
Chemogenetic Depletion of Hypophysiotropic GnRH Neurons Does Not Affect Fertility in Mature Female Zebrafish


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Evaluation of Transgenic Line Tg(gnrh3:Gal4ff; UAS:nfsb-mCherry)
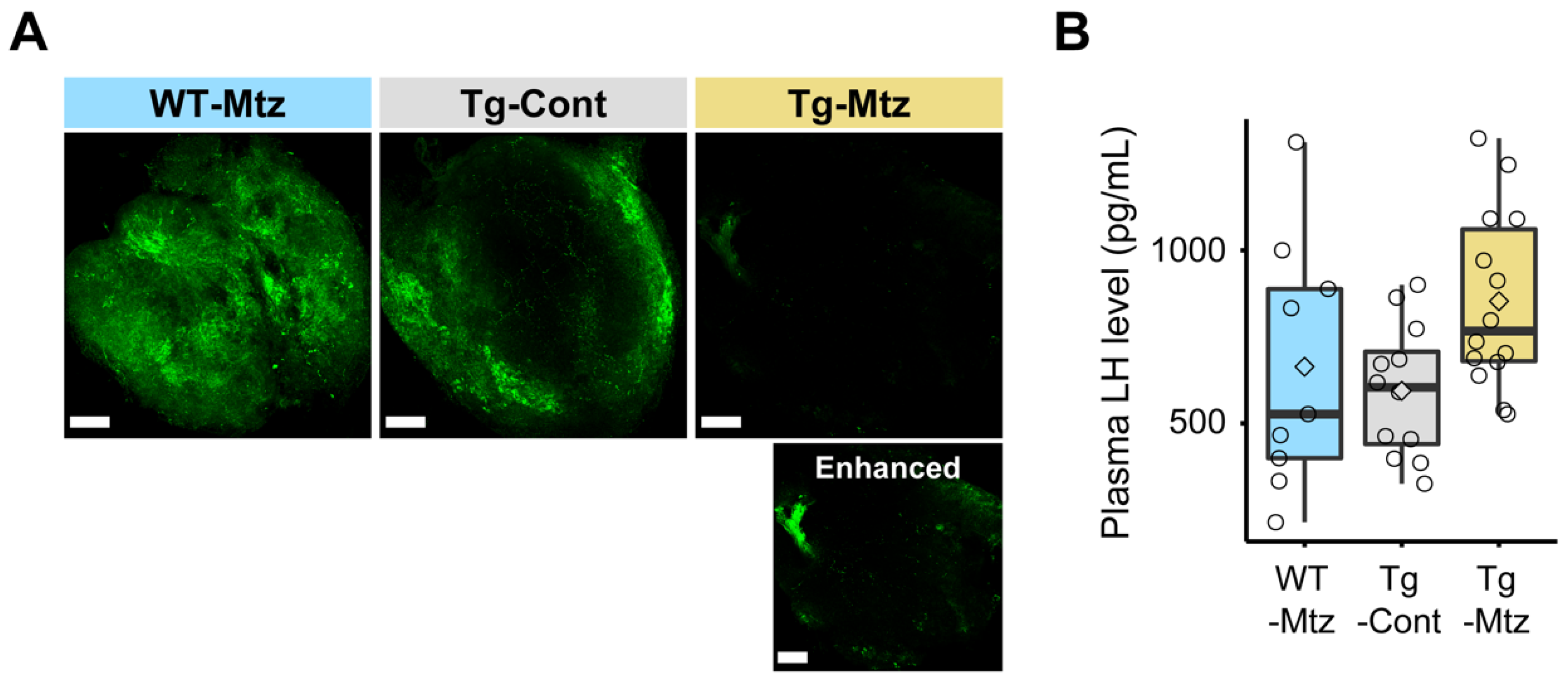
2.2. Attenuation of GnRH3 Neurons Is Verified
2.3. Plasma LH Levels Are Unchanged in GnRH3 Neuron-Ablated Females
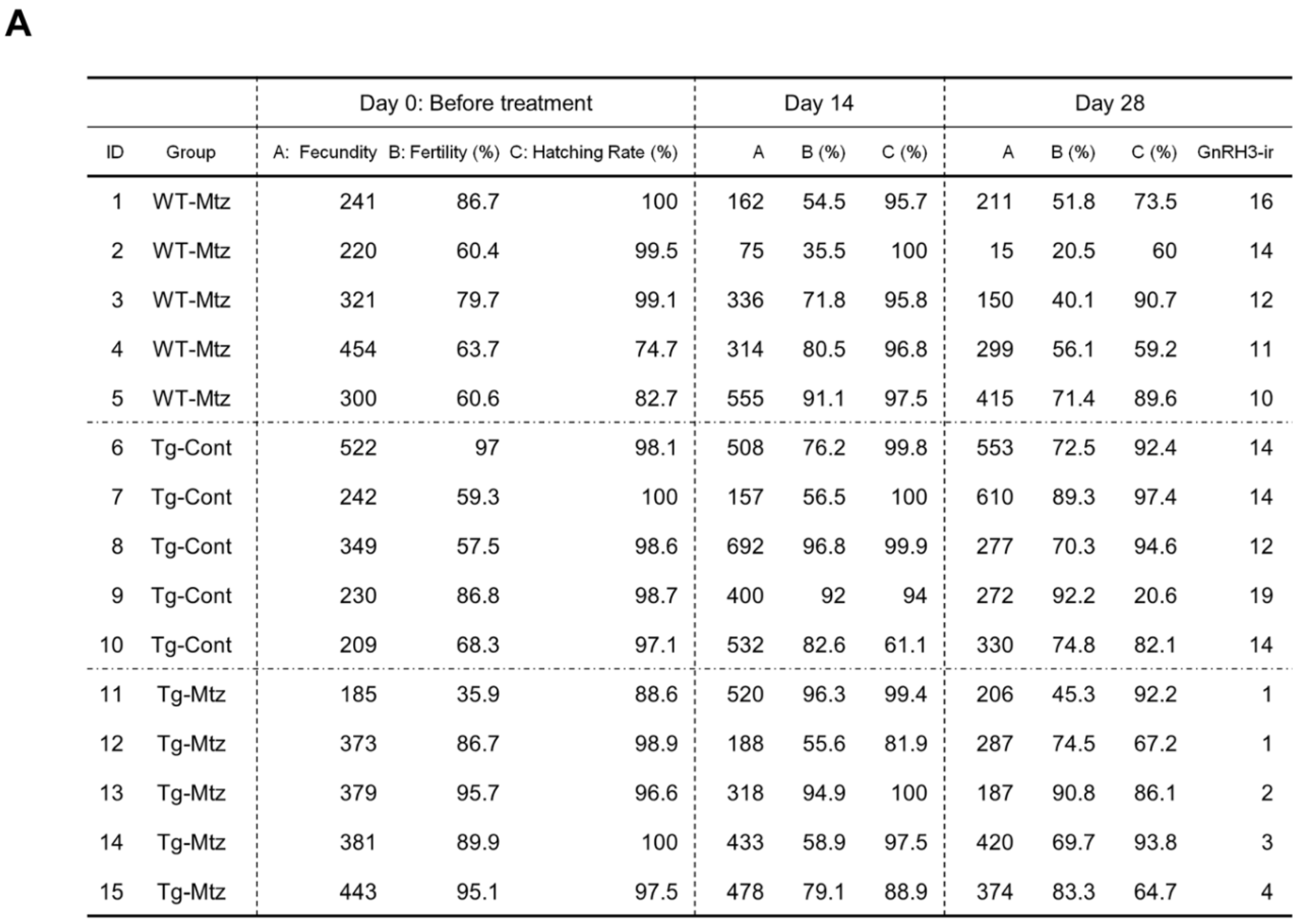
2.4. GnRH3 Neuron-Ablated Females Exhibit Normal Fertility
2.5. Pituitary GnRH2 Peptide Contents Are Unchanged in GnRH3 Neuron-Ablated Females
2.6. Ablated GnRH3 Neurons Regenerate after the Removal of Mtz during Early Development
2.7. Acute Ablation of GnRH3 Neurons in Adult Females Does Not Affect Fertility
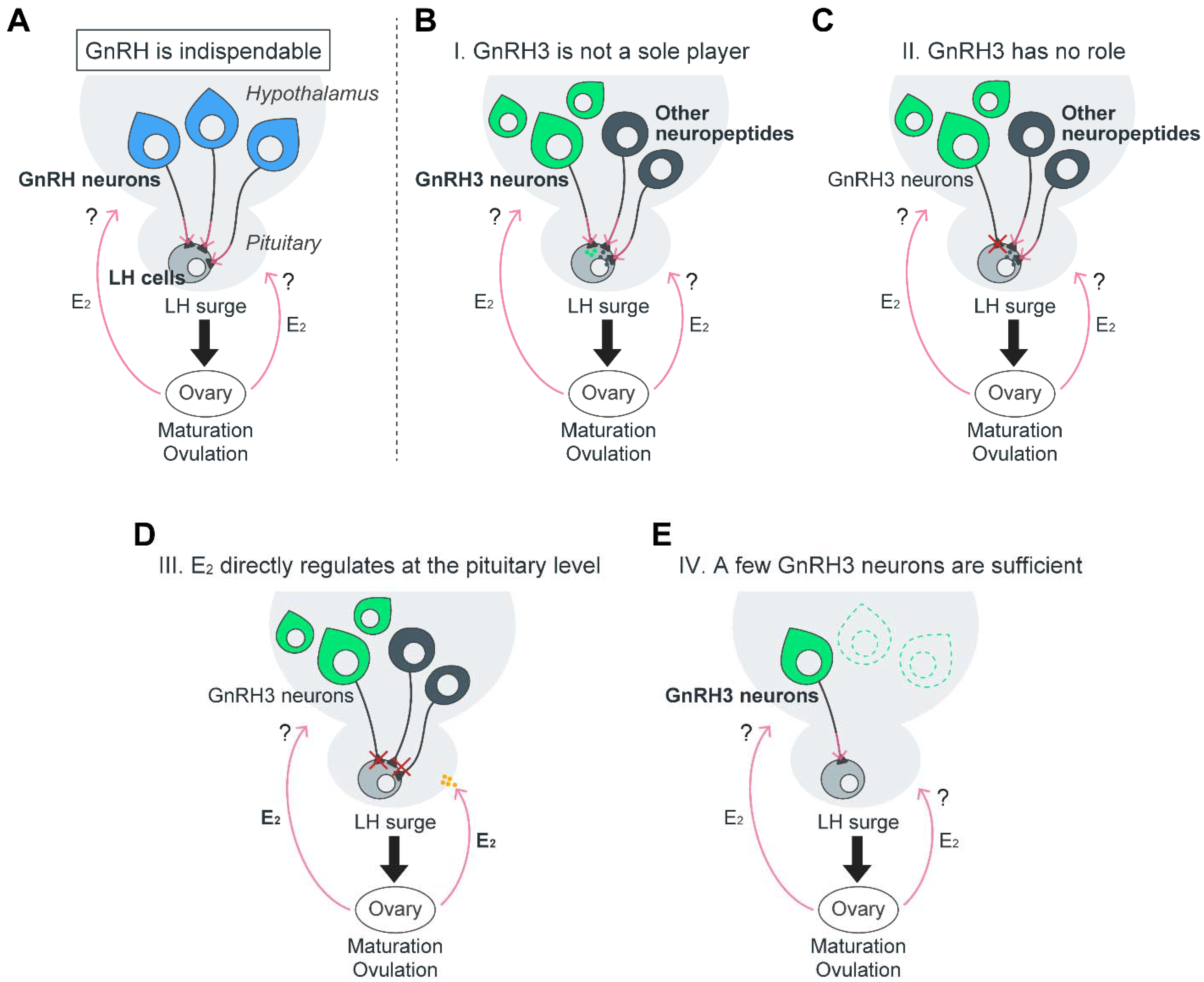
3. Discussion
3.1. GnRH1 and GnRH3 Functionally Differ: GnRH3 Likely Functions as a Neuromodulator and Not as an LH Secretagogue
3.2. The Effect of the Hypophysiotropic GnRH on Pituitary LH Secretion and lhb Expression Is Still Controversial in Female Zebrafish
3.3. Other Central and Peripheral Factors May Induce LH Secretion in Sexually Mature Female Zebrafish
3.4. A Small Amount of GnRH3 May Still Trigger LH Secretion
3.5. GnRH2 Likely Does Not Play a Role in Inducing Ovulation and Spawning in GnRH3 Neuron-Ablated Females
3.6. Summary
4. Material and Methods
4.1. Animal Husbandry
4.2. Generation of Transgenic Fish
4.3. Conditional Attenuation of GnRH3 Neurons in Adulthood
4.4. Conditional Attenuation of GnRH3 Neurons during Early Development
4.5. Characterization of Reproductive Performance
4.6. Confocal Imaging
4.7. Tissue Sampling
4.8. Ovarian Histology
4.9. Real-Time Quantitative PCR
4.10. Whole Mount Immunostaining
4.11. Protein Quantification by ELISA
4.12. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cattanach, B.; Iddon, C.A.; Charlton, H.; Chiappa, S.A.; Fink, G. Gonadotrophin-releasing hormone deficiency in a mutant mouse with hypogonadism. Nature 1977, 269, 338–340. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.-M.; de Guillebon, A.; Lang-Muritano, M.; Plummer, L.; Cerrato, F.; Tsiaras, S.; Gaspert, A.; Lavoie, H.B.; Wu, C.-H.; Crowley, W.F.; et al. GNRH1 mutations in patients with idiopathic hypogonadotropic hypogonadism. Proc. Natl. Acad. Sci. USA 2009, 106, 11703–11708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouligand, J.; Ghervan, C.; Tello, J.A.; Brailly-Tabard, S.; Salenave, S.; Chanson, P.; Lombès, M.; Millar, R.P.; Guiochon-Mantel, A.; Young, J. Isolated familial hypogonadotropic hypogonadism and a GNRH1 mutation. N. Engl. J. Med. 2009, 360, 2742–2748. [Google Scholar] [CrossRef] [PubMed]
