
Captopril Combined with Furosemide or Hydrochlorothiazide Affects Macrophage Functions in Mouse Contact Hypersensitivity Response


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
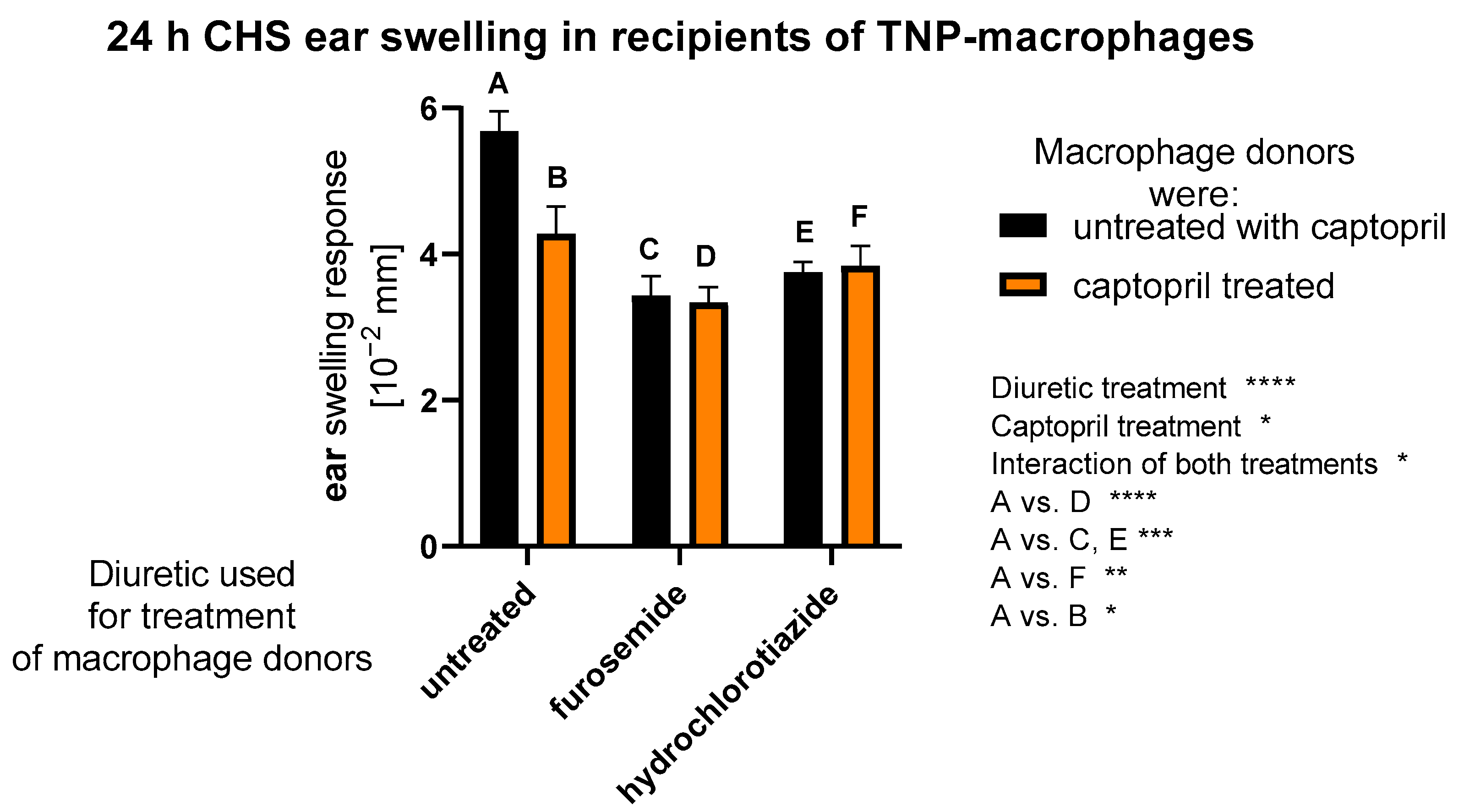
2.1. Treatment with Captopril with or without Diuretic Drug Affects CHS Response Induced by Macrophages
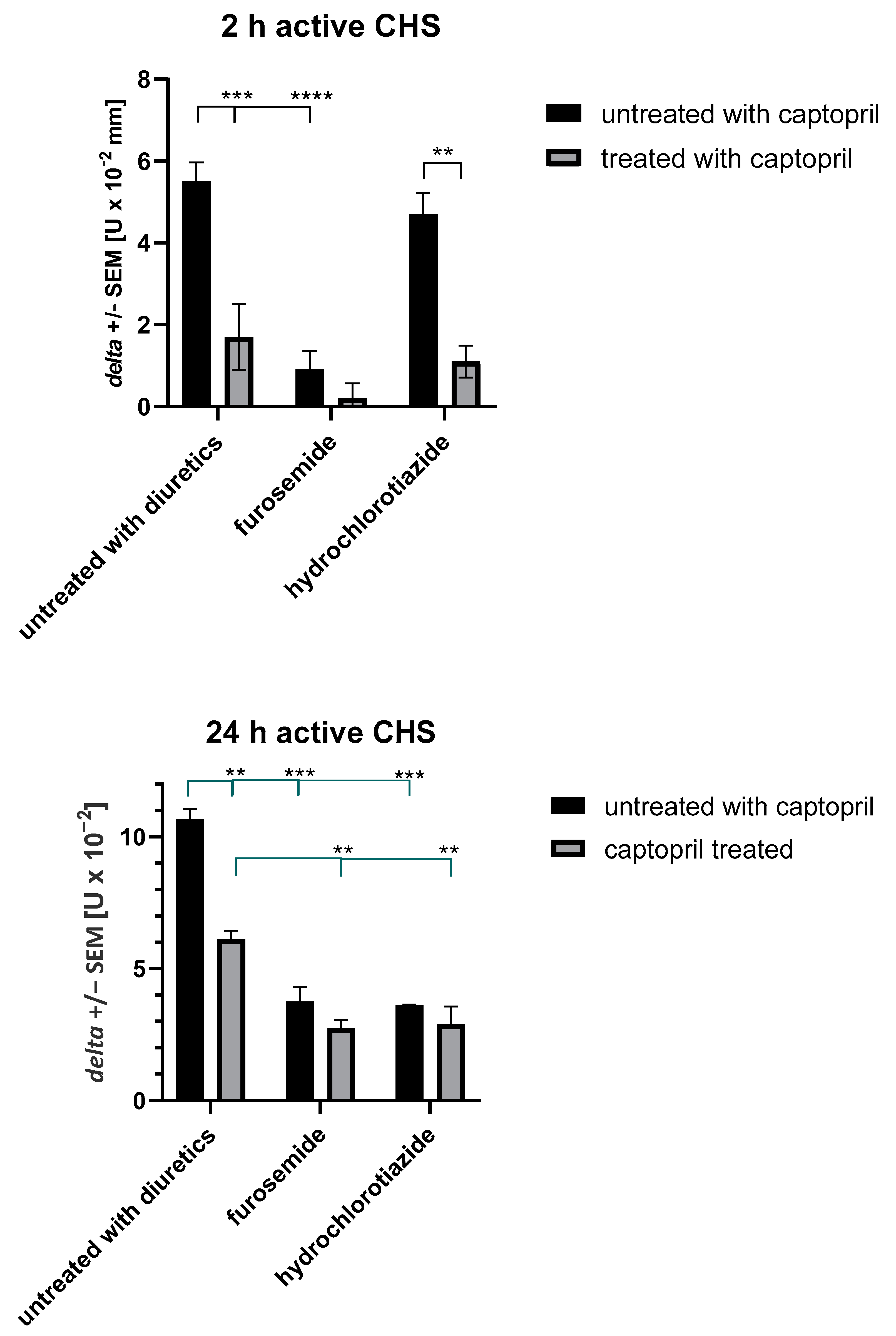
2.2. Treatment with Captopril with or without Diuretic Drug Affects Active CHS Response
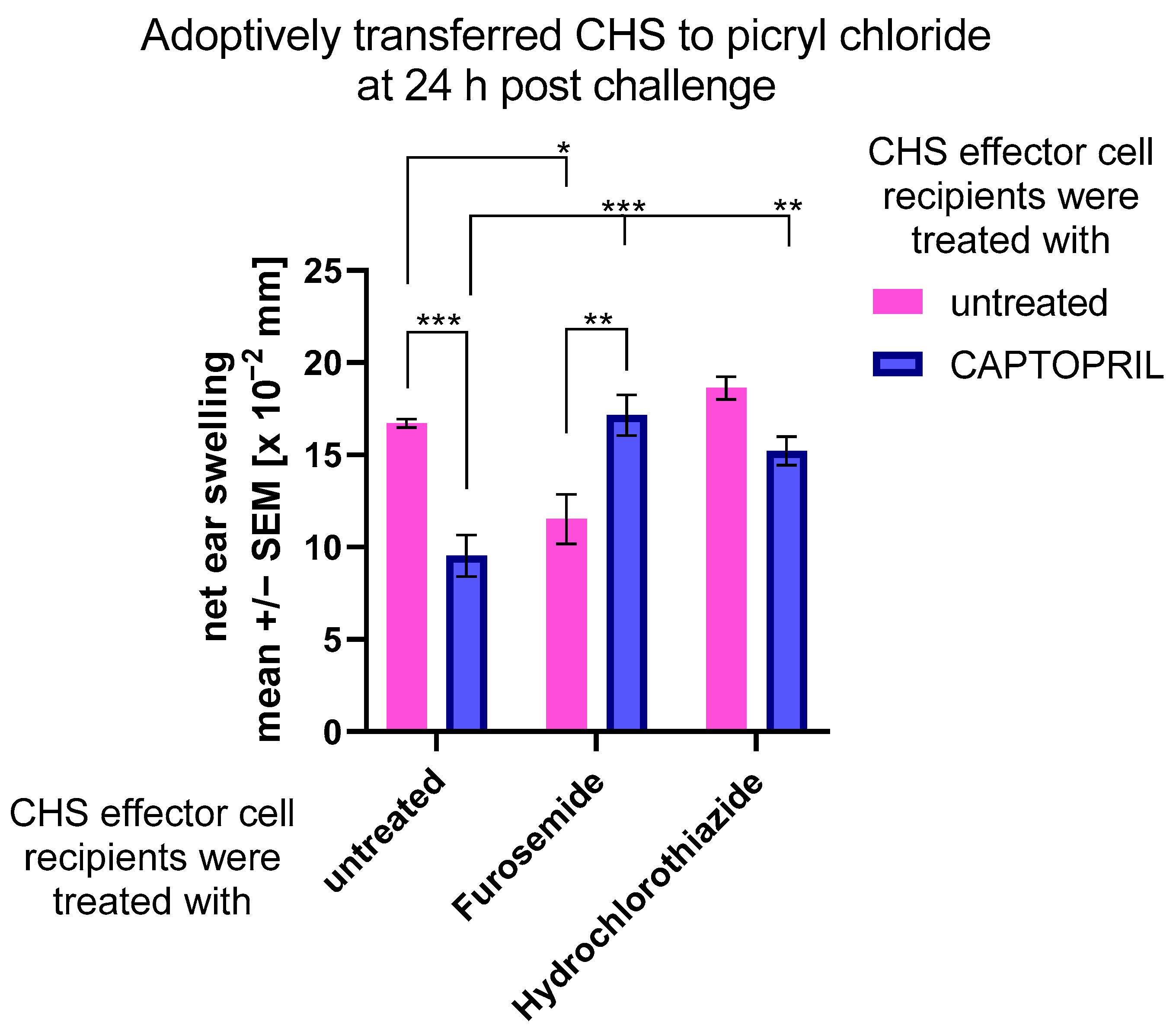
2.3. Treatment with Captopril with or without Diuretic Drug Differently Impacts the Adoptively Transferred CHS Response
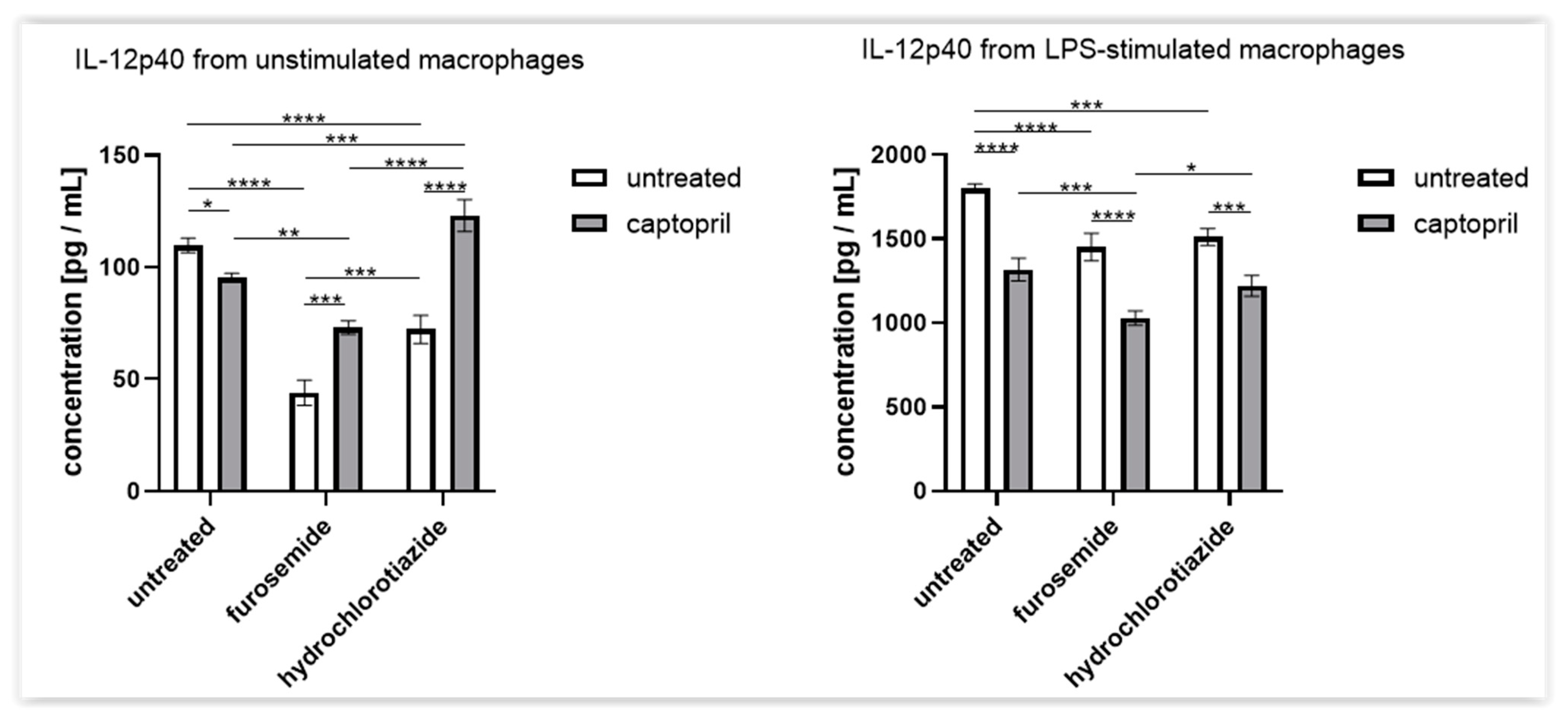
2.4. Treatment with Captopril with or without Diuretic Drug Differently Influences the Secretion of IL-12p40 by Macrophages
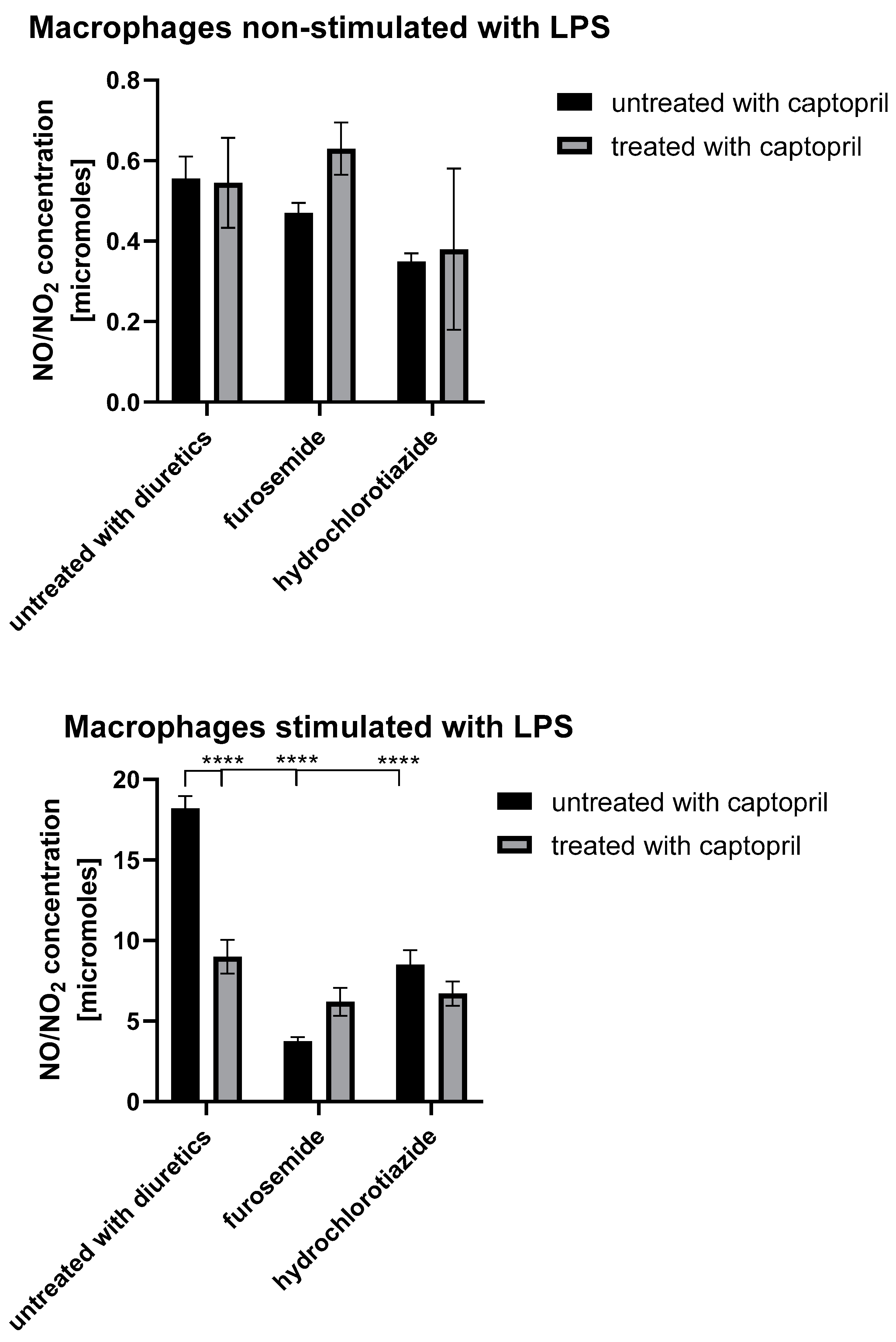
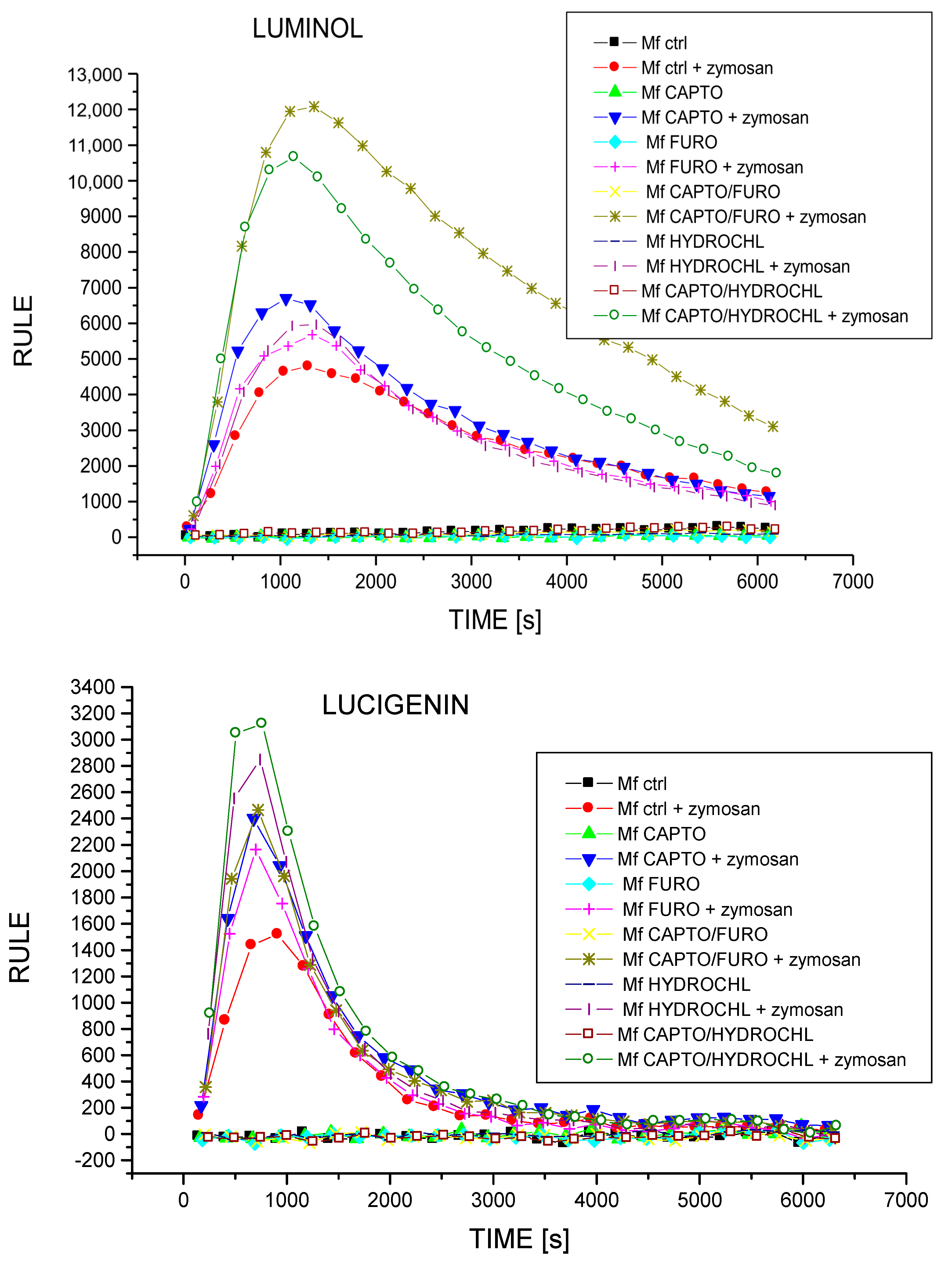
2.5. Treatment with Captopril with or without Diuretic Drug Differently Influences the Secretion of NO and ROIs by Macrophages
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Antihypertensive Drug Administration
4.3. Harvest of Oil-Induced Peritoneal Macrophages
4.4. Assessing ROI Generation by Macrophages in Chemiluminescence Assay
4.5. Measurement of IL-12p40 and Nitric Oxide Concentration in Macrophage Culture Supernatant
4.6. Induction of Active or Adoptively Transferred CHS Reaction in Mice
4.7. Transfer of Hapten-Conjugated Macrophages to Induce CHS Reaction
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Whelton, P.K.; Carey, R.M.; Aronow, W.S.; Casey, D.E., Jr.; Collins, K.J.; Dennison Himmelfarb, C.; DePalma, S.M.; Gidding, S.; Jamerson, K.A.; Jones, D.W.; et al. 2017 ACC/AHA/AAPA/ABC/ACPM/AGS/APhA/ASH/ASPC/NMA/PCNA Guideline for the Prevention, Detection, Evaluation, and Management of High Blood Pressure in Adults: A Report of the American College of Cardiology/American Heart Association Task Force on Clinical Practice Guidelines. J. Am. Coll. Cardiol. 2018, 71, e127–e248. [Google Scholar] [PubMed]
- Furberg, C.D.; Wright, J.T.; Davis, B.R.; Cutler, J.A.; Alderman, M.; Black, H.; Cushman, W.; Grimm, R.; Haywood, L.J.; Leenen, F.; et al. Major outcomes in high-risk hypertensive patients randomized to angiotensin-converting enzyme inhibitor or calcium channel blocker vs. diuretic: The Antihypertensive and Lipid-Lowering Treatment to Prevent Heart Attack Trial (ALLHAT). J. Am. Med. Assoc. 2002, 288, 2981–2997. [Google Scholar]
- Weber, M.A.; Julius, S.; Kjeldsen, S.E.; Brunner, H.R.; Ekman, S.; Hansson, L.; Hua, T.; Laragh, J.H.; McInnes, G.T.; Mitchell, L.; et al. Blood pressure dependent and independent effects of antihypertensive treatment on clinical events in the VALUE Trial. Lancet 2004, 363, 2049–2051. [Google Scholar] [CrossRef]
- JJulius, S.; Kjeldsen, S.E.; Weber, M.; Brunner, H.R.; Ekman, S.; Hansson, L.; Hua, T.; Laragh, J.; McInnes, G.T.; Mitchell, L.; et al. Outcomes in hypertensive patients at high cardiovascular risk treated with regimens based on valsartan or amlodipine: The VALUE randomised trial. Lancet 2004, 363, 2022–2031. [Google Scholar] [CrossRef]
