
Hypergravity Load Modulates Acetaminophen Nephrotoxicity via Endoplasmic Reticulum Stress in Association with Hepatic microRNA-122 Expression


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
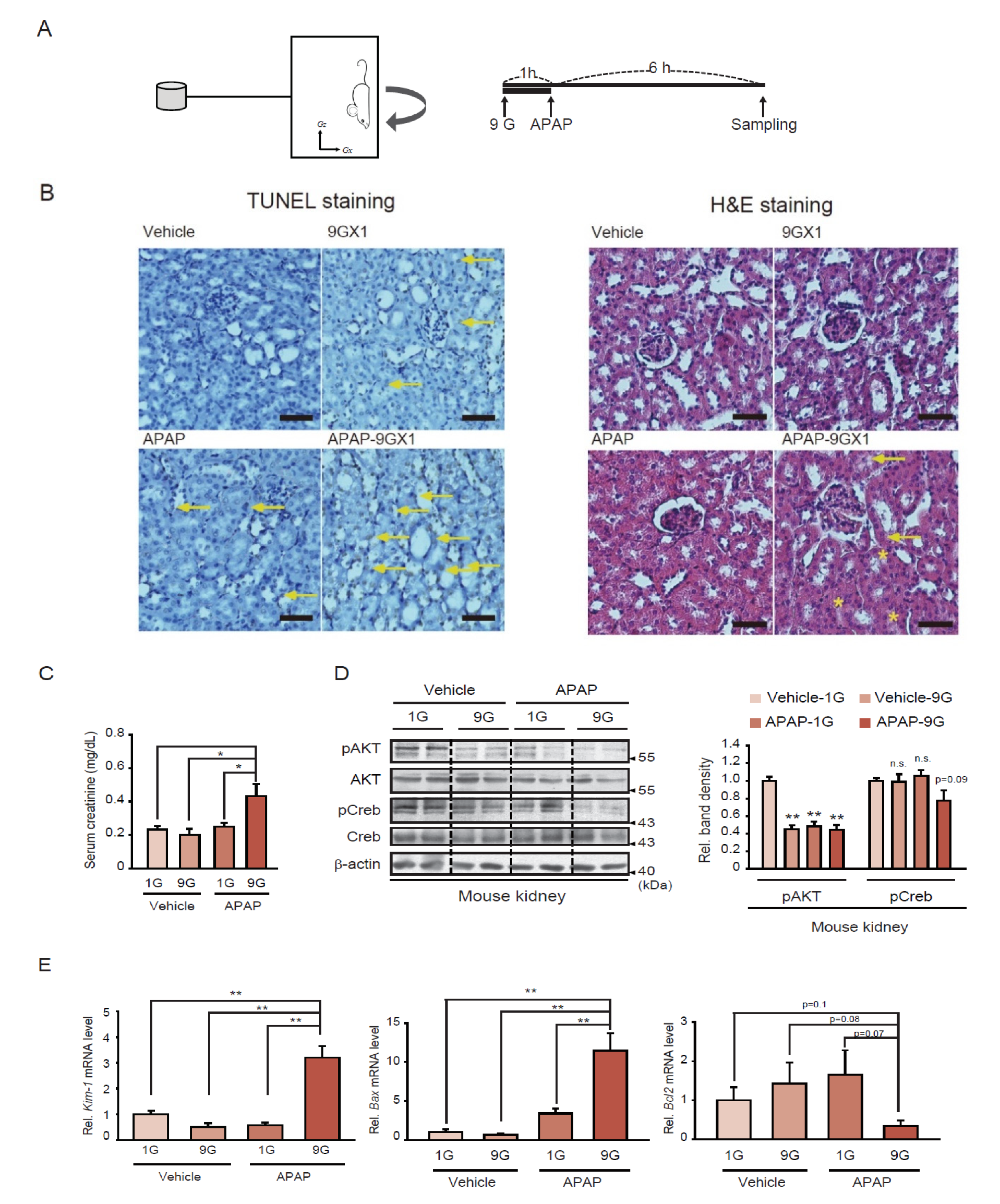
2.1. Kidney Injury Associated with a Single Hypergravity Load Coupled with APAP Treatment
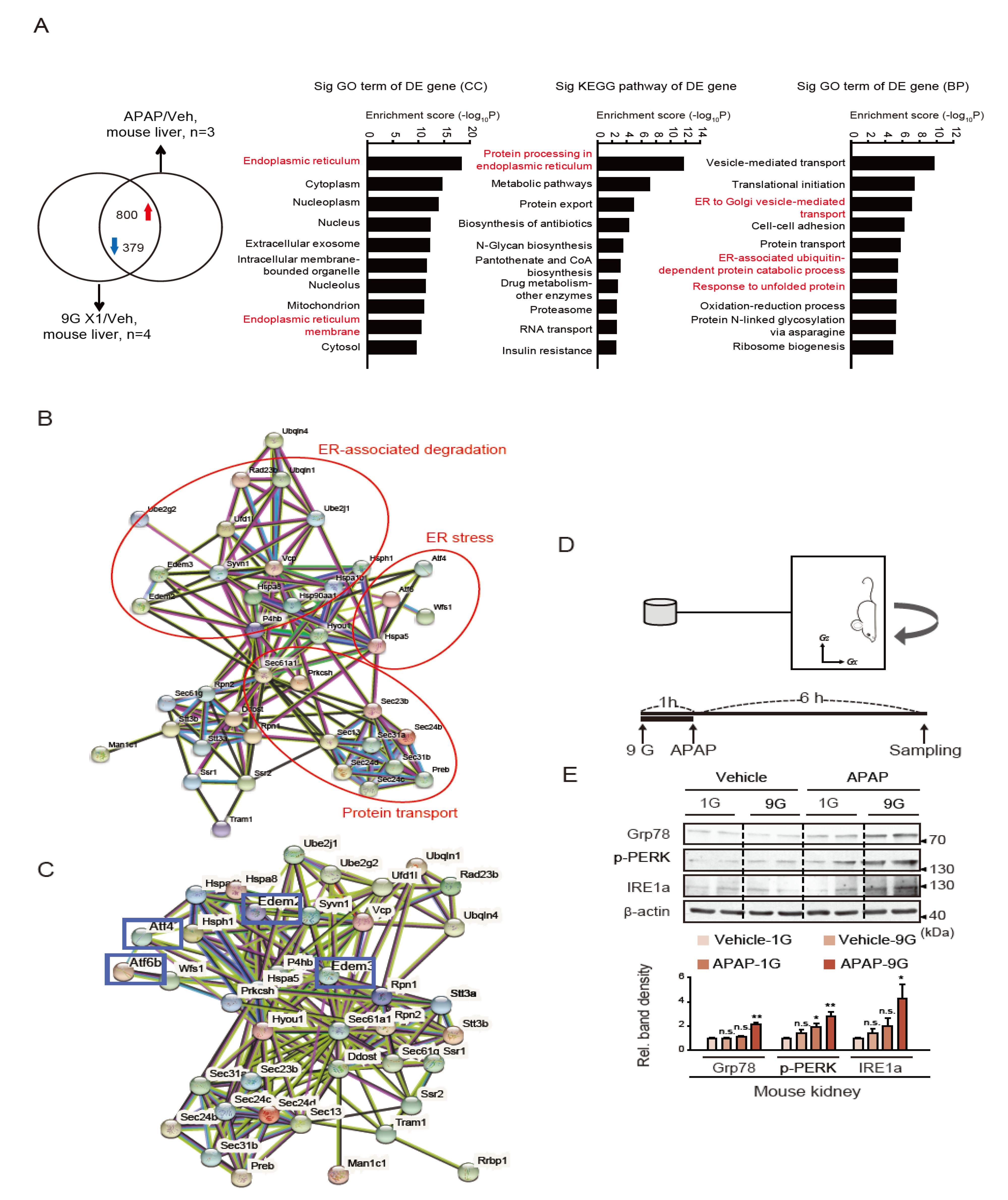
2.2. Induction of ER Stress by a Single Hypergravity Load Coupled with APAP Treatment
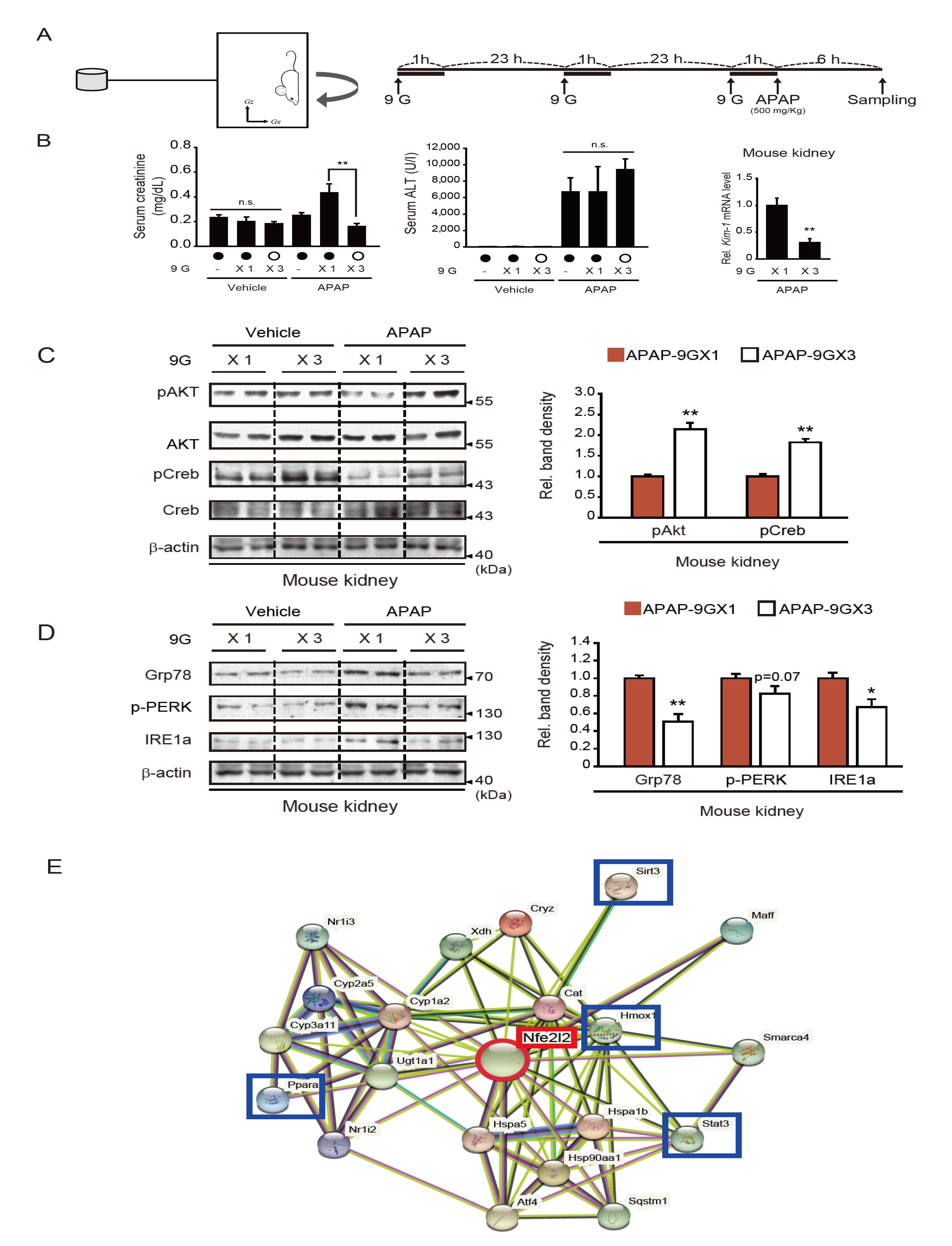
2.3. Repeated +9 Gx Hypergravity Exposure Attenuates APAP-Induced Renal Injury via Preconditioning
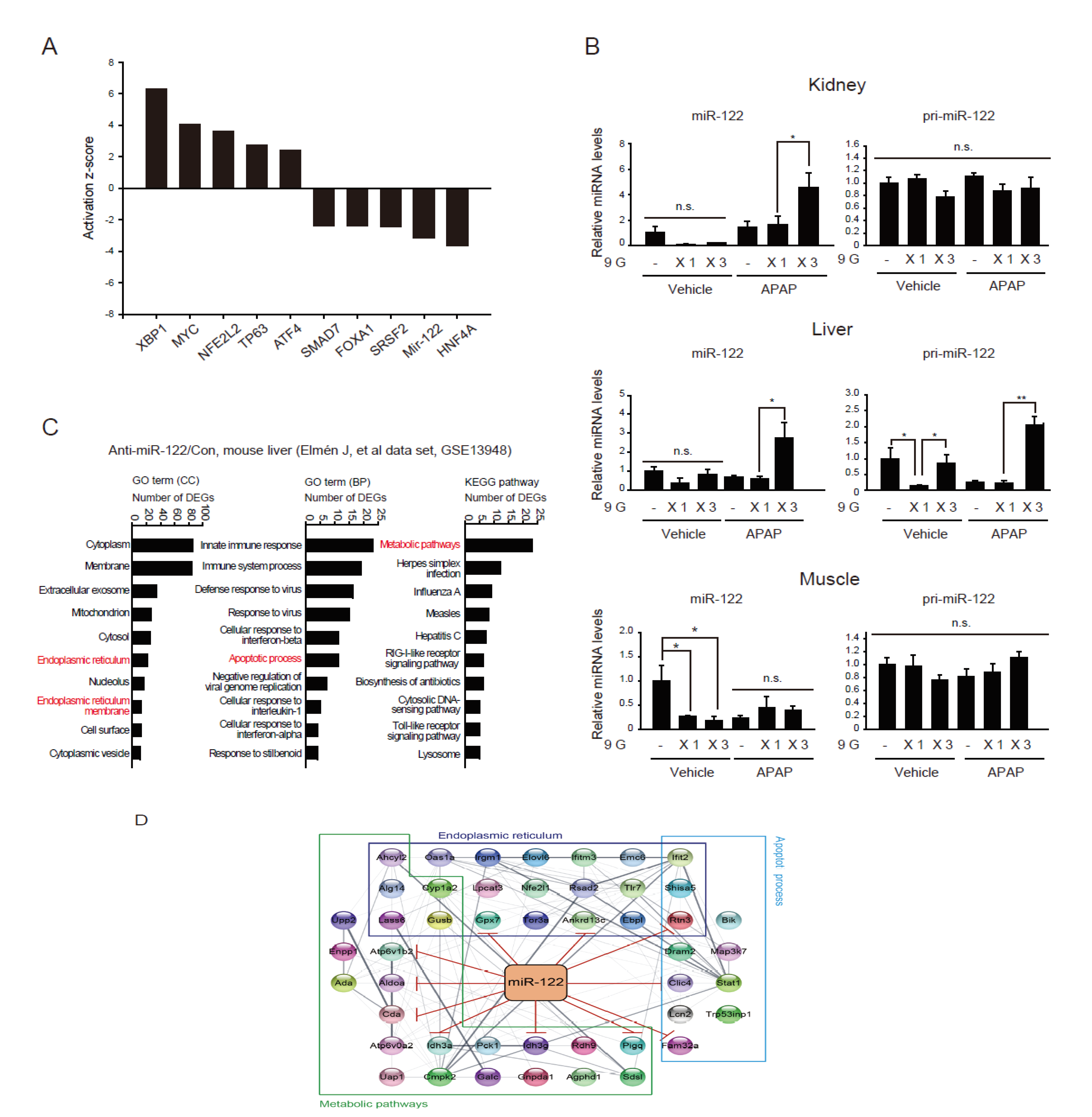
2.4. Adaptive Increases in miR-122 Levels in the Kidney after Preconditioning with Multiple Hypergravity Loads



3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Centrifugation Experiment
4.3. Blood Chemistry
4.4. Hematoxylin and Eosin Staining
4.5. TUNEL Assays
4.6. Immunoblot Analysis
4.7. Real-Time Polymerase Chain Reaction Assays
4.8. RNA Quality Check
4.9. Integrative Network Analysis
4.10. Data Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| ALT | Alanine aminotransferase |
| APAP | Acetaminophen |
| Bax | Bcl-2-associated X protein |
| DEG | Differentially expressed gene |
| ER | Endoplasmic reticulum |
| GEO | Gene expression Omnibus |
| miR | MicroRNA |
| mRNA | Messenger RNA |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| TUNEL | Terminal deoxynucleotidyl transferase dUTP nick end labeling |
References
