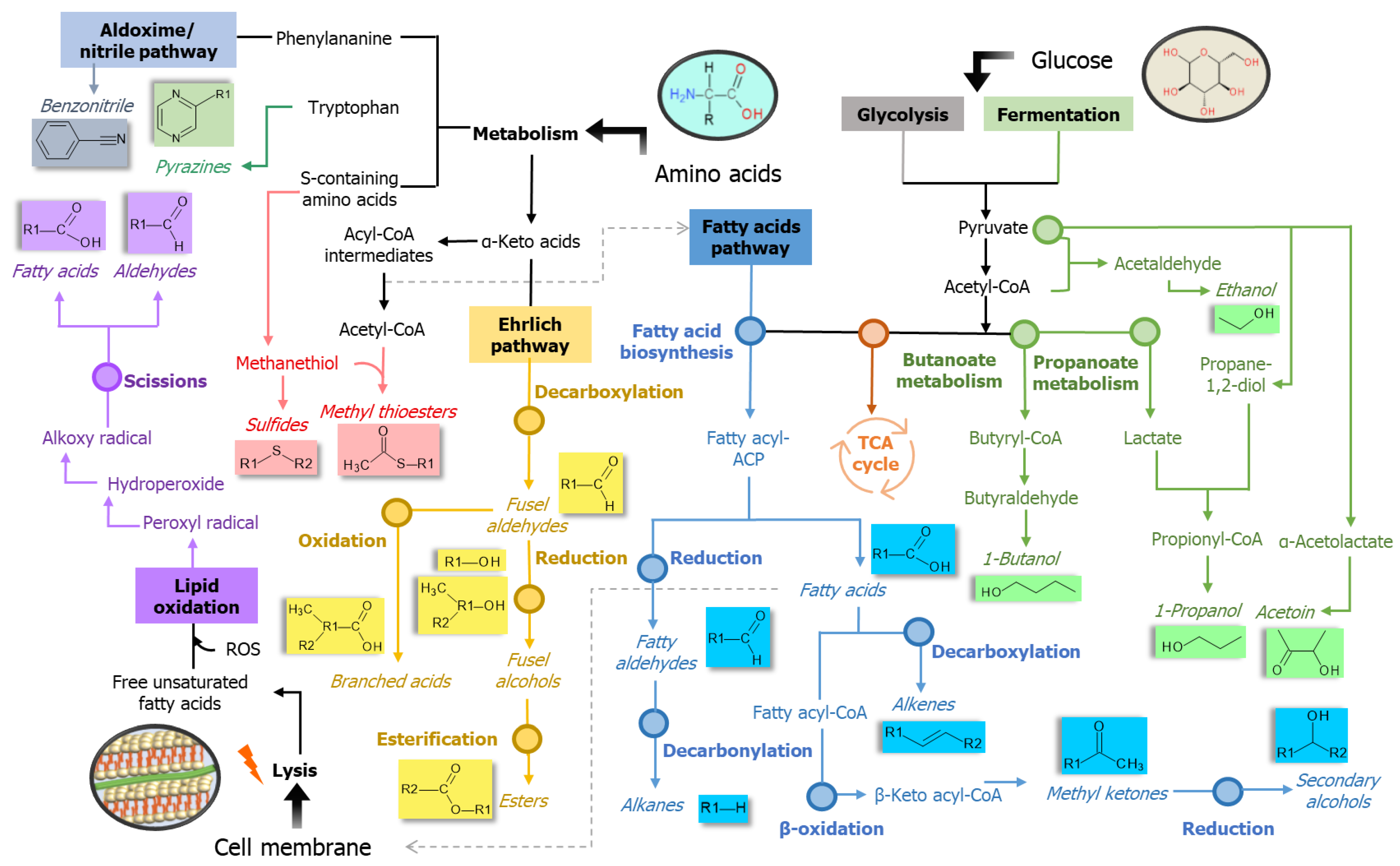
2.6. Ascertainment of Affected Bacterial Pathways
3. Material and Methods
3.1. Sample Preparation
3.1.1. Bacteria Isolation and Identification
3.1.2. Minimal Inhibitory Concentrations (MIC)
3.1.3. Sample Preparation for HS-SPME/GC–MS Analysis
3.2. HS-SPME/GC–MS Analysis
3.3. Statistics and Data Analysis Approaches
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Durán, N.; Durán, M.; de Jesus, M.B.; Seabra, A.B.; Fávaro, W.J.; Nakazato, G. Silver nanoparticles: A new view on mechanistic aspects on antimicrobial activity. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Howell-Jones, R.S.; Wilson, M.J.; Hill, K.E.; Howard, A.J.; Price, P.E.; Thomas, D.W. A review of the microbiology, antibiotic usage and resistance in chronic skin wounds. J. Antimicrob. Chemother. 2005, 55, 143–149. [Google Scholar] [CrossRef] [Green Version]
- Cataldo, M.C.; Bonura, C.; Caputo, G.; Aleo, A.; Rizzo, G.; Geraci, D.M.; Calà, C.; Fasciana, T.; Mattaliano, A.R.; Mammina, C. Colonization of pressure ulcers by multidrug-resistant microorganisms in patients receiving home care. Scand. J. Infect. Dis. 2011, 43, 947–952. [Google Scholar] [CrossRef] [PubMed]
- Rai, M.; Yadav, A.; Gade, A. Silver nanoparticles as a new generation of antimicrobials. Biotechnol. Adv. 2008, 27, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Yin, I.X.; Zhang, J.; Zhao, I.S.; Mei, M.L.; Li, Q.; Chu, C.H. The antibacterial mechanism of silver nanoparticles and its application in dentistry. Int. J. Nanomed. 2020, 15, 2555–2562. [Google Scholar] [CrossRef] [Green Version]
- Salas-Orozco, M.; Niño-Martínez, N.; Martínez-Castañón, G.-A.; Méndez, F.T.; Jasso, M.E.C.; Ruiz, F. Mechanisms of Resistance to Silver Nanoparticles in Endodontic Bacteria: A Literature Review. J. Nanomater. 2019, 2019, 7630316. [Google Scholar] [CrossRef]
- Dakal, T.C.; Kumar, A.; Majumdar, R.S.; Yadav, V. Mechanistic basis of antimicrobial actions of silver nanoparticles. Front. Microbiol. 2016, 7, 1831. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, S.; Poulose, E.K. Silver nanoparticles: Mechanism of antimicrobial action, synthesis, medical applications, and toxicity effects. Int. Nano Lett. 2012, 2, 32. [Google Scholar] [CrossRef] [Green Version]
- Gumel, A.M.; Surayya, M.M.; Yaro, M.N.; Waziri, I.Z.; Amina, A.A. Biogenic synthesis of silver nanoparticles and its synergistic antimicrobial potency: An overview. J. Appl. Biotechnol. Bioeng. 2019, 6, 22–28. [Google Scholar] [CrossRef]
- Sondi, I.; Salopek-Sondi, B. Silver nanoparticles as antimicrobial agent: A case study on E. coli as a model for Gram-negative bacteria. J. Colloid Interface Sci. 2004, 275, 177–182. [Google Scholar] [CrossRef]
- Morones, J.R.; Elechiguerra, J.L.; Camacho, A.; Holt, K.; Kouri, J.B.; Ramírez, J.T.; Yacaman, M.J. The bactericidal effect of silver nanoparticles. Nanotechnology 2005, 16, 2346–2353. [Google Scholar] [CrossRef] [Green Version]