- Nissen, S.E.; Tuzcu, E.M.; Libby, P.; Thompson, P.D.; Ghali, M.; Garza, D.; Berman, L.; Shi, H.; Buebendorf, E.; Topol, E.J.; et al. Effect of antihypertensive agents on cardiovascular events in patients with coronary disease and normal blood pressure: The CAMELOT study: A randomized controlled trial. JAMA 2004, 292, 2217–2225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, B.; Mancia, G.; Spiering, W.; Agabiti Rosei, E.; Azizi, M.; Burnier, M.; Clement, D.L.; Coca, A.; De Simone, G.; Dominiczak, A.; et al. 2018 ESC/ESH Guidelines for the management of arterial hypertension. Eur. Heart J. 2018, 39, 3021–3104. [Google Scholar] [CrossRef]
- Garjón, J.; Saiz, L.C.; Azparren, A.; Gaminde, I.; Ariz, M.J.; Erviti, J. First-line combination therapy versus first-line monotherapy for primary hypertension. Cochrane Database Syst. Rev. 2020, 2, CD010316. [Google Scholar] [CrossRef]
- Law, M.R.; Morris, J.K.; Wald, N.J. Use of blood pressure lowering drugs in the prevention of cardiovascular disease: Meta-analysis of 147 randomised trials in the context of expectations from prospective epidemiological studies. BMJ 2009, 338, b1665. [Google Scholar] [CrossRef] [Green Version]
- Epstein, M.; Bakris, G. Newer approaches to antihypertensive therapy. Use of fixed-dose combination therapy. Arch. Intern. Med. 1996, 156, 1969–1978. [Google Scholar] [CrossRef]
- Bangalore, S.; Kamalakkannan, G.; Parkar, S.; Messerli, F.H. Fixed-dose combinations improve medication compliance: A meta-analysis. Am. J. Med. 2007, 120, 713–719. [Google Scholar] [CrossRef]
- Brixner, D.I.; Jackson, K.C.; Sheng, X.; Nelson, R.E.; Keskinaslan, A. Assessment of adherence, persistence, and costs among valsartan and hydrochlorothiazide retrospective cohorts in free-and fixed-dose combinations. Curr. Med. Res. Opin. 2008, 24, 2597–2607. [Google Scholar] [CrossRef]
- Brown, M.J.; McInnes, G.T.; Papst, C.C.; Zhang, J.; MacDonald, T.M. Aliskiren and the calcium channel blocker amlodipine combination as an initial treatment strategy for hypertension control (ACCELERATE): A randomised, parallel-group trial. Lancet 2011, 377, 312–320. [Google Scholar] [CrossRef]
- Egan, B.M.; Bandyopadhyay, D.; Shaftman, S.R.; Wagner, C.S.; Zhao, Y.; Yu-Isenberg, K.S. Initial monotherapy and combination therapy and hypertension control the first year. Hypertension 2012, 59, 1124–1131. [Google Scholar] [CrossRef] [Green Version]
- Thom, S.; Poulter, N.; Field, J.; Patel, A.; Prabhakaran, D.; Stanton, A.; Grobbee, D.E.; Bots, M.L.; Reddy, K.S.; Cidambi, R.; et al. Effects of a fixed-dose combination strategy on adherence and risk factors in patients with or at high risk of CVD: The UMPIRE randomized clinical trial. JAMA 2013, 310, 918–929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurent, S.; Mancia, G.; Poulter, N. Perindopril 3.5 mg/amlodipine 2.5 mg versus renin-angiotensin system inhibitor monotherapy as first-line treatment in hypertension: A combined analysis. J. Hypertens. 2018, 36, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Guerrero-García, C.; Rubio-Guerra, A.F. Combination therapy in the treatment of hypertension. Drugs Context 2018, 7, 212531. [Google Scholar] [CrossRef] [PubMed]