- Herbison, A.E.; Porteous, R.; Pape, J.-R.; Mora, J.M.; Hurst, P.R. Gonadotropin-releasing hormone neuron requirements for puberty, ovulation, and fertility. Endocrinology 2007, 149, 597–604. [Google Scholar] [CrossRef]
- Takahashi, A.; Kanda, S.; Abe, T.; Oka, Y. Evolution of the hypothalamic-pituitary-gonadal axis regulation in vertebrates revealed by knockout medaka. Endocrinology 2016, 157, 3994–4002. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Nagahama, Y. Structural and functional evolution of gonadotropin-releasing hormone in vertebrates. Acta Physiol. 2008, 193, 3–15. [Google Scholar] [CrossRef]
- Zohar, Y.; Zmora, N.; Trudeau, V.L.; Muñoz-Cueto, J.A.; Golan, M. A half century of fish gonadotropin-releasing hormones: Breaking paradigms. J. Neuroendocrinol. 2021, e13069. [Google Scholar] [CrossRef]
- Muñoz-Cueto, J.A.; Zmora, N.; Paullada-Salmerón, J.A.; Marvel, M.; Mañanos, E.; Zohar, Y. The gonadotropin-releasing hormones: Lessons from fish. Gen. Comp. Endocrinol. 2020, 291, 113422. [Google Scholar] [CrossRef]
- Kah, O.; Lethimonier, C.; Lareyre, J.-J. Gonadotrophin-releasing hormone (GnRH) in the animal kingdom. J. Soc. Biol. 2004, 198, 53–60. [Google Scholar] [CrossRef]
- Wray, S.; Grant, P.; Gainer, H. Evidence That cells expressing luteinizing hormone-releasing hormone mRNA in the mouse are derived from progenitor cells in the olfactory placode. Proc. Natl. Acad. Sci. USA 1989, 86, 8132–8136. [Google Scholar] [CrossRef] [Green Version]
- Schwanzel-Fukuda, M.; Pfaff, D.W. Origin of luteinizing hormone-releasing hormone neurons. Nature 1989, 338, 161–164. [Google Scholar] [CrossRef]
- Trudeau, V.L.; Somoza, G.M. Multimodal hypothalamo-hypophysial communication in the vertebrates. Gen. Comp. Endocrinol. 2020, 293, 113475. [Google Scholar] [CrossRef]
- Millar, R.P. GnRH II and Type II GnRH rceptors. Trends. Endocrinol. Metab. 2003, 14, 35–43. [Google Scholar] [CrossRef]
- Millar, R.P. GnRHs and GnRH receptors. Anim. Reprod. Sci. 2005, 88, 5–28. [Google Scholar] [CrossRef]
- Okubo, K.; Aida, K. Gonadotropin-releasing hormones (GnRHs) in a primitive teleost, the arowana: Phylogenetic evidence that three paralogous lineages of GnRH occurred prior to the emergence of teleosts. Gen. Comp. Endocrinol. 2001, 124, 125–133. [Google Scholar] [CrossRef]
- Chen, C.-C.; Fernald, R. GnRH and GnRH receptors: Distribution, function and evolution. J. Fish Biol. 2008, 73, 1099–1120. [Google Scholar] [CrossRef]
- Aguillon, R.; Batut, J.; Subramanian, A.; Madelaine, R.; Dufourcq, P.; Schilling, T.F.; Blader, P. Cell-type heterogeneity in the early zebrafish olfactory epithelium is generated from progenitors within preplacodal ectoderm. eLife 2018, 7, e32041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palevitch, O.; Kight, K.; Abraham, E.; Wray, S.; Zohar, Y.; Gothilf, Y. Ontogeny of the GnRH systems in zebrafish brain: In situ hybridization and promoter-reporter expression analyses in intact animals. Cell Tissue Res. 2007, 327, 313–322. [Google Scholar] [CrossRef]