- De Cesari, C.; Barravecchia, I.; Pyankova, O.V.; Vezza, M.; Germani, M.M.; Scebba, F.; Van Loon, J.; Angeloni, D. Hypergravity Activates a Pro-Angiogenic Homeostatic Response by Human Capillary Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 2354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tominari, T.; Ichimaru, R.; Taniguchi, K.; Yumoto, A.; Shirakawa, M.; Matsumoto, C.; Watanabe, K.; Hirata, M.; Itoh, Y.; Shiba, D.; et al. Hypergravity and microgravity exhibited reversal effects on the bone and muscle mass in mice. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Mao, X.W.; Nishiyama, N.C.; Byrum, S.D.; Stanbouly, S.; Jones, T.; Holley, J.; Sridharan, V.; Boerma, M.; Tackett, A.J.; Willey, J.S.; et al. Spaceflight induces oxidative damage to blood-brain barrier integrity in a mouse model. FASEB J. 2020, 34, 15516–15530. [Google Scholar] [CrossRef]
- Del Signore, A.; Mandillo, S.; Rizzo, A.; Di Mauro, E.; Mele, A.; Negri, R.; Oliverio, A.; Paggi, P. Hippocampal gene expression is modulated by hypergravity. Eur. J. Neurosci. 2004, 19, 667–677. [Google Scholar] [CrossRef]
- Gil Lee, S.; Wu, H.M.; Lee, C.G.; Oh, C.S.; Chung, S.W.; Kim, S.G. Binge Alcohol Intake After Hypergravity Stress Sustainably Decreases AMPK and Transcription Factors Necessary for Hepatocyte Survival. Alcohol. Clin. Exp. Res. 2017, 41, 76–86. [Google Scholar] [CrossRef]
- Gil Lee, S.; Lee, C.G.; Wu, H.M.; Oh, C.S.; Chung, S.W.; Kim, S.G. A load of mice to hypergravity causes AMPKα repression with liver injury, which is overcome by preconditioning loads via Nrf2. Sci. Rep. 2015, 5, 15643. [Google Scholar] [CrossRef] [Green Version]
- Genchi, G.G.; Cialdai, F.; Monici, M.; Mazzolai, B.; Mattoli, V.; Ciofani, G. Hypergravity Stimulation Enhances PC12 Neuron-Like Cell Differentiation. BioMed Res. Int. 2015, 2015, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dechaumet, B.; Cleret, D.; Linossier, M.