- Mitini-Nkhoma, S.C.; Fernando, N.; Ishaka, G.K.D.; Handunnetti, S.M.; Pathirana, S.L. Ion Transport Modulators Differentially Modulate Inflammatory Responses in THP-1-Derived Macrophages. J. Immunol. Res. 2021, 2021, 1–9. [Google Scholar] [CrossRef]
- SSun, N.-N.; Zhang, Y.; Huang, W.-H.; Zheng, B.-J.; Jin, S.-Y.; Li, X.; Meng, Y. Macrophage exosomes transfer angiotensin II type 1 receptor to lung fibroblasts mediating bleomycin-induced pulmonary fibrosis. Chin. Med J. 2021, 134, 2175–2185. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, H.; Ge, Y.; Cao, L.; He, Y.; Sun, G.; Jia, S.; Ma, A.; Liu, J.; Rong, D.; et al. AT1R Regulates Macrophage Polarization Through YAP and Regulates Aortic Dissection Incidence. Front. Physiol. 2021, 12, 644903. [Google Scholar] [CrossRef]
- Nazimek, K.; Filipczak-Bryniarska, I.; Bryniarski, K. The role of medicaments, exosomes and miRNA molecules in modulation of macrophage immune activity. Postepy Hig. I Med. Dosw. 2015, 69, 1114–1129. [Google Scholar]
- Shimada, K.; Murayama, T.; Yokode, M.; Kita, T.; Fujita, M.; Kishimoto, C. Olmesartan, a novel angiotensin II type 1 receptor antagonist, reduces severity of atherosclerosis in apolipoprotein E deficient mice associated with reducing superoxide production. Nutr. Metab. Cardiovasc. Dis. NMCD 2011, 21, 672–678. [Google Scholar] [CrossRef] [PubMed]
- Nazimek, K.; Kozlowski, M.; Bryniarski, P.; Strobel, S.; Bryk, A.; Myszka, M.; Tyszka, A.; Kuszmiersz, P.; Nowakowski, J.; Filipczak-Bryniarska, I. Repeatedly administered antidepressant drugs modulate humoral and cellular immune response in mice through action on macrophages. Exp. Biol. Med. 2016, 241, 1540–1550. [Google Scholar] [CrossRef]
- Nazimek, K.; Strobel, S.; Bryniarski, P.; Kozlowski, M.; Filipczak-Bryniarska, I.; Bryniarski, K. The role of macrophages in anti-inflammatory activity of antidepressant drugs. Immunobiology 2017, 222, 823–830. [Google Scholar] [CrossRef] [PubMed]
- Bryniarski, P.; Nazimek, K.; Marcinkiewicz, J. Anti-Inflammatory Activities of Captopril and Diuretics on Macrophage Activity in Mouse Humoral Immune Response. Int. J. Mol. Sci. 2021, 22, 11374. [Google Scholar] [CrossRef]
- Siragy, H.M.; Xue, C.; Webb, R.L. Beneficial effects of combined benazepril-amlodipine on cardiac nitric oxide, cGMP, and TNF-alpha production after cardiac ischemia. J. Cardiovasc. Pharmacol. 2006, 47, 636–642. [Google Scholar] [CrossRef]
- Fukuzawa, M.; Satoh, J.; Ohta, S.; Takahashi, K.; Miyaguchi, S.; Qiang, X.; Sakata, Y.; Nakazawa, T.; Takizawa, Y.; Toyota, T. Modulation of tumor necrosis factor-alpha production with anti-hypertensive drugs. Immunopharmacology 2000, 48, 65–74. [Google Scholar] [CrossRef]
