
A Novel Antimicrobial Peptide Sparanegtin Identified in Scylla paramamosain Showing Antimicrobial Activity and Immunoprotective Role In Vitro and Vivo

Abstract
:1. Introduction
2. Results
2.1. Cloning and Sequence Analysis of Sparanegtin
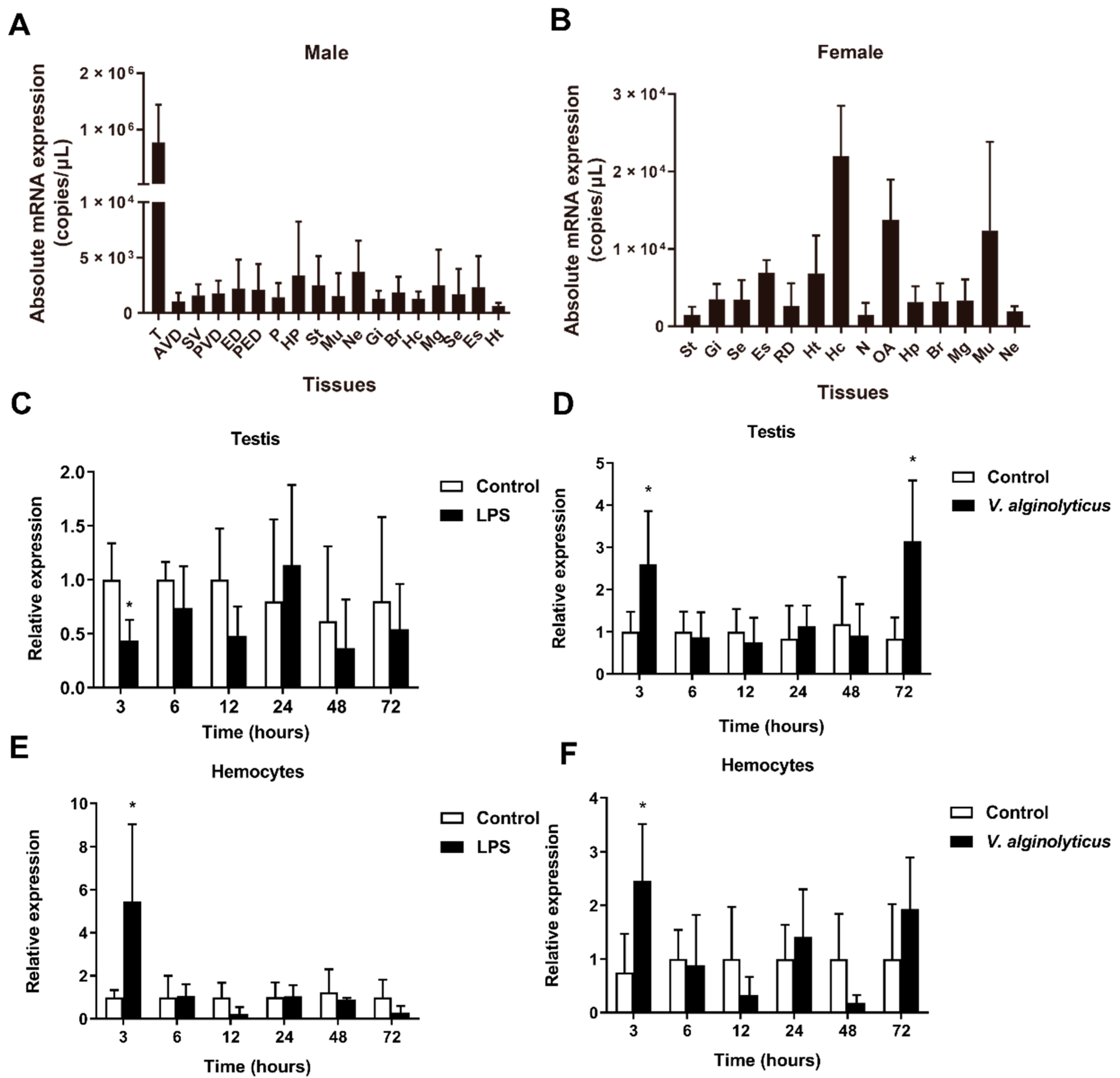
2.2. Gene Expression Profiles of Sparanegtin
2.3. rSparanegtin Shows Antimicrobial Activity
2.4. Preliminary Study on the Antibacterial Mechanism of rSparanegtin
2.4.1. Binding Properties
2.4.2. Killing Kinetic
2.4.3. rSparanegtin Induces Morphological Changes in Microorganisms
2.5. rSparanegtin Shows No Cytotoxicity and Could Reduce the V. alginolyticus Endotoxin Level In Vitro
2.6. The Immunoprotective Effect of rSparanegtin on S. paramamosain
2.6.1. Survival Rate Comparison
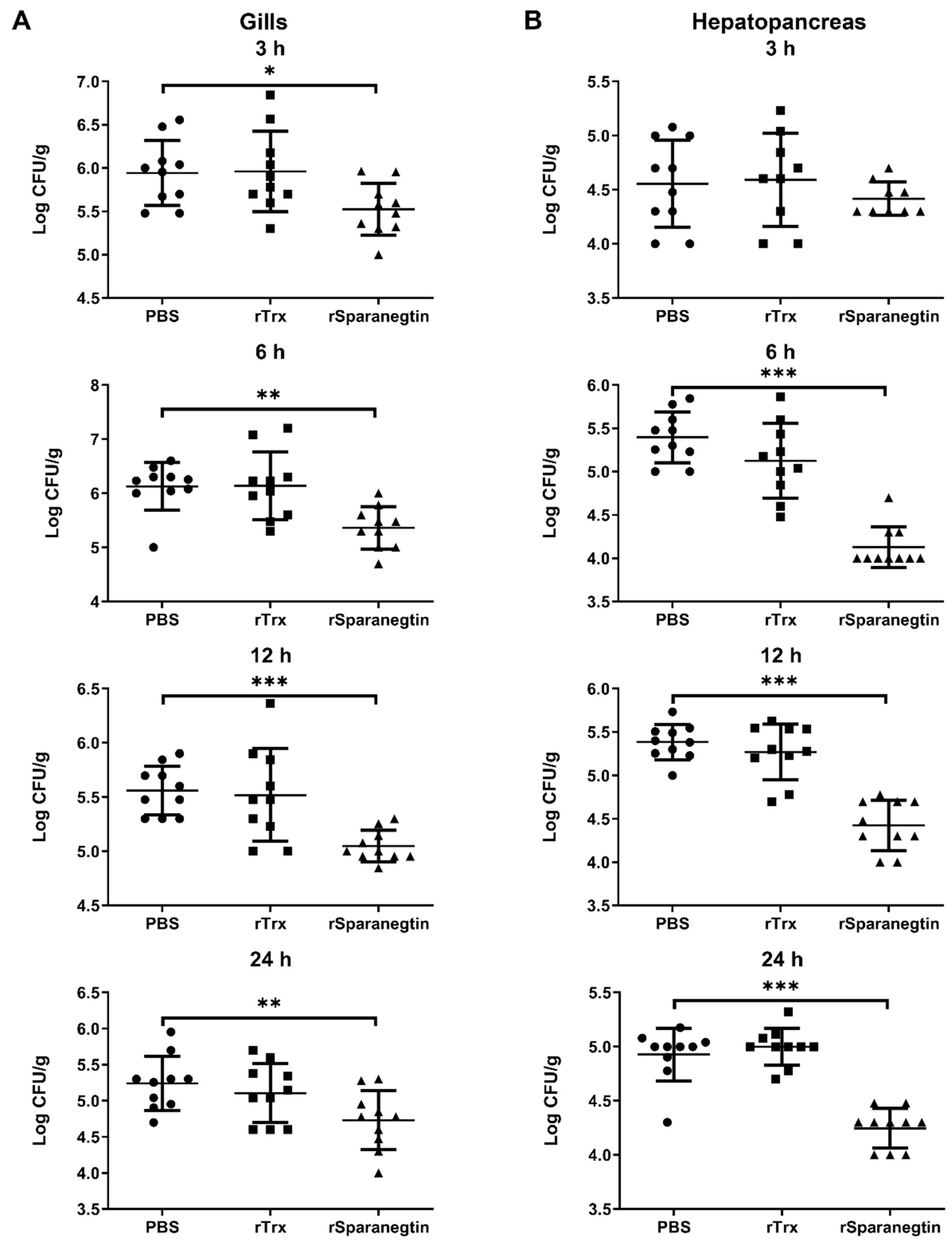
2.6.2. Pre-Incubation of rSparanegtin and V. alginolyticus Reduces Bacterial Load in the Tissues
2.6.3. Pre-Incubation of rSparanegtin and V. alginolyticus Modulate Immune-Related Gene Expression Profiles
3. Discussion
4. Materials and Methods
4.1. Microorganism Strains
4.2. Animals, Challenge and Tissue Collection
4.3. Cloning, Expression, Purification, and Analysis of Recombinant Proteins
4.4. Quantitative Real-Time PCR
4.5. Antimicrobial Assay
4.6. Binding Assays
4.7. Time-Killing Kinetic Assay
4.8. SEM Observation
4.9. Cytotoxicity Assay
4.10. Endotoxin Assay
4.11. Evaluation of the In Vivo Activity of rSparanegtin on S. paramamosain Infected with V. alginolyticus
4.12. Bacterial Load Assay and Quantification of Immune-Related Gene Expression after Different Treatment
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wenning, R. The State of World Fisheries and Aquaculture (Sofia) 2020 Report. Integr. Environ. Asses. 2020, 16, 800–801. [Google Scholar]
- Liu, X.; Steele, J.C.; Meng, X.Z. Usage, residue, and human health risk of antibiotics in Chinese aquaculture: A review. Environ. Pollut. 2017, 223, 161–169. [Google Scholar] [CrossRef]
- Shah, S.Q.; Cabello, F.C.; L’Abee-Lund, T.M.; Tomova, A.; Godfrey, H.P.; Buschmann, A.H.; Sorum, H. Antimicrobial resistance and antimicrobial resistance genes in marine bacteria from salmon aquaculture and non-aquaculture sites. Environ. Microbiol. 2014, 16, 1310–1320. [Google Scholar] [CrossRef]
- Verraes, C.; Van Boxstael, S.; Van Meervenne, E.; Van Coillie, E.; Butaye, P.; Catry, B.; de Schaetzen, M.A.; Van Huffel, X.; Imberechts, H.; Dierick, K.; et al. Antimicrobial Resistance in the Food Chain: A Review. Int. J. Environ. Res. Public Health 2013, 10, 2643–2669. [Google Scholar] [CrossRef] [Green Version]