-T.; Vanden-Bossche, A.; Chanon, S.; Lefai, E.; Laroche, N.; Lafage-Proust, M.-H.; Vico, L. Hypergravity as a gravitational therapy mitigates the effects of knee osteoarthritis on the musculoskeletal system in a murine model. PLoS ONE 2020, 15, e0243098. [Google Scholar] [CrossRef]
- Ciofani, G.; Ricotti, L.; Rigosa, J.; Menciassi, A.; Mattoli, V.; Monici, M. Hypergravity effects on myoblast proliferation and differentiation. J. Biosci. Bioeng. 2012, 113, 258–261. [Google Scholar] [CrossRef]
- Ogneva, I.V.; Usik, M.A.; Biryukov, N.S.; Zhdankina, Y.S. Sperm Motility of Mice under Simulated Microgravity and Hypergravity. Int. J. Mol. Sci. 2020, 21, 5054. [Google Scholar] [CrossRef]
- Dara, L.; Ji, C.; Kaplowitz, N. The contribution of endoplasmic reticulum stress to liver diseases. Hepatology 2011, 53, 1752–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sieber, J.; Lindenmeyer, M.T.; Kampe, K.; Campbell, K.N.; Cohen, C.D.; Hopfer, H.; Mundel, P.; Jehle, A.W. Regulation of podocyte survival and endoplasmic reticulum stress by fatty acids. Am. J. Physiol. Physiol. 2010, 299, F821–F829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iurlaro, R.; Muñoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I.; Ron, D. Integrating the mechanisms of apoptosis induced by endoplasmic reticulum stress. Nat. Cell Biol. 2011, 13, 184–190. [Google Scholar] [CrossRef]
- Takeda, K.; Nagashima, S.; Shiiba, I.; Uda, A.; Tokuyama, T.; Ito, N.; Fukuda, T.; Matsushita, N.; Ishido, S.; Iwawaki, T.; et al. MITOL prevents ER stress-induced apoptosis by IRE 1α ubiquitylation at ER -mitochondria contact sites. EMBO J. 2019, 38, e100999. [Google Scholar] [CrossRef]