- Nemati, F.; Rahbar-Roshandel, N.; Hosseini, F.; Mahmoudian, M.; Shafiei, M. Anti-inflammatory effects of anti-hypertensive agents: Influence on interleukin-1β secretion by peripheral blood polymorphonuclear leukocytes from patients with essential hypertension. Clin. Exp. Hypertens. 2011, 33, 66–76. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.-S.; Schulman, I.H.; Jaimes, E.A.; Raij, L. Thiazide diuretics, endothelial function, and vascular oxidative stress. J. Hypertens. 2008, 26, 494–500. [Google Scholar] [CrossRef]
- Itani, H.A.; McMaster, W.G.; Saleh, M.A.; Nazarewicz, R.R.; Mikolajczyk, T.P.; Kaszuba, A.M.; Konior, A.; Prejbisz, A.; Januszewicz, A.; Norlander, A.E.; et al. Activation of Human T Cells in Hypertension: Studies of Humanized Mice and Hypertensive Humans. Hypertension 2016, 68, 123–132. [Google Scholar] [CrossRef] [Green Version]
- Marvar, P.J.; Thabet, S.R.; Guzik, T.J.; Lob, H.E.; McCann, L.A.; Weyand, C.; Gordon, F.J.; Harrison, D.G. Central and peripheral mechanisms of T-lymphocyte activation and vascular inflammation produced by angiotensin II-induced hypertension. Circ. Res. 2010, 107, 263–270. [Google Scholar] [CrossRef] [Green Version]
- Sheikhi, A.; Jaberi, Y.; Esmaeilzadeh, A.; Khani, M.; Moosaeefard, M.; Shafaqatian, M. The effect of cardiovascular drugs on pro-inflammatory cytokine secretion and natural killer activity of peripheral blood mononuclear cells of patients with chronic heart failure in vitro. Pak. J. Biol. Sci. PJBS 2007, 10, 1580–1587. [Google Scholar] [CrossRef]
- Tuttolomondo, A.; Pinto, A.; Di Raimondo, D.; Corrao, S.; Di Sciacca, R.; Scaglione, R.; Caruso, C.; Licata, G. Changes in natriuretic peptide and cytokine plasma levels in patients with heart failure, after treatment with high dose of furosemide plus hypertonic saline solution (HSS) and after a saline loading. Nutr. Metab. Cardiovasc. Dis. NMCD 2011, 21, 372–379. [Google Scholar] [CrossRef] [PubMed]
- Tuttolomondo, A.; Di Raimondo, D.; Bellia, C.; Clemente, G.; Pecoraro, R.; Maida, C.; Simonetta, I.; Vassallo, V.; Di Bona, D.; Gulotta, E.; et al. Immune-Inflammatory and Metabolic Effects of High Dose Furosemide plus Hypertonic Saline Solution (HSS) Treatment in Cirrhotic Subjects with Refractory Ascites. PLoS ONE 2016, 11, e0165443. [Google Scholar] [CrossRef] [Green Version]
- Brennecke, A.; Villar, L.; Wang, Z.; Doyle, L.M.; Meek, A.; Reed, M.; Barden, C.; Weaver MD, D.F. Is Inhaled Furosemide a Potential Therapeutic for COVID-19? Am. J. Med. Sci. 2020, 360, 216–221. [Google Scholar] [CrossRef]
- TTröger, B.; Heidemann, M.; Osthues, I.; Knaack, D.; Gopel, W.; Herting, E.; Knobloch, J.K.-M.; Härtel, C. Modulation of S. epidermidis-induced innate immune responses in neonatal whole blood. J. Microbiol. Immunol. Infect. 2020, 53, 240–249. [Google Scholar] [CrossRef] [PubMed]
- Prandota, J. Furosemide: Progress in understanding its diuretic, anti-inflammatory, and bronchodilating mechanism of action, and use in the treatment of respiratory tract diseases. Am. J. Ther. 2002, 9, 317–328. [Google Scholar] [CrossRef]