- Brown, K.; Uwiera, R.R.E.; Kalmokoff, M.L.; Brooks, S.P.J.; Inglis, G.D. Antimicrobial growth promoter use in livestock: A requirement to understand their modes of action to develop effective alternatives. Int. J. Antimicrob. Agents 2017, 49, 12–24. [Google Scholar] [CrossRef]
- Tian, M.; He, X.M.; Feng, Y.Z.; Wang, W.T.; Chen, H.S.; Gong, M.; Liu, D.; Clarke, J.L.; van Eerde, A. Pollution by Antibiotics and Antimicrobial Resistance in LiveStock and Poultry Manure in China, and Countermeasures. Antibiotics 2021, 10, 539. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, A.; Onishi, H.; Eguchi, M. Quantitative PCR assay for the detection of the parasitic ciliate Cryptocaryon irritans. Fish. Sci. 2011, 77, 607–613. [Google Scholar] [CrossRef]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Shabir, U.; Ali, S.; Magray, A.R.; Ganai, B.A.; Firdous, P.; Hassan, T.; Nazir, R. Fish antimicrobial peptides (AMP'S) as essential and promising molecular therapeutic agents: A review. Microb. Pathog. 2018, 114, 50–56. [Google Scholar] [CrossRef]
- Batoni, G.; Maisetta, G.; Esin, S. Antimicrobial peptides and their interaction with biofilms of medically relevant bacteria. Biochim. Biophys. Acta 2016, 1858, 1044–1060. [Google Scholar] [CrossRef]
- Lu, S.; Walters, G.; Parg, R.; Dutcher, J.R. Nanomechanical response of bacterial cells to cationic antimicrobial peptides. Soft Matter 2014, 10, 1806–1815. [Google Scholar] [CrossRef]
- Hanson, M.A.; Dostalova, A.; Ceroni, C.; Poidevin, M.; Kondo, S.; Lemaitre, B. Synergy and remarkable specificity of antimicrobial peptides in vivo using a systematic knockout approach. Elife 2019, 8, e44341. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Lu, X.J.; Li, C.H.; Chen, J. Molecular characterization of the liver-expressed antimicrobial peptide 2 (LEAP-2) in a teleost fish, Plecoglossus altivelis: Antimicrobial activity and molecular mechanism. Mol. Immunol. 2015, 65, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.Y.; Wu, J.L.; Hui, C.F.; Lin, C.H.; Chen, J.Y. Insights into the antibacterial and immunomodulatory functions of the antimicrobial peptide, epinecidin-1, against Vibrio vulnificus infection in zebrafish. Fish. Shellfish Immunol. 2011, 31, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Li, H.X.; Lu, X.J.; Li, C.H.; Chen, J. Molecular characterization and functional analysis of two distinct liver-expressed antimicrobial peptide 2 (LEAP-2) genes in large yellow croaker (Larimichthys crocea). Fish. Shellfish Immunol. 2014, 38, 330–339. [Google Scholar] [CrossRef]