- Abraham, E.; Palevitch, O.; Ijiri, S.; Du, S.; Gothilf, Y.; Zohar, Y. Early development of forebrain gonadotrophin-releasing hormone (GnRH) neurones and the role of GnRH as an autocrine migration factor. J. Neuroendocrinol. 2008, 20, 394–405. [Google Scholar] [CrossRef]
- Golan, M.; Zelinger, E.; Zohar, Y.; Levavi-Sivan, B. Architecture of GnRH-gonadotrope-vasculature reveals a dual mode of gonadotropin regulation in fish. Endocrinology 2015, 156, 4163–4173. [Google Scholar] [CrossRef] [Green Version]
- Golan, M.; Boulanger-Weill, J.; Pinot, A.; Fontanaud, P.; Faucherre, A.; Gajbhiye, D.S.; Hollander-Cohen, L.; Fiordelisio-Coll, T.; Martin, A.O.; Mollard, P. Synaptic communication mediates the assembly of a self-organizing circuit that controls reproduction. Sci. Adv. 2021, 7, eabc8475. [Google Scholar] [CrossRef]
- Abraham, E.; Palevitch, O.; Gothilf, Y.; Zohar, Y. Targeted Gonadotropin-releasing hormone-3 neuron ablation in zebrafish: Effects on neurogenesis, neuronal migration, and reproduction. Endocrinology 2009, 151, 332–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spicer, O.; Wong, T.-T.; Zmora, N.; Zohar, Y. Targeted mutagenesis of the hypophysiotropic Gnrh3 in zebrafish (Danio rerio) reveals no effects on reproductive performance. PLoS ONE 2016, 11, e0158141. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Tang, H.; Xie, R.; Li, S.; Liu, X.; Lin, H.; Zhang, Y.; Cheng, C.H.K. Genetic evidence for multifactorial control of the reproductive axis in zebrafish. Endocrinology 2017, 158, 604–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, A.; Kontarakis, Z.; Gerri, C.; Nolte, H.; Hölper, S.; Krüger, M.; Stainier, D.Y. Genetic compensation induced by deleterious mutations but not gene knockdowns. Nature 2015, 524, 230–233. [Google Scholar] [CrossRef]
- Diss, G.; Ascencio, D.; DeLuna, A.; Landry, C.R. Molecular mechanisms of paralogous compensation and the robustness of cellular networks. J. Exp. Zool. B Mol. Dev. Evol. 2014, 322, 488–499. [Google Scholar] [CrossRef]
- Tsokas, P.; Hsieh, C.; Yao, Y.; Lesburguères, E.; Wallace, E.; Tcherepanov, A.; Jothianandan, D.; Hartley, B.; Pan, L.; Rivard, B.; et al. Compensation for PKMζ in long-term potentiation and spatial long-term memory in mutant mice. eLife 2016, 5, e14846. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, S.; Tsuruta, S.; Okada, K.; Saegusa, A.; Miyagaki, Y.; Baba, T. Functional compensation for the loss of testis-specific poly(A)-binding protein, PABPC2, during mouse spermatogenesis. J. Reprod. Dev. 2016, 62, 305–310. [Google Scholar] [CrossRef] [Green Version]