- Shrivastava, S.; Bera, T.; Roy, A.; Singh, G.; Ramachandrarao, P.; Dash, D. Characterization of enhanced antibacterial effects of novel silver nanoparticles. Nanotechnology 2007, 18, 225103. [Google Scholar] [CrossRef]
- Singh, J.; Dutta, T.; Kim, K.-H.; Rawat, M.; Samddar, P.; Kumar, P. ‘Green’ synthesis of metals and their oxide nanoparticles: Applications for environmental remediation. J. Nanobiotechnol. 2018, 16, 84. [Google Scholar] [CrossRef]
- Shi, Y.; Liang, B.; Alsalme, A.; Lin, R.-B.; Chen, B. Mechanochemical synthesis of an ethylene sieve UTSA-280. J. Solid State Chem. 2020, 287, 121321. [Google Scholar] [CrossRef]
- Railean-Plugaru, V.; Pomastowski, P.; Rafinska, K.; Wypij, M.; Kupczyk, W.; Dahm, H.; Jackowski, M.; Buszewski, B. Antimicrobial properties of biosynthesized silver nanoparticles studied by flow cytometry and related techniques. Electrophoresis 2016, 37, 752–761. [Google Scholar] [CrossRef]
- Kubasheva, Z.; Sprynskyy, M.; Railean-Plugaru, V.; Pomastowski, P.; Ospanova, A.; Buszewski, B. Synthesis and Antibacterial Activity of (AgCl, Ag)NPs/Diatomite Hybrid Composite. Materials 2020, 13, 3409. [Google Scholar] [CrossRef] [PubMed]
- Clish, C.B. Metabolomics: An emerging but powerful tool for precision medicine. Mol. Case Stud. 2015, 1, a000588. [Google Scholar] [CrossRef] [Green Version]
- Schulz, S.; Dickschat, J.S. Bacterial volatiles: The smell of small organisms. Nat. Prod. Rep. 2007, 24, 814–842. [Google Scholar] [CrossRef]
- Milanowski, M.; Monedeiro, F.; Złoch, M.; Ratiu, I.-A.; Pomastowski, P.; Ligor, T.; De Martinis, B.S.; Buszewski, B. Profiling of VOCs released from different salivary bacteria treated with non-lethal concentrations of silver nitrate. Anal. Biochem. 2019, 578, 36–44. [Google Scholar] [CrossRef]
- Buszewski, B.; Rațiu, I.A.; Milanowski, M.; Pomastowski, P.; Ligor, T. The effect of biosilver nanoparticles on different bacterial strains’ metabolism reflected in their VOCs profiles. J. Breath Res. 2018, 12, 027105. [Google Scholar] [CrossRef] [Green Version]
- Monedeiro, F.; Pomastowski, P.; Milanowski, M.; Ligor, T.; Buszewski, B. Monitoring of Bactericidal Effects of Silver Nanoparticles Based on Protein Signatures and VOC Emissions from Escherichia coli and Selected Salivary Bacteria. J. Clin. Med. 2019, 8, 2024. [Google Scholar] [CrossRef] [Green Version]
- Dana, A.N.; Bauman, W.A. Bacteriology of pressure ulcers in individuals with spinal cord injury: What we know and what we should know. J. Spinal Cord Med. 2015, 38, 147–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braga, I.A.; Brito, C.S.; Filho, A.D.; Filho, P.P.G.; Ribas, R.M. Pressure ulcer as a reservoir of multiresistant Gram-negative bacilli: Risk factors for colonization and development of bacteremia. Braz. J. Infect. Dis. 2017, 21, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Godebo, G.; Kibru, G.; Tassew, H. Multidrug-resistant bacterial isolates in infected wounds at Jimma University Specialized Hospital, Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 17. [Google Scholar] [CrossRef] [Green Version]