- Lin, F.Y.; Gao, Y.; Wang, H.; Zhang, Q.X.; Zeng, C.L.; Liu, H.P. Identification of an anti-lipopolysacchride factor possessing both antiviral and antibacterial activity from the red claw crayfish Cherax quadricarinatus. Fish. Shellfish Immunol. 2016, 57, 213–221. [Google Scholar] [CrossRef]
- Shan, Z.; Zhu, K.; Peng, H.; Chen, B.; Liu, J.; Chen, F.; Ma, X.; Wang, S.; Qiao, K.; Wang, K. The New Antimicrobial Peptide SpHyastatin from the Mud Crab Scylla paramamosain with Multiple Antimicrobial Mechanisms and High Effect on Bacterial Infection. Front. Microbiol. 2016, 7, 1140. [Google Scholar] [CrossRef] [PubMed]
- Long, S.; Chen, F.Y.; Wang, K.J. Characterization of a new homologous anti-lipopolysaccharide factor SpALF7 in mud crab Scylla paramamosain. Aquaculture 2021, 534, 736333. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, F.; Chen, H.Y.; Peng, H.; Hao, H.; Wang, K.J. A Novel Antimicrobial Peptide Scyreprocin From Mud Crab Scylla paramamosain Showing Potent Antifungal and Anti-biofilm Activity. Front. Microbiol. 2020, 11, 1589. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.Y.; Tsai, T.Y.; Su, B.C.; Hui, C.F.; Chen, J.Y. Study of the Antimicrobial Activity of Tilapia Piscidin 3 (TP3) and TP4 and Their Effects on Immune Functions in Hybrid Tilapia (Oreochromis spp.). PLoS ONE 2017, 12, e0169678. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.W.; Liu, C.H.; Hu, S.Y. Dietary administration of probiotic Paenibacillus ehimensis NPUST1 with bacteriocin-like activity improves growth performance and immunity against Aeromonas hydrophila and Streptococcus iniae in Nile tilapia (Oreochromis niloticus). Fish. Shellfish Immunol. 2019, 84, 695–703. [Google Scholar] [CrossRef]
- Schittek, B.; Hipfel, R.; Sauer, B.; Bauer, J.; Kalbacher, H.; Stevanovic, S.; Schirle, M.; Schroeder, K.; Blin, N.; Meier, F.; et al. Dermcidin: A novel human antibiotic peptide secreted by sweat glands. Nat. Immunol. 2001, 2, 1133–1137. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzon, D.; Saulnier, D.; Garnier, J.; Jouffrey, C.; Bulet, P.; Bachere, E. Crustacean immunity—Antifungal peptides are generated from the C terminus of shrimp hemocyanin in response to microbial challenge. J. Biol. Chem. 2001, 276, 47070–47077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.J.; Huang, W.S.; Yang, M.; Chen, H.Y.; Bo, J.; Li, S.J.; Wang, G.Z. A male-specific expression gene, encodes a novel anionic antimicrobial peptide, scygonadin, in Scylla serrata. Mol. Immunol. 2007, 44, 1961–1968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, K.; Xu, W.F.; Chen, H.Y.; Peng, H.; Zhang, Y.Q.; Huang, W.S.; Wang, S.P.; An, Z.; Shan, Z.G.; Chen, F.Y.; et al. A new antimicrobial peptide SCY2 identified in Scylla paramamosain exerting a potential role of reproductive immunity. Fish Shellfish Immunol. 2016, 51, 251–262. [Google Scholar] [CrossRef]
- Samakovlis, C.; Kylsten, P.; Kimbrell, D.A.; Engstrom, A.; Hultmark, A. The andropin gene and its product, a male-specific antibacterial peptide in Drosophila melanogaster. EMBO J. 1991, 10, 163–169. [Google Scholar] [CrossRef]
- Felgenhauer, B.E. Internal anatomy of the Decapoda: An overview. Microsc. Anat. Invertebr. 1992, 10, 45–75. [Google Scholar]
- Braga, A.; Nakayama, C.L.; Poersch, L.; Wasielesky, W. Unistellate spermatozoa of decapods: Comparative evaluation and evolution of the morphology. Zoomorphology 2013, 132, 261–284. [Google Scholar] [CrossRef]
- Sun, C.; Xu, W.T.; Zhang, H.W.; Dong, L.P.; Zhang, T.; Zhao, X.F.; Wang, J.X. An anti-lipopolysaccharide factor from red swamp crayfish, Procambarus clarkii, exhibited antimicrobial activities in vitro and in vivo. Fish Shellfish Immunol. 2011, 30, 295–303. [Google Scholar] [CrossRef]
- Liu, N.; Lan, J.F.; Sun, J.J.; Jia, W.M.; Zhao, X.F.; Wang, J.X. A novel crustin from Marsupenaeus japonicus promotes hemocyte phagocytosis. Dev. Comp. Immunol. 2015, 49, 313–322. [Google Scholar] [CrossRef]
- Li, M.; Ma, C.X.; Zhu, P.; Yang, Y.H.; Lei, A.Y.G.; Chen, X.H.; Liang, W.W.; Chen, M.; Xiong, J.H.; Li, C.Z. A new crustin is involved in the innate immune response of shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2019, 94, 398–406. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Koh, J.J.; Liu, S.; Lakshminarayanan, R.; Verma, C.S.; Beuerman, R.W. Membrane Active Antimicrobial Peptides: Translating Mechanistic Insights to Design. Front. Neurosci. 2017, 11, 73. [Google Scholar] [CrossRef] [Green Version]