- Nagy, J.I.; Rash, J.E. Cx36, Cx43 and Cx45 in Mouse and rat cerebellar cortex: Species-specific expression, compensation in Cx36 null mice and co-localization in neurons vs. glia. Eur. J. Neurosci. 2017, 46, 1790–1804. [Google Scholar] [CrossRef]
- Spergel, D.J. Modulation of gonadotropin-releasing hormone neuron activity and secretion in mice by non-peptide neurotransmitters, gasotransmitters, and gliotransmitters. Front. Endocrinol. 2019, 10, 329. [Google Scholar] [CrossRef]
- Lee, K.; Porteous, R.; Campbell, R.E.; Lüscher, B.; Herbison, A.E. Knockdown of GABAA receptor signaling in GnRH neurons has minimal effects upon fertility. Endocrinology 2010, 151, 4428–4436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerikan, B.; Shaheen, R.; Colo, G.P.; Gläßer, C.; Hata, S.; Knobeloch, K.-P.; Alkuraya, F.S.; Fässler, R.; Schiebel, E. Cell-intrinsic adaptation arising from chronic ablation of a key Rho GTPase regulator. Dev. Cell 2016, 39, 28–43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsumoto, S.; Yamazaki, C.; Masumoto, K.; Nagano, M.; Naito, M.; Soga, T.; Hiyama, H.; Matsumoto, M.; Takasaki, J.; Kamohara, M.; et al. Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc. Natl. Acad. Sci. USA 2006, 103, 4140–4145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, H.M.; Mellon, P.L. A Small population of hypothalamic neurons govern fertility: The critical role of VAX1 in GnRH neuron development and Fertility Maintenance. Neurosci. Commun. 2016, 2, e1373. [Google Scholar]
- Larder, R.; Kimura, I.; Meadows, J.; Clark, D.D.; Mayo, S.; Mellon, P.L. Gene dosage of Otx2 is important for fertility in male mice. Mol. Cell. Endocrinol. 2013, 377, 16–22. [Google Scholar] [CrossRef] [Green Version]
- Larder, R.; Clark, D.D.; Miller, N.L.G.; Mellon, P.L. Hypothalamic dysregulation and infertility in mice lacking the homeodomain protein Six6. J. Neurosci. 2011, 31, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Givens, M.L.; Rave-Harel, N.; Goonewardena, V.D.; Kurotani, R.; Berdy, S.E.; Swan, C.H.; Rubenstein, J.L.R.; Robert, B.; Mellon, P.L. Developmental regulation of gonadotropin-releasing hormone gene expression by the MSX and DLX homeodomain protein families. J. Biol. Chem. 2005, 280, 19156–19165. [Google Scholar] [CrossRef] [Green Version]
- Diaczok, D.; DiVall, S.; Matsuo, I.; Wondisford, F.E.; Wolfe, A.M.; Radovick, S. Deletion of Otx2 in GnRH neurons results in a mouse model of hypogonadotropic hypogonadism. Mol. Endocrinol. 2011, 25, 833–846. [Google Scholar] [CrossRef] [Green Version]
- Felmer, R.; Cui, W.; Clark, A. Inducible ablation of adipocytes in adult transgenic mice expressing the E. coli nitroreductase gene. J. Endocrinol. 2002, 175, 487–498. [Google Scholar] [CrossRef] [Green Version]