- Esmail, M.A.M.; Abdulghany, H.M.; Khairy, R.M. Prevalence of Multidrug-Resistant Enterococcus faecalis in Hospital-Acquired Surgical Wound Infections and Bacteremia: Concomitant Analysis of Antimicrobial Resistance Genes. Infect. Dis. Res. Treat. 2019, 12, 1178633719882929. [Google Scholar] [CrossRef]
- Buszewski, B.; Railean-Plugaru, V.; Pomastowski, P.; Rafinska, K.; Szultka-Mlynska, M.; Kowalkowski, T. Antimicrobial Effectiveness of Bioactive Silver Nanoparticles Synthesized by Actinomycetes HGG16n Strain. Curr. Pharm. Biotechnol. 2017, 18, 168–176. [Google Scholar] [CrossRef] [PubMed]
- Railean-Plugaru, V.; Pomastowski, P.; Meller, K.; Złoch, M.; Rafinska, K.; Buszewski, B. Lactococcus lactis as a safe and inexpensive source of bioactive silver composites. Appl. Microbiol. Biotechnol. 2017, 101, 7141–7153. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Lu, X. Microbial Synthesis of Alka(e)nes. Front. Bioeng. Biotechnol. 2013, 1, 10. [Google Scholar] [CrossRef] [Green Version]
- Ladygina, N.; Dedyukhina, E.G.; Vainshtein, M.B. A review on microbial synthesis of hydrocarbons. Process Biochem. 2006, 41, 1001–1014. [Google Scholar] [CrossRef]
- Schaich, K.M.; Shahidi, F.; Zhong, Y.; Eskin, N.A.M. Lipid Oxidation. In Biochemistry of Foods; Eskin, N.A., Shahidi, F., Eds.; Elsevier: Cambridge, UK, 2013; pp. 419–478. ISBN 9780122423529. [Google Scholar]
- Hachicho, N.; Hoffmann, P.; Ahlert, K.; Heipieper, H.J. Effect of silver nanoparticles and silver ions on growth and adaptive response mechanisms of Pseudomonas putida mt-2. FEMS Microbiol. Lett. 2014, 355, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anaya, N.M.; Faghihzadeh, F.; Ganji, N.; Bothun, G.; Oyanedel-Craver, V. Comparative study between chemostat and batch reactors to quantify membrane permeability changes on bacteria exposed to silver nanoparticles. Sci. Total Environ. 2015, 565, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Zeng, J.; Wang, X.; Drlica, K.; Zhao, X. Post-stress bacterial cell death mediated by reactive oxygen species. Proc. Natl. Acad. Sci. USA 2019, 116, 10064–10071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.; Drlica, K. Reactive oxygen species and the bacterial response to lethal stress. Curr. Opin. Microbiol. 2014, 21, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becerra, M.C.; Páez, P.L.; Laróvere, L.E.; Albesa, I. Lipids and DNA oxidation in Staphylococcus aureus as a consequence of oxidative stress generated by ciprofloxacin. Mol. Cell. Biochem. 2006, 285, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Guerzoni, M.E.; Lanciotti, R.; Cocconcelli, P.S. Alteration in cellular fatty acid composition as a response to salt, acid, oxidative and thermal stresses in Lactobacillus helveticus. Microbiology 2001, 147, 2255–2264. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.T.; Zheng, X.; Li, S. Tea aroma formation. Food Sci. Hum. Wellness 2015, 4, 9–27. [Google Scholar] [CrossRef] [Green Version]