- Shan, Z.G.; Zhu, K.X.; Chen, F.Y.; Liu, J.; Chen, B.; Qiao, K.; Peng, H.; Wang, K.J. In vivo activity and the transcriptional regulatory mechanism of the antimicrobial peptide SpHyastatin in Scylla paramamosain. Fish Shellfish Immunol. 2016, 59, 155–165. [Google Scholar] [CrossRef]
- Pushpanathan, M.; Gunasekaran, P.; Rajendhran, J. Antimicrobial peptides: Versatile biological properties. Int. J. Pept. 2013, 2013, 675391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, M.C.; Lin, S.B.; Lee, S.C.; Lin, C.C.; Hui, C.F.; Chen, J.Y. Antimicrobial peptide of an anti-lipopolysaccharide factor modulates of the inflammatory response in RAW264.7 cells. Peptides 2010, 31, 1262–1272. [Google Scholar] [CrossRef] [PubMed]
- Pan, C.Y.; Chao, T.T.; Chen, J.C.; Chen, J.Y.; Liu, W.C.; Lin, C.H.; Kuo, C.M. Shrimp (Penaeus monodon) anti-lipopolysaccharide factor reduces the lethality of Pseudomonas aeruginosa sepsis in mice. Int. Immunopharmacol. 2007, 7, 687–700. [Google Scholar] [CrossRef] [PubMed]
- Vallespi, M.G.; Alvarez-Obregon, J.C.; Rodriguez-Alonso, I.; Montero, T.; Garay, H.; Reyes, O.; Arana, M.J. A Limulus anti-LPS factor-derived peptide modulates cytokine gene expression and promotes resolution of bacterial acute infection in mice. Int. Immunopharmacol. 2003, 3, 247–256. [Google Scholar] [CrossRef]
- Li, W.X. Canonical and non-canonical JAK-STAT signaling. Trends Cell Biol. 2008, 18, 545–551. [Google Scholar] [CrossRef] [Green Version]
- Chu, S.H.; Liu, L.; Abbas, M.N.; Li, Y.Y.; Kausar, S.; Qian, X.Y.; Ye, Z.Z.; Yu, X.M.; Li, X.K.; Liu, M.; et al. Peroxiredoxin 6 modulates Toll signaling pathway and protects DNA damage against oxidative stress in red swamp crayfish (Procambarus clarkii). Fish Shellfish Immunol. 2019, 89, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Z.C.; Wan, Z.C.; Yang, L.; Li, W.W.; Wang, Q. JAK/STAT signalling regulates antimicrobial activities in Eriocheir sinensis. Fish Shellfish Immunol. 2019, 84, 491–501. [Google Scholar] [CrossRef]
- Dharmaraja, A.T. Role of Reactive Oxygen Species (ROS) in Therapeutics and Drug Resistance in Cancer and Bacteria. J. Med. Chem. 2017, 60, 3221–3240. [Google Scholar] [CrossRef] [PubMed]
- Kanzaki, H.; Wada, S.; Narimiya, T.; Yamaguchi, Y.; Katsumata, Y.; Itohiya, K.; Fukaya, S.; Miyamoto, Y.; Nakamura, Y. Pathways that Regulate ROS Scavenging Enzymes, and Their Role in Defense Against Tissue Destruction in Periodontitis. Front. Physiol. 2017, 8, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.S.; Zhang, Q.; Zhao, Y.R.; Jia, W.M.; Zhao, X.F.; Wang, J.X. A new group of anti-lipopolysaccharide factors from Marsupenaeus japonicus functions in antibacterial response. Dev. Comp. Immunol. 2015, 48, 33–42. [Google Scholar] [CrossRef]