- Prola, A.; Da Silva, J.P.; Guilbert, A.; Lecru, L.; Piquereau, J.; Ribeiro, M.; Mateo, P.; Gressette, M.; Fortin, D.; Boursier, C.; et al. SIRT1 protects the heart from ER stress-induced cell death through eIF2α deacetylation. Cell Death Differ. 2017, 24, 343–356. [Google Scholar] [CrossRef] [Green Version]
- Mukherjee, S.; Singh, N.; Sengupta, N.; Fatima, M.; Seth, P.; Mahadevan, A.; Shankar, S.K.; Bhattacharyya, A.; Basu, A. Japanese encephalitis virus induces human neural stem/progenitor cell death by elevating GRP78, PHB and hnRNPC through ER stress. Cell Death Dis. 2018, 8, e2556. [Google Scholar] [CrossRef] [Green Version]
- Knupp, J.; Arvan, P.; Chang, A. Increased mitochondrial respiration promotes survival from endoplasmic reticulum stress. Cell Death Differ. 2018, 26, 487–501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Chen, Y.-J.; Dobbs, N.; Sakai, T.; Liou, J.; Miner, J.J.; Yan, N. STING-mediated disruption of calcium homeostasis chronically activates ER stress and primes T cell death. J. Exp. Med. 2019, 216, 867–883. [Google Scholar] [CrossRef]
- Lugea, A.; Gerloff, A.; Su, H.-Y.; Xu, Z.; Go, A.; Hu, C.; French, S.W.; Wilson, J.S.; Apte, M.V.; Waldron, R.T.; et al. The Combination of Alcohol and Cigarette Smoke Induces Endoplasmic Reticulum Stress and Cell Death in Pancreatic Acinar Cells. Gastroenterology 2017, 153, 1674–1686. [Google Scholar] [CrossRef] [Green Version]
- Koo, J.H.; Lee, H.J.; Kim, W.; Kim, S.G. Endoplasmic Reticulum Stress in Hepatic Stellate Cells Promotes Liver Fibrosis via PERK-Mediated Degradation of HNRNPA1 and Up-regulation of SMAD2. Gastroenterology 2016, 150, 181–193.e8. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y.; Lim, S.W.; Koo, J.H.; Kim, W.; Kim, S.G. PHLDA3 overexpression in hepatocytes by endoplasmic reticulum stress via IRE1-Xbp1s pathway expedites liver injury. Gut 2015, 65, 1377–1388. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.M. Acetaminophen and the U.S. acute liver failure study group: Lowering the risks of hepatic failure. Hepatology 2004, 40, 6–9. [Google Scholar] [CrossRef] [PubMed]
- James, L.; E Sullivan, J.; Roberts, D. The proper use of acetaminophen. Paediatr. Child Health 2011, 16, 544–547. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; McGill, M.R.; Cook, S.F.; Sharpe, M.R.; Winefield, R.D.; Wilkins, D.G.; Rollins, D.E.; Jaeschke, H. Time course of acetaminophen-protein adducts and acetaminophen metabolites in circulation of overdose patients and in HepaRG cells. Xenobiotica 2015, 45, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Reshi, M.S.; Shrivastava, S.; Jaswal, A.; Sinha, N.; Uthra, C.; Shukla, S. Gold nanoparticles ameliorate acetaminophen induced hepato-renal injury in rats. Exp. Toxicol. Pathol. 2017, 69, 231–240. [Google Scholar] [CrossRef]
- Mour, G.; Feinfeld, D.A.; Caraccio, T.; McGuigan, M. Acute renal dysfunction in acetaminophen poisoning. Ren. Fail. 2005, 27, 381–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.M.; Seo, S.Y.; Kim, T.H.; Kim, S.G. Decrease of microRNA-122 causes hepatic insulin resistance by inducing protein tyrosine phosphatase 1B, which is reversed by licorice flavonoid. Hepatology 2012, 56, 2209–2220. [Google Scholar] [CrossRef] [PubMed]
- Joo, M.S.; Shin, S.-B.; Kim, E.J.; Koo, H.J.; Yim, H.; Kim, S.G. Nrf2-lncRNA controls cell fate by modulating p53-dependent Nrf2 activation as an miRNA sponge for Plk2 and p21 cip1. FASEB J. 2019, 33, 7953–7969. [Google Scholar] [CrossRef] [Green Version]