- Groenhagen, U.; Baumgartner, R.; Bailly, A.; Gardiner, A.; Eberl, L.; Schulz, S.; Weisskopf, L. Production of Bioactive Volatiles by Different Burkholderia ambifaria Strains. J. Chem. Ecol. 2013, 39, 892–906. [Google Scholar] [CrossRef] [PubMed]
- Audrain, B.; Farag, M.A.; Ryu, C.-M.; Ghigo, J.-M. Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 2015, 39, 222–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atsumi, S.; Hanai, T.; Liao, J.C. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels. Nature 2008, 451, 86–89. [Google Scholar] [CrossRef]
- Marilley, L.; Casey, M.G. Flavours of cheese products: Metabolic pathways, analytical tools and identification of producing strains. Int. J. Food Microbiol. 2004, 90, 139–159. [Google Scholar] [CrossRef]
- Menendez-Bravo, S.; Comba, S.; Gramajo, H.; Arabolaza, A. Metabolic engineering of microorganisms for the production of structurally diverse esters. Appl. Microbiol. Biotechnol. 2017, 101, 3043–3053. [Google Scholar] [CrossRef]
- Tai, Y.S.; Xiong, M.; Zhang, K. Engineered biosynthesis of medium-chain esters in Escherichia coli. Metab. Eng. 2015, 27, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Redelman, C.V.; Maduakolam, C.; Anderson, G.G. Alcohol treatment enhances Staphylococcus aureus biofilm development. FEMS Immunol. Med. Microbiol. 2012, 66, 411–418. [Google Scholar] [CrossRef] [Green Version]
- Saxena, R.K.; Anand, P.; Saran, S.; Isar, J.; Agarwal, L. Microbial production and applications of 1,2-propanediol. Indian J. Microbiol. 2010, 50, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Toraya, T.; Honda, S.; Fukui, S. Fermentation of 1,2-Propanediol and 1,2-Ethanediol by Some Genera of Enterobacteriaceae, Involving Coenzyme B12-Dependent Diol Dehydratase. J. Bacteriol. 1979, 139, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Russmayer, H.; Marx, H.; Sauer, M. Microbial 2-butanol production with Lactobacillus diolivorans. Biotechnol. Biofuels 2019, 12, 262. [Google Scholar] [CrossRef] [PubMed]
- Dawson, L.F.; Stabler, R.A.; Wren, B.W. Assessing the role of p-cresol tolerance in Clostridium difficile. J. Med. Microbiol. 2008, 57, 745–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Passmore, I.J.; Letertre, M.P.M.; Preston, M.D.; Bianconi, I.; Harrison, M.A.; Nasher, F.; Kaur, H.; Hong, H.A.; Baines, S.D.; Cutting, S.M.; et al. Para-cresol production by Clostridium difficile affects microbial diversity and membrane integrity of Gram-negative bacteria. PLoS Pathog. 2018, 14, e1007191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutanovas, S.; Rutkienė, R.; Urbelis, G.; Tauraitė, D.; Stankevičiūtė, J.; Meškys, R. Bioconversion of methylpyrazines and pyridines using novel pyrazines-degrading microorganisms. CHEMIJA 2013, 24, 67–73. [Google Scholar]