- Liu, N.; Zhang, R.R.; Fan, Z.X.; Zhao, X.F.; Wang, X.W.; Wang, J.X. Characterization of a type-I crustin with broad-spectrum antimicrobial activity from red swamp crayfish Procambarus clarkii. Dev. Comp. Immunol. 2016, 61, 145–153. [Google Scholar] [CrossRef]
- Guo, Z.L.; Qiao, X.; Cheng, R.M.; Shi, N.N.; Wang, A.L.; Feng, T.T.; Chen, Y.; Zhang, F.; Yu, H.; Wang, Y.P. As-CATH4 and 5, two vertebrate-derived natural host defense peptides, enhance the immuno-resistance efficiency against bacterial infections in Chinese mitten crab, Eriocheir sinensis. Fish Shellfish Immunol. 2017, 71, 202–209. [Google Scholar] [CrossRef]
- Xu, W.F.; Qiao, K.; Huang, S.P.; Peng, H.; Huang, W.S.; Chen, F.Y.; Zhang, N.; Wang, G.Z.; Wang, K.J. The expression pattern of scygonadin during the ontogenesis of Scylla paramamosain predicting its potential role in reproductive immunity. Dev. Comp. Immunol. 2011, 35, 1076–1088. [Google Scholar] [CrossRef]
- Chen, F.Y.; Liu, H.P.; Bo, J.; Ren, H.L.; Wang, K.J. Identification of genes differentially expressed in hemocytes of Scylla paramamosain in response to lipopolysaccharide. Fish Shellfish Immunol. 2010, 28, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, S.S.; Chen, G.X.; Xu, H.D.; Zou, W.B.; Yan, W.R.; Wang, Q.Q.; Deng, H.W.; Zhang, H.Q.; Yu, G.J.; He, J.G.; et al. Transcriptome analysis of mud crab (Scylla paramamosain) gills in response to Mud crab reovirus (MCRV). Fish Shellfish Immunol. 2017, 60, 545–553. [Google Scholar] [CrossRef]
- Wei, L.; Yang, J.; He, X.; Mo, G.; Hong, J.; Yan, X.; Lin, D.; Lai, R. Structure and function of a potent lipopolysaccharide-binding antimicrobial and anti-inflammatory peptide. J. Med. Chem. 2013, 56, 3546–3556. [Google Scholar] [CrossRef] [PubMed]
- Deepika, A.; Makesh, M.; Rajendran, K.V. Development of primary cell cultures from mud crab, Scylla serrata, and their potential as an in vitro model for the replication of white spot syndrome virus. In Vitro Cell Dev. Biol.-Anim. 2014, 50, 406–416. [Google Scholar] [CrossRef] [PubMed]
- Qiu, W.; Chen, F.; Chen, R.; Li, S.; Zhu, X.; Xiong, M.; Wang, K.J. A New C-Type Lectin Homolog SpCTL6 Exerting Immunoprotective Effect and Regulatory Role in Mud Crab Scylla paramamosain. Front. Immunol. 2021, 12, 661823. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganisms. | CGMCC No. a | MIC (μM) b | MBC (μM) b |