- Xu, B.; Makris, A.; Thornton, C.; Ogle, R.; Horvath, J.S.; Hennessy, A. Antihypertensive drugs clonidine, diazoxide, hydralazine and furosemide regulate the production of cytokines by placentas and peripheral blood mononuclear cells in normal pregnancy. J. Hypertens. 2006, 24, 915–922. [Google Scholar] [CrossRef]
- Yuengsrigul, A.; Chin, T.W.; Nussbaum, E. Immunosuppressive and cytotoxic effects of furosemide on human peripheral blood mononuclear cells. Ann. Allergy Asthma Immunol. 1999, 83, 559–566. [Google Scholar] [CrossRef]
- Santoni, G.; Morelli, M.B.; Amantini, C.; Santoni, M.; Nabissi, M.; Marinelli, O.; Santoni, A. “Immuno-Transient Receptor Potential Ion Channels”: The Role in Monocyte- and Macrophage-Mediated Inflammatory Responses. Front. Immunol. 2019, 9, 1273. [Google Scholar] [CrossRef]
- Laskin, D.L.; Pendino, K.J. Macrophages and inflammatory mediators in tissue injury. Annu. Rev. Pharmacol. Toxicol. 1995, 35, 655–677. [Google Scholar] [CrossRef] [PubMed]
- Marcinkiewicz, J. Nitric oxide and antimicrobial activity of reactive oxygen intermediates. Immunopharmacology 1997, 37, 35–41. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kaneda, K.; Kasama, T. Immunopathogenesis of delayed-type hypersensitivity. Microsc. Res. Tech. 2001, 53, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Tsung, K.; Meko, J.B.; Peplinski, G.R.; Tsung, Y.L.; Norton, J.A. IL-12 induces T helper 1-directed antitumor response. J. Immunol. 1997, 158, 3359–3365. [Google Scholar]
- Suwanpradid, J.; Lee, M.J.; Hoang, P.; Kwock, J.; Floyd, L.P.; Smith, J.S.; Yin, Z.; Atwater, A.R.; Rajagopal, S.; Kedl, R.M.; et al. IL-27 Derived From Macrophages Facilitates IL-15 Production and T Cell Maintenance Following Allergic Hypersensitivity Responses. Front. Immunol. 2021, 12, 713304. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, D.H.; Igyártó, B.Z.; Gaspari, A.A. Early immune events in the induction of allergic contact dermatitis. Nat. Rev. Immunol. 2012, 12, 114–124. [Google Scholar] [CrossRef]
- Martin, S.F.; Dudda, J.C.; Bachtanian, E.; Lembo, A.; Liller, S.; Dürr, C.; Heimesaat, M.M.; Bereswill, S.; Fejer, G.; Vassileva, R.; et al. Toll-like receptor and IL-12 signaling control susceptibility to contact hypersensitivity. J. Exp. Med. 2008, 205, 2151–2162. [Google Scholar] [CrossRef]
- Martin, S.F.; Esser, P.R.; Weber, F.C.; Jakob, T.; Freudenberg, M.A.; Schmidt, M.; Goebeler, M. Mechanisms of chemical-induced innate immunity in allergic contact dermatitis. Allergy 2011, 66, 1152–1163. [Google Scholar] [CrossRef]
- Passero, L.F.D.; Cavallone, I.N.; Belda, W. Reviewing the Etiologic Agents, Microbe-Host Relationship, Immune Response, Diagnosis, and Treatment in Chromoblastomycosis. J. Immunol. Res. 2021, 2021, 9742832. [Google Scholar] [CrossRef]