- Stepanić, V.; Matijašić, M.; Horvat, T.; Verbanac, D.; Chlupáćová, M.K.; Saso, L.; Žarković, N. Antioxidant activities of alkyl substituted pyrazine derivatives of chalcones—In vitro and in silico study. Antioxidants 2019, 8, 90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Létoffé, S.; Audrain, B.; Bernier, S.P.; Delepierre, M.; Ghigo, J.M. Aerial exposure to the bacterial volatile compound trimethylamine modifies antibiotic resistance of physically separated bacteria by raising culture medium pH. MBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Van Der Werf, M.J.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Environmental and physiological factors affecting the succinate product ratio during carbohydrate fermentation by Actinobacillus sp. 130Z. Arch. Microbiol. 1997, 167, 332–342. [Google Scholar] [CrossRef]
- Milisavljevic, V.; Tran, L.P.; Batmalle, C.; Bootsma, H.J. Benzyl alcohol and ethanol can enhance the pathogenic potential of clinical Staphylococcus epidermidis strains. Am. J. Infect. Control 2008, 36, 552–558. [Google Scholar] [CrossRef]
- Xiu, Z.; Zhang, Q.; Puppala, H.L.; Colvin, V.L.; Alvarez, P.J.J. Negligible Particle-Specific Antibacterial Activity of Silver Nanoparticles. Nano Lett. 2012, 12, 4271–4275. [Google Scholar] [CrossRef] [PubMed]
- Veronesi, G.; Deniaud, A.; Gallon, T.; Jouneau, P.-H.; Villanova, J.; Delangle, P.; Carrière, M.; Kieffer, I.; Charbonnier, P.; Mintz, E.; et al. Visualization, quantification and coordination of Ag + ions released from silver nanoparticles in hepatocytes. Nanoscale 2016, 8, 17012–17021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahearn, D.G.; May, L.L.; Gabriel, M.M. Adherence of organisms to silver-coated surfaces. J. Ind. Microbiol. 1995, 15, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, W.J.A.; Rosenberg, H. Effect of Silver Ions on Transport and Retention of Phosphate by Escherichia coli. J. Bacteriol. 1982, 152, 7–13. [Google Scholar] [PubMed]
- Haze, S.; Gozu, Y.; Nakamura, S.; Kohno, Y.; Sawano, K.; Ohta, H.; Yamazaki, K. 2-Nonenal Newly Found in Human Body Odor Tends to Increase with Aging. J. Investig. Dermatol. 2001, 116, 520–524. [Google Scholar] [CrossRef] [Green Version]
- Gosset, G. Production of aromatic compounds in bacteria. Curr. Opin. Biotechnol. 2009, 20, 651–658. [Google Scholar] [CrossRef]
- Orlov, I.A.; Sankova, T.P.; Babich, P.S.; Sosnin, I.M.; Ilyechova, E.Y.; Kirilenko, D.A.; Brunkov, P.N.; Ataev, G.L.; Romanov, A.E.; Puchkova, L.V. New silver nanoparticles induce apoptosis-like process in E. coli and interfere with mammalian copper metabolism. Int. J. Nanomed. 2016, 11, 6561–6574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bondarenko, O.M.; Sihtmäe, M.; Kuzmičiova, J.; Ragelienė, L.; Kahru, A.; Daugelavičius, R. Plasma membrane is the target of rapid antibacterial action of silver nanoparticles in escherichia coli and Pseudomonas aeruginosa. Int. J. Nanomed. 2018, 13, 6779–6790. [Google Scholar] [CrossRef] [Green Version]