- Curado, S.; Anderson, R.M.; Jungblut, B.; Mumm, J.; Schroeter, E.; Stainier, D. Conditional targeted cell ablation in zebrafish: A new tool for regeneration studies. Dev. Dyn. 2007, 236, 1025–1035. [Google Scholar] [CrossRef]
- Curado, S.; Stainier, D.Y.; Anderson, R.M. Nitroreductase-mediated cell/tissue ablation in zebrafish: A spatially and temporally controlled ablation method with applications in developmental and regeneration studies. Nat. Protoc. 2008, 3, 948–954. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisharath, H.; Parsons, M.J. Zebrafish, methods and protocols. Methods Mol. Biol. 2009, 546, 133–143. [Google Scholar]
- Yamamoto, N.; Oka, Y.; Kawashima, S. Lesions of gonadotropin-releasing hormone-immunoreactive terminal nerve cells: Effects on the reproductive behavior of male dwarf gouramis. Neuroendocrinology 1997, 65, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Wojtowicz, J.L.; Malin, J.H.; Huang, T.; Lee, E.B.; Chen, Z. GnRH-mediated olfactory and visual inputs promote mating-like behaviors in male zebrafish. PLoS ONE 2017, 12, e0174143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohno, S. Evolution by Gene Duplication; Springer: Berlin, Germany; Heidelberg, Germany, 1970. [Google Scholar]
- Bassi, I.; Luzzani, F.; Marelli, F.; Vezzoli, V.; Cotellessa, L.; Prober, D.A.; Persani, L.; Gothilf, Y.; Bonomi, M. Prokineticin receptor 2 affects GnRH3 neuron ontogeny but not fertility in zebrafish. Sci. Rep. 2020, 10, 7632. [Google Scholar] [CrossRef]
- Paredes, J.; Cowan, M.; López-Olmeda, J.; Muñoz-Cueto, J.A.; Sánchez-Vázquez, F. Daily rhythms of expression in reproductive genes along the brain-pituitary-gonad axis and liver of zebrafish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2019, 231, 158–169. [Google Scholar] [CrossRef]
- Chu, L.; Li, J.; Liu, Y.; Hu, W.; Cheng, C.H.K. Targeted gene disruption in zebrafish reveals noncanonical functions of LH signaling in reproduction. Mol. Endocrinol. 2014, 28, 1785–1795. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, S.; Zmora, N.; Levavi-Sivan, B.; Zohar, Y. Vasoactive intestinal peptide indirectly elicits pituitary LH secretion independent of GnRH in female zebrafish. Endocrinology 2022, 163, bqab264. [Google Scholar] [CrossRef]
- Biran, J.; Palevitch, O.; Ben-Dor, S.; Levavi-Sivan, B. Neurokinin Bs and neurokinin B receptors in zebrafish-potential role in controlling fish reproduction. Proc. Natl. Acad. Sci. USA 2012, 109, 10269–10274. [Google Scholar] [CrossRef] [Green Version]
- Tello, J.A.; Wu, S.; Rivier, J.E.; Sherwood, N.M. Four functional GnRH receptors in zebrafish: Analysis of structure, signaling, synteny and phylogeny. Integr. Comp. Biol. 2008, 48, 570–587. [Google Scholar] [CrossRef] [Green Version]