|---|---|---|---|
| Gram-negative bacteria | |||
| Escherichia coli | 1.2389 | 24–48 | 24–48 |
| Pseudomonas aeruginosa | 1.2421 | 12–24 | 24–48 |
| Pseudomonas fluorescens | 1.1802 | 12–24 | >48 |
| Aeromonas hydrophila | 1.2017 | 12–24 | >48 |
| Shigella flexneri | 1.1868 | 12–24 | >48 |
| Gram-positive bacteria | |||
| Bacillus subtilis | 1.3358 | 24–48 | >48 |
| Staphylococcus epidermidis | 1.4260 | 24–48 | >48 |
| Staphylococcus aureus | 1.2465 | 12–24 | >48 |
| Fungi | |||
| Cryptococcus neoformans | 2.1563 | 24–48 | >48 |
| Pichia pastoris (GS115) | Invitrogen | 24–48 | >48 |
| Primers | Sequence (5′−3′) |
|---|---|
| Sparanegtin-ORF-F | ATGGCGGCGGCGGCTTCAGG |
| Sparanegtin-ORF-R | TCAAACCCCCGCCGAAGTGA |
| Sparanegtin-5′-R1 | CGGCTGCCGTAGGAAAGGAA |
| Sparanegtin-5′-R2 | AATATCCGCTGTGGCCGACG |
| Sparanegtin-3′-F1 | CGTCGGCCACAGCGGATATT |
| Sparanegtin-3′-F2 | ACCAACTTGGCAAGAGGAGCG |
| Long primer | CTAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT |
| Short primer | CTAATACGACTCACTATAGGGC |
| NUP | AAGCAGTGGTATCAACGCAGAGT |
| M13–47F | CGCCAGGGTTTTCCCAGTCACGAC |
| M13–48R | AGCGGATAACAATTTCACACAGGA |
| Sparanegtin-qPCR-F | TCCCCGGTTTCCCGACCCAG |
| Sparanegtin-qPCR-R | ACCAGGAGGCAGCACCGTCT |
| GAPDH-qPCR-F | CTCCACTGGTGCCGCTAAGGCTGTA |
| GAPDH-qPCR-R | CAAGTCAGGTCAACCACGGACACAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, X.; Chen, F.; Li, S.; Peng, H.; Wang, K.-J. A Novel Antimicrobial Peptide Sparanegtin Identified in Scylla paramamosain Showing Antimicrobial Activity and Immunoprotective Role In Vitro and Vivo. Int. J. Mol. Sci. 2022, 23, 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010015
Zhu X, Chen F, Li S, Peng H, Wang K-J. A Novel Antimicrobial Peptide Sparanegtin Identified in Scylla paramamosain Showing Antimicrobial Activity and Immunoprotective Role In Vitro and Vivo. International Journal of Molecular Sciences. 2022; 23(1):15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010015
Chicago/Turabian StyleZhu, Xuewu, Fangyi Chen, Shuang Li, Hui Peng, and Ke-Jian Wang. 2022. "A Novel Antimicrobial Peptide Sparanegtin Identified in Scylla paramamosain Showing Antimicrobial Activity and Immunoprotective Role In Vitro and Vivo" International Journal of Molecular Sciences 23, no. 1: 15. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010015