- Han, D.; Lerner, A.G.; Walle, L.V.; Upton, J.-P.; Xu, W.; Hagen, A.; Backes, B.J.; Oakes, S.A.; Papa, F.R. IRE1α Kinase Activation Modes Control Alternate Endoribonuclease Outputs to Determine Divergent Cell Fates. Cell 2009, 138, 562–575. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.; Chevet, E.; Oakes, S.A. Proteostasis control by the unfolded protein response. Nat. Cell Biol. 2015, 17, 829–838. [Google Scholar] [CrossRef] [Green Version]
- Hetz, C.; Papa, F.R. The Unfolded Protein Response and Cell Fate Control. Mol. Cell 2018, 69, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allagnat, F.; Christulia, F.; Ortis, F.; Pirot, P.; Lortz, S.; Lenzen, S.; Eizirik, D.L.; Cardozo, A.K. Sustained production of spliced X-box binding protein 1 (XBP1) induces pancreatic beta cell dysfunction and apoptosis. Diabetology 2010, 53, 1120–1130. [Google Scholar] [CrossRef] [Green Version]
- Han, C.Y.; Rho, H.S.; Kim, A.; Kim, T.H.; Jang, K.; Jun, D.W.; Kim, J.W.; Kim, B.; Kim, S.G. FXR Inhibits Endoplasmic Reticulum Stress-Induced NLRP3 Inflammasome in Hepatocytes and Ameliorates Liver Injury. Cell Rep. 2018, 24, 2985–2999. [Google Scholar] [CrossRef] [Green Version]
- Krek, A.; Grün, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; Da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Trappe, T.A.; Carroll, C.C.; Dickinson, J.M.; Lemoine, J.K.; Haus, J.M.; Sullivan, B.E.; Lee, J.D.; Jemiolo, B.; Weinheimer, E.M.; Hollon, C.J. Influence of acetaminophen and ibuprofen on skeletal muscle adaptations to resistance exercise in older adults. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R655–R662. [Google Scholar] [CrossRef] [Green Version]