- Komagata, K.; Suzuki, K.I. Lipid and Cell-Wall Analysis in Bacterial Systematics. In Methods in Microbiology; Colwell, R.R., Grigorova, R., Eds.; Academic Press: Orlando, FL, USA, 1987; Volume 19, pp. 161–207. [Google Scholar]
- Prion, S.; Haerling, K.A. Making Sense of Methods and Measurement: Spearman-Rho Ranked-Order Correlation Coefficient. Clin. Simul. Nurs. 2014, 10, 535–536. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Rock, C.O. Membrane lipid homeostasis in bacteria. Nat. Rev. Microbiol. 2008, 6, 222–233. [Google Scholar] [CrossRef]
- Monedeiro, F.; Monedeiro-Milanowski, M.; Ligor, T.; Buszewski, B. A Review of GC-Based Analysis of Non-Invasive Biomarkers of Colorectal Cancer and Related Pathways. J. Clin. Med. 2020, 9, 3191. [Google Scholar] [CrossRef]
- Ravindran, A.; Chandrasekaran, N.; Mukherjee, A. Studies on Differential Behavior of Silver Nanoparticles Towards Thiol Containing Amino Acids. Curr. Nanosci. 2012, 8, 141–149. [Google Scholar] [CrossRef]
- Michaeloudes, C.; Seiffert, J.; Chen, S.; Ruenraroengsak, P.; Bey, L.; Theodorou, I.G.; Ryan, M.; Cui, X.; Zhang, J.; Shaffer, M.; et al. Effect of silver nanospheres and nanowires on human airway smooth muscle cells: Role of sulfidation. Nanoscale Adv. 2020, 2, 5635–5647. [Google Scholar] [CrossRef]
- Forney, F.W.; Markovetz, A.J. The biology of methyl ketones. J. Lipid Res. 1971, 12, 383–395. [Google Scholar] [CrossRef]
- Aittokallio, T. Graph-based methods for analysing networks in cell biology. Brief. Bioinform. 2006, 7, 243–255. [Google Scholar] [CrossRef]
- Bhalla, T.C.; Kumar, V.; Kumar, V. Enzymes of aldoxime–nitrile pathway for organic synthesis. Rev. Environ. Sci. Biotechnol. 2018, 17, 229–239. [Google Scholar] [CrossRef]
- Rudakiya, D.M.; Pawar, K. Bactericidal potential of silver nanoparticles synthesized using cell-free extract of Comamonas acidovorans: In vitro and in silico approaches. 3 Biotech 2017, 7, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Fayaz, A.M.; Balaji, K.; Girilal, M.; Yadav, R.; Kalaichelvan, P.T.; Venketesan, R. Biogenic synthesis of silver nanoparticles and their synergistic effect with antibiotics: A study against gram-positive and gram-negative bacteria. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Panáček, A.; Kvítek, L.; Smékalová, M.; Večeřová, R.; Kolář, M.; Röderová, M.; Dyčka, F.; Šebela, M.; Prucek, R.; Tomanec, O.; et al. Bacterial resistance to silver nanoparticles and how to overcome it. Nat. Nanotechnol. 2018, 13, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Gunawan, C.; Teoh, W.Y.; Marquis, C.P.; Amal, R. Induced Adaptation of Bacillus sp. to Antimicrobial Nanosilver. Small 2013, 9, 3554–3560. [Google Scholar] [CrossRef] [PubMed]
- Graves, J.L.; Tajkarimi, M.; Cunningham, Q.; Campbell, A.; Nonga, H.; Harrison, S.H.; Barrick, J.E. Rapid evolution of silver nanoparticle resistance in Escherichia coli. Front. Genet. 2015, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.L.; Zhu, D.; An, X.L.; Ding, J.; Zhu, Y.G.; Cui, L. Does nano silver promote the selection of antibiotic resistance genes in soil and plant? Environ. Int. 2019, 128, 399–406. [Google Scholar] [CrossRef]