- Chang, J.P.; Peter, R.E. Effects of pimozide and des Gly10, [d-Ala6] luteinizing hormone-releasing hormone ethylamide on serum gonadotropin concentrations, germinal vesicle migration, and ovulation in female goldfish, Carassius auratus. Gen. Comp. Endocrinol. 1983, 52, 30–37. [Google Scholar] [CrossRef]
- Lin, H.-R.; Kraak, G.V.D.; Zhou, X.-J.; Liang, J.-Y.; Peter, R.E.; Rivier, J.E.; Vale, W.W. Effects of [d-Arg6, Trp7, Leu8, Pro9 NEt]-luteinizing hormone-releasing hormone (SGnRH-A) and [d-Ala6, Pro9NEt]-luteinizing hormone-releasing hormone (LHRH-A), in combination with pimozide or domperidone, on gonadotropin release and ovulation in the Chinese loach and common carp. Gen. Comp. Endocrinol. 1988, 69, 31–40. [Google Scholar]
- Fontaine, R.; Affaticati, P.; Yamamoto, K.; Jolly, C.; Bureau, C.; Baloche, S.; Gonnet, F.; Vernier, P.; Dufour, S.; Pasqualini, C. Dopamine inhibits reproduction in female zebrafish (Danio rerio) via three pituitary D2 receptor subtypes. Endocrinology 2013, 154, 807–818. [Google Scholar] [CrossRef] [Green Version]
- Lin, S.-W.; Ge, W. Differential regulation of gonadotropins (FSH and LH) and growth hormone (GH) by neuroendocrine, endocrine, and paracrine factors in the zebrafish—An in vitro approach. Gen. Comp. Endocrinol. 2009, 160, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Trudeau, V.L. Facing the challenges of neuropeptide gene knockouts: Why do they not inhibit reproduction in adult teleost fish? Front. Neurosci. 2018, 12, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.; Wang, M.; Ji, G.; Yang, S.; Zhang, S.; Liu, Z. Demonstration of a functional kisspeptin/kisspeptin receptor system in amphioxus with implications for origin of neuroendocrine regulation. Endocrinology 2017, 158, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.; Zhang, W.S.; Lu, C.; Tao, B.; Chen, L.; Hu, W.; Trudeau, V.L. Targeted mutation of secretogranin-2 disrupts sexual behavior and reproduction in zebrafish. Proc. Natl. Acad. Sci. USA 2020, 117, 12772–12783. [Google Scholar] [CrossRef]
- Moenter, S.M.; Silveira, M.A.; Wang, L.; Adams, C. Central aspects of systemic oestradiol negative-and positive-feedback on the reproductive neuroendocrine system. J. Neuroendocrinol. 2020, 32, e12724. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Liu, Y.; Luo, D.; Ogawa, S.; Yin, Y.; Li, S.; Zhang, Y.; Hu, W.; Parhar, I.S.; Lin, H.; et al. The Kiss/Kissr systems are dispensable for zebrafish reproduction: Evidence from gene knockout studies. Endocrinology 2015, 156, 589–599. [Google Scholar] [CrossRef]