- Askenase, P.W.; Bryniarski, K.; Paliwal, V.; Redegeld, F.; Groot Kormelink, T.; Kerfoot, S.; Hutchinson, A.T.; Van Loveren, H.; Campos, R.; Itakura, A.; et al. (2015) A subset of AID-dependent B-1a cells initiates hypersensitivity and pneumococcal pneumonia resistance. Ann. N. Y. Acad. Sci. 2015, 1362, 200–214. [Google Scholar] [CrossRef]
- Beheshti, F.; Akbari, H.R.; Baghcheghi, Y.; Mansouritorghabeh, F.; Mortazavi Sani, S.S.; Hosseini, M. Beneficial effects of angiotensin converting enzyme inhibition on scopolamine-induced learning and memory impairment in rats, the roles of brain-derived neurotrophic factor, nitric oxide and neuroinflammation. Clin. Exp. Hypertens. 2021, 43, 505–515. [Google Scholar] [CrossRef]
- Ardila, D.L.V.; Walsh, K.A.; Fifis, T.; Paolini, R.; Kastrappis, G.; Christophi, C.; Perini, M.V. Immunomodulatory effects of renin-angiotensin system inhibitors on T lymphocytes in mice with colorectal liver metastases. J. Immunother. Cancer 2020, 8, e000487. [Google Scholar] [CrossRef]
- Gan, Z.; Huang, D.; Jiang, J.; Li, Y.; Li, H.; Ke, Y. Captopril alleviates hypertension-induced renal damage, inflammation, and NF-κB activation. Braz. J. Med. Biol. Res. 2018, 51, e7338. [Google Scholar] [CrossRef]
- El-Ashmawy, N.E.; Khedr, N.F.; El-Bahrawy, H.A.; Hamada, O.B. Anti-inflammatory and Antioxidant Effects of Captopril Compared to Methylprednisolone in L-Arginine-Induced Acute Pancreatitis. Dig. Dis. Sci. 2018, 63, 1497–1505. [Google Scholar] [CrossRef] [PubMed]
- Demiralay, E.; Saglam, I.Y.; Ozdamar, E.N.; Sehirli, A.O.; Sener, G.; Saglam, E. nNOS expression in the brain of rats after burn and the effect of the ACE inhibitor captopril. Burn.: J. Int. Soc. Burn. Inj. 2013, 39, 897–904. [Google Scholar] [CrossRef]
- Abareshi, A.; Hosseini, M.; Beheshti, F.; Norouzi, F.; Khazaei, M.; Sadeghnia, H.R.; Boskabady, M.H.; Shafei, M.N.; Anaeigoudari, A. The effects of captopril on lipopolysaccharide induced learning and memory impairments and the brain cytokine levels and oxidative damage in rats. Life Sci. 2016, 167, 46–56. [Google Scholar] [CrossRef]
- Abdel-Wahab, B.A.; Metwally, M.E.; El-khawanki, M.M.; Hashim, A.M. Protective effect of captopril against clozapine-induced myocarditis in rats: Role of oxidative stress, proinflammatory cytokines and DNA damage. Chem.-Biol. Interact. 2014, 216, 43–52. [Google Scholar] [CrossRef]
- Drobná, M.; Misak, A.; Holland, T.; Kristek, F.; Grman, M.; Tomasova, L.; Berenyiova, A.; Cacanyiova, S.; Ondrias, K. Captopril partially decreases the effect of H(2)S on rat blood pressure and inhibits H(2)S-induced nitric oxide release from S-nitrosoglutathione. Physiol. Res. 2015, 64, 479–486. [Google Scholar] [CrossRef]
- Fouad, A.A.; Al-Mulhim, A.S.; Jresat, I.; Morsy, M.A. Protective effects of captopril in diabetic rats exposed to ischemia/reperfusion renal injury. J. Pharm. Pharmacol. 2012, 65, 243–252. [Google Scholar] [CrossRef]
- Asraf, K.; Torika, N.; Apte, R.N.; Fleisher-Berkovich, S. Microglial Activation Is Modulated by Captopril: In Vitro and In Vivo Studies. Front. Cell. Neurosci. 2018, 12, 116. [Google Scholar] [CrossRef] [PubMed]