- Pareek, V.; Devineau, S.; Sivasankaran, S.K.; Bhargava, A.; Panwar, J.; Srikumar, S.; Fanning, S. Silver Nanoparticles Induce a Triclosan-Like Antibacterial Action Mechanism in Multi-Drug Resistant Klebsiella pneumoniae. Front. Microbiol. 2021, 12, 638640. [Google Scholar] [CrossRef]
- Parveen, A.; Yalagatti, M.S.; Abbaraju, V.; Deshpande, R. Emphasized Mechanistic Antimicrobial Study of Biofunctionalized Silver Nanoparticles on Model Proteus mirabilis. J. Drug Deliv. 2018, 2018, 3850139. [Google Scholar] [CrossRef] [Green Version]
- Saeb, A.T.M.; Al-Rubeaan, K.A.; Abouelhoda, M.; Selvaraju, M.; Tayeb, H.T. Genome sequencing and analysis of the first spontaneous Nanosilver resistant bacterium Proteus mirabilis strain SCDR1. Antimicrob. Resist. Infect. Control 2017, 6, 119. [Google Scholar] [CrossRef] [Green Version]
- Baumler, D.J.; Hung, K.F.; Jeong, K.C.; Kaspar, C.W. Production of methanethiol and volatile sulfur compounds by the archaeon “Ferroplasma acidarmanus”. Extremophiles 2007, 11, 841–851. [Google Scholar] [CrossRef]
- Miranda, R.R.; Gorshkov, V.; Korzeniowska, B.; Kempf, S.J.; Neto, F.F.; Kjeldsen, F. Co-exposure to silver nanoparticles and cadmium induce metabolic adaptation in HepG2 cells. Nanotoxicology 2018, 12, 781–795. [Google Scholar] [CrossRef] [Green Version]
- Buszewski, B.; Pomastowski, P.; Rafińska, K.; Railean-Plugaru, V. Izolacja Nowych Szczepów Bakterii Kwasu Mlekowego i Sposób Otrzymywania Nanocząsteczek za Pomocą Tych Szczepów Oraz Lecznicze Zastosowanie Takich Nanocząstek Srebra. P.418480. 2016. Available online: https://ewyszukiwarka.pue.uprp.gov.pl/search/pwp-details/P.418480?lng=en (accessed on 28 April 2020).
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanehisa, M.; Goto, S. KEGG: Kyoto Encyclopedia of Genes and Genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monedeiro, F.; Railean-Plugaru, V.; Monedeiro-Milanowski, M.; Pomastowski, P.; Buszewski, B. Metabolic Profiling of VOCs Emitted by Bacteria Isolated from Pressure Ulcers and Treated with Different Concentrations of Bio-AgNPs. Int. J. Mol. Sci. 2021, 22, 4696. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094696
Monedeiro F, Railean-Plugaru V, Monedeiro-Milanowski M, Pomastowski P, Buszewski B. Metabolic Profiling of VOCs Emitted by Bacteria Isolated from Pressure Ulcers and Treated with Different Concentrations of Bio-AgNPs. International Journal of Molecular Sciences. 2021; 22(9):4696. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094696
Chicago/Turabian StyleMonedeiro, Fernanda, Viorica Railean-Plugaru, Maciej Monedeiro-Milanowski, Paweł Pomastowski, and Bogusław Buszewski. 2021. "Metabolic Profiling of VOCs Emitted by Bacteria Isolated from Pressure Ulcers and Treated with Different Concentrations of Bio-AgNPs" International Journal of Molecular Sciences 22, no. 9: 4696. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094696