- Nakajo, M.; Kanda, S.; Karigo, T.; Takahashi, A.; Akazome, Y.; Uenoyama, Y.; Kobayashi, M.; Oka, Y. Evolutionally conserved function of kisspeptin neuronal system is non-reproductive regulation as revealed by non-mammalian study. Endocrinology 2018, 159, 163–183. [Google Scholar] [CrossRef]
- Kanda, S. Evolution of the regulatory mechanism for hypothalamic-pituitary-gonadal axis in vertebrates—Hypothesis from a comparative view. Gen. Comp. Endocrinol. 2019, 284, 113075. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Tang, H.; Chen, Y.; Yin, Y.; Ogawa, S.; Liu, M.; Guo, Y.; Qi, X.; Liu, Y.; Parhar, I.S.; et al. Estrogen directly stimulates LHb expression at the pituitary level during puberty in female zebrafish. Mol. Cell. Endocrinol. 2018, 461, 1–11. [Google Scholar] [CrossRef]
- Neal-Perry, G.; Nejat, E.; Dicken, C. The neuroendocrine physiology of female reproductive aging: An update. Maturitas 2010, 67, 34–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.; Shu, J.; Neal-Perry, G. Intracerebroventricular infusion of vasoactive intestinal peptide rescues the luteinizing hormone surge in middle-aged female rats. Front. Endocrinol. 2012, 3, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansano, N.S.; Paradele, R.S.; Bohlen, T.M.; Zanardi, I.M.; Chaves, F.M.; Silveira, M.A.; Tavares, M.R.; Donato Jr., J.; Frazao, R. Vasoactive intestinal peptide exerts an excitatory effect on hypothalamic kisspeptin neurons during estrogen negative feedback. Mol. Cell. Endocrinol. 2022, 542, 111532. [Google Scholar] [CrossRef]
- Halpern, M.E.; Rhee, J.; Goll, M.G.; Akitake, C.M.; Parsons, M.; Leach, S.D. Gal4/UAS transgenic tools and their application to zebrafish. Zebrafish 2008, 5, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Feng, K.; Cui, X.; Song, Y.; Tao, B.; Chen, J.; Wang, J.; Liu, S.; Sun, Y.; Zhu, Z.; Trudeau, V.L.; et al. Gnrh3 regulates PGC Proliferation and sex differentiation in developing zebrafish. Endocrinology 2020, 161, bqz024. [Google Scholar] [CrossRef] [PubMed]
- Gamble, J.A.; Karunadasa, D.K.; Pape, J.-R.; Skynner, M.J.; Todman, M.G.; Bicknell, R.J.; Allen, J.P.; Herbison, A.E. Disruption of ephrin signaling associates with disordered axophilic migration of the gonadotropin-releasing hormone neurons. J. Neurosci. 2005, 25, 3142–3150. [Google Scholar] [CrossRef] [PubMed]
- Marvel, M.; Levavi-Sivan, B.; Wong, T.-T.; Zmora, N.; Zohar, Y. Gnrh2 maintains reproduction in fasting zebrafish through dynamic neuronal projection changes and regulation of gonadotropin synthesis, oogenesis, and reproductive behaviors. Sci. Rep. 2021, 11, 6657. [Google Scholar] [CrossRef]
- Marvel, M.; Spicer, O.; Wong, T.-T.; Zmora, N.; Zohar, Y. Knockout of Gnrh2 in zebrafish (Danio rerio) reveals its roles in regulating feeding behavior and oocyte quality. Gen. Comp. Endocrinol. 2019, 280, 15–23. [Google Scholar] [CrossRef]