- Nocito, C.; Lubinsky, C.; Hand, M.; Khan, S.; Patel, T.; Seliga, A.; Winfield, M.; Zuluaga-Ramirez, V.; Fernandes, N.; Shi, X.; et al. Centrally Acting Angiotensin-Converting Enzyme Inhibitor Suppresses Type I Interferon Responses and Decreases Inflammation in the Periphery and the CNS in Lupus-Prone Mice. Front. Immunol. 2020, 11, 573677. [Google Scholar] [CrossRef] [PubMed]
- Efrati, S.; Berman, S.; Hamad, R.A.; Siman-Tov, Y.; Ilgiyaev, E.; Maslyakov, I.; Weissgarten, J. Effect of captopril treatment on recuperation from ischemia/reperfusion-induced acute renal injury. Nephrol. Dial. Transplant. 2012, 27, 136–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.-Q.; Zhang, Q.; Chen, L.; Yin, C.-S.; Chen, P.; Tang, J.; Rong, R.; Li, T.-T.; Hu, L.-Q. Captopril inhibits maturation of dendritic cells and maintains their tolerogenic property in atherosclerotic rats. Int. Immunopharmacol. 2015, 28, 715–723. [Google Scholar] [CrossRef] [PubMed]
- Park, H.-S.; Han, A.; Yeo, H.-L.; Park, M.-J.; You, M.-J.; Choi, H.J.; Hong, C.-W.; Lee, S.-H.; Kim, S.H.; Kim, B.; et al. Chronic high dose of captopril induces depressive-like behaviors in mice: Possible mechanism of regulatory T cell in depression. Oncotarget 2017, 8, 72528–72543. [Google Scholar] [CrossRef] [Green Version]
- Boskabadi, J.; Askari, V.R.; Hosseini, M.; Boskabady, M.H. Immunomodulatory properties of captopril, an ACE inhibitor, on LPS-induced lung inflammation and fibrosis as well as oxidative stress. Inflammopharmacology 2019, 27, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Schindler, R.; Dinarello, C.A.; Koch, K.M. Angiotensin-converting-enzyme inhibitors suppress synthesis of tumour necrosis factor and interleukin 1 by human peripheral blood mononuclear cells. Cytokine 1995, 7, 526–533. [Google Scholar] [CrossRef]
- Constantinescu, C.S.; Goodman, D.B.; Ventura, E.S. Captopril and lisinopril suppress production of interleukin-12 by human peripheral blood mononuclear cells. Immunol. Lett. 1998, 62, 25–31. [Google Scholar] [CrossRef]
- Wang, Z.; Vilekar, P.; Huang, J.; Weaver, D.F. Furosemide as a Probe Molecule for the Treatment of Neuroinflammation in Alzheimer’s Disease. ACS Chem. Neurosci. 2020, 11, 4152–4168. [Google Scholar] [CrossRef]
- Soloperto, M.; Marini, M.; Brasca, C.; Fasoli, A.; Mattoli, S. The protective effect of frusemide on the generation of superoxide anions by human bronchial epithelial cells and pulmonary macrophages in vitro. Pulm. Pharmacol. 1991, 4, 80–84. [Google Scholar] [CrossRef]
- Wang, S.; Xiang, Y.-Y.; Ellis, R.; Wattie, J.; Feng, M.; Inman, M.D.; Lu, W.-Y. Effects of furosemide on allergic asthmatic responses in mice. Clin. Exp. Allergy 2011, 41, 1456–1467. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Thornton, C.; Makris, A.; Ogle, R.; Hennessy, A. Anti-hypertensive drugs alter cytokine production from preeclamptic placentas and peripheral blood mononuclear cells. Hypertens. Pregnancy 2007, 26, 343–356. [Google Scholar] [CrossRef]