- Trappe, T.A.; White, F.; Lambert, C.P.; Cesar, D.; Hellerstein, M.; Evans, W.J. Effect of ibuprofen and acetaminophen on postexercise muscle protein synthesis. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E551–E556. [Google Scholar] [CrossRef] [Green Version]
- Hardie, D.G. AMPK: Positive and negative regulation, and its role in whole-body energy homeostasis. Curr. Opin. Cell Biol. 2015, 33, 1–7. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Dormoy-Raclet, V.; Cammas, A.; Celona, B.; Lian, X.J.; Van Der Giessen, K.; Zivojnovic, M.; Brunelli, S.; Riuzzi, F.; Sorci, G.; Wilhelm, B.T.; et al. HuR and miR-1192 regulate myogenesis by modulating the translation of HMGB1 mRNA. Nat. Commun. 2013, 4, 2388. [Google Scholar] [CrossRef] [Green Version]
- Tadie, J.-M.; Bae, H.-B.; Deshane, J.S.; Bell, C.P.; Lazarowski, E.R.; Chaplin, D.D.; Thannickal, V.J.; Abraham, E.; Zmijewski, J.W. Toll-Like Receptor 4 Engagement Inhibits Adenosine 5′-Monophosphate-Activated Protein Kinase Activation through a High Mobility Group Box 1 Protein-Dependent Mechanism. Mol. Med. 2012, 18, 659–668. [Google Scholar] [CrossRef]
- Mohar, I.; Stamper, B.D.; Rademacher, P.M.; White, C.C.; Nelson, S.D.; Kavanagh, T.J. Acetaminophen-induced liver damage in mice is associated with gender-specific adduction of peroxiredoxin-6. Redox Biol. 2014, 2, 377–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.M.; Kim, T.H.; Kim, A.; Koo, J.H.; Joo, M.S.; Kim, S.G. Liver X Receptor α-Induced Cannabinoid Receptor 2 Inhibits Ubiquitin-Specific Peptidase 4 Through miR-27b, Protecting Hepatocytes From TGF-β. Hepatol. Commun. 