- Davison, J.M.; Akitake, C.M.; Goll, M.G.; Rhee, J.M.; Gosse, N.; Baier, H.; Halpern, M.E.; Leach, S.D.; Parsons, M.J. Transactivation from Gal4-VP16 transgenic insertions for tissue-specific cell labeling and ablation in zebrafish. Dev. Biol. 2007, 304, 811–824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horstick, E.J.; Jordan, D.C.; Bergeron, S.A.; Tabor, K.M.; Serpe, M.; Feldman, B.; Burgess, H.A. Increased functional protein expression using nucleotide sequence features enriched in highly expressed genes in zebrafish. Nucleic Acids Res. 2015, 43, e48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, Y.A.; Woods, D.C.; Wood, A.W. A transgenic zebrafish model of targeted oocyte ablation and de novo oogenesis. Dev. Dyn. 2011, 240, 1929–1937. [Google Scholar] [CrossRef] [PubMed]
- Cavanaugh, A.M.; Huang, J.; Chen, J.-N. Two developmentally distinct populations of neural crest cells contribute to the zebrafish heart. Dev. Biol. 2015, 404, 103–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Feng, Y.; Wang, F.; Dong, X.; Jiang, L.; Liu, C.; Zhao, Q.; Li, K. Generation of all-male-like sterile zebrafish by eliminating primordial germ cells at early development. Sci. Rep. 2018, 8, 1834. [Google Scholar] [CrossRef] [Green Version]
- Elbaz, I.; Yelin-Bekerman, L.; Nicenboim, J.; Vatine, G.; Appelbaum, L. Genetic ablation of hypocretin neurons alters behavioral state transitions in zebrafish. J. Neurosci. 2012, 32, 12961–12972. [Google Scholar] [CrossRef] [Green Version]
- Chung, A.-Y.; Kim, P.-S.; Kim, S.; Kim, E.; Kim, D.; Jeong, I.; Kim, H.-K.; Ryu, J.-H.; Kim, C.-H.; Choi, J.; et al. Generation of demyelination models by targeted ablation of oligodendrocytes in the zebrafish CNS. Mol. Cells 2013, 36, 82–87. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Zang, L.; Shimada, Y.; Nishimura, Y.; Tanaka, T.; Nishimura, N. Repeated blood collection for blood tests in adult zebrafish. J. Vis. Exp. 2015, 102, e53272. [Google Scholar] [CrossRef]
- Hollander-Cohen, L.; Golan, M.; Aizen, J.; Shpilman, M.; Levavi-Sivan, B. Characterization of carp gonadotropins: Structure, annual profile, and carp and zebrafish pituitary topographic organization. Gen. Comp. Endocrinol. 2018, 264, 28–38. [Google Scholar] [CrossRef]
- Holland, M.C.H.; Gothilf, Y.; Meiri, I.; King, J.A.; Okuzawa, K.; Elizur, A.; Zohar, Y. Levels of the native forms of gnrh in the pituitary of the gilthead seabream, Sparus aurata, at several characteristic stages of the gonadal cycle. Gen. Comp. Endocrinol. 1998, 112, 394–405. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing, Version 4.1.1; R Foundation for Statistical Computing: Vienna, Austria, 2021. Available online: https://