- Muniz-Junqueira, M.I.; Mota, L.M.; Aires, R.B.; Junqueira, L.F. Digitalis inhibits and furosemide does not change the in vitro phagocytic function of neutrophils of healthy subjects. Int. Immunopharmacol. 2003, 3, 1439–1445. [Google Scholar] [CrossRef]
- Aloud, B.M.; Petkau, J.C.; Yu, L.; McCallum, J.; Kirby, C.; Netticadan, T.; Blewett, H. Effects of cyanidin 3-O-glucoside and hydrochlorothiazide on T-cell phenotypes and function in spontaneously hypertensive rats. Food Funct. 2020, 11, 8560–8572. [Google Scholar] [CrossRef] [PubMed]
- Khan, B.V.; Rahman, S.T.; Haque, T.; Merchant, N.; Bhaheetharan, S.; Harris, J.; Umar, K.; Wahi, J.; Ferdinand, K.C. Vascular effects of nebivolol added to hydrochlorothiazide in African Americans with hypertension and echocardiographic evidence of diastolic dysfunction: The NASAA study. J. Cardiovasc. Pharmacol. Ther. 2012, 17, 291–297. [Google Scholar] [CrossRef]
- Wang, Y.; Evangelista, S.; Liu, Y.; Zhang, M.-S. Beneficial effects of nebivolol and hydrochlorothiazide combination in spontaneously hypertensive rats. J. Int. Med. Res. 2013, 41, 603–617. [Google Scholar] [CrossRef] [Green Version]
- Rahman, S.T.; Lauten, W.B.; Khan, Q.A.; Navalkar, S.; Parthasarathy, S.; Khan, B.V. Effects of eprosartan versus hydrochlorothiazide on markers of vascular oxidation and inflammation and blood pressure (renin-angiotensin system antagonists, oxidation, and inflammation). Am. J. Cardiol. 2002, 89, 686–690. [Google Scholar] [CrossRef]
- Czajkowska, B.; Ptak, M.; Bobek, M.; Bryniarski, K.; Szczepanik, M. Different isoenzyme patterns of nonspecific esterases and the level of IL6 production as markers of macrophage functions. Folia Histochem. Et Cytobiol. 1995, 33, 111–115. [Google Scholar]
- Marzinzig, M.; Nussler, A.K.; Stadler, J.; Marzinzig, E.; Barthlen, W.; Nussler, N.C.; Beger, H.G.; Morris, S.M., Jr.; Bruckner, U.B. Improved methods to measure end products of nitric oxide in biological fluids: Nitrite, nitrate, and S-nitrosothiols. Nitric Oxide: Biol. Chem. 1997, 1, 177–189. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryniarski, P.; Nazimek, K.; Marcinkiewicz, J. Captopril Combined with Furosemide or Hydrochlorothiazide Affects Macrophage Functions in Mouse Contact Hypersensitivity Response. Int. J. Mol. Sci. 2022, 23, 74. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010074
Bryniarski P, Nazimek K, Marcinkiewicz J. Captopril Combined with Furosemide or Hydrochlorothiazide Affects Macrophage Functions in Mouse Contact Hypersensitivity Response. International Journal of Molecular Sciences. 2022; 23(1):74. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010074
Chicago/Turabian StyleBryniarski, Paweł, Katarzyna Nazimek, and Janusz Marcinkiewicz. 2022. "Captopril Combined with Furosemide or Hydrochlorothiazide Affects Macrophage Functions in Mouse Contact Hypersensitivity Response" International Journal of Molecular Sciences 23, no. 1: 74. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23010074