2019, 3, 1373–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.H.; Kim, Y.W.; Kim, S.G. AMPK-mediated GSK3β inhibition by isoliquiritigenin contributes to protecting mitochondria against iron-catalyzed oxidative stress. Biochem. Pharmacol. 2010, 79, 1352–1362. [Google Scholar] [CrossRef]
- Koo, J.H.; Kim, T.H.; Park, S.-Y.; Joo, M.S.; Han, C.Y.; Choi, C.S.; Kim, S.G. Gα13 ablation reprograms myofibers to oxidative phenotype and enhances whole-body metabolism. J. Clin. Investig. 2017, 127, 3845–3860. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, M.S.; Koo, J.H.; Kim, T.H.; Kim, Y.S.; Kim, S.G. LRH1-driven transcription factor circuitry for hepatocyte identity: Super-enhancer cistromic analysis. EBioMedicine 2019, 40, 488–503. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, H.-M.; Lee, S.-G.; Oh, C.-S.; Kim, S.-G. Hypergravity Load Modulates Acetaminophen Nephrotoxicity via Endoplasmic Reticulum Stress in Association with Hepatic microRNA-122 Expression. Int. J. Mol. Sci. 2021, 22, 4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094901
Wu H-M, Lee S-G, Oh C-S, Kim S-G. Hypergravity Load Modulates Acetaminophen Nephrotoxicity via Endoplasmic Reticulum Stress in Association with Hepatic microRNA-122 Expression. International Journal of Molecular Sciences. 2021; 22(9):4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094901
Chicago/Turabian StyleWu, Hong-Min, Sang-Gil Lee, Choong-Sik Oh, and Sang-Geon Kim. 2021. "Hypergravity Load Modulates Acetaminophen Nephrotoxicity via Endoplasmic Reticulum Stress in Association with Hepatic microRNA-122 Expression" International Journal of Molecular Sciences 22, no. 9: 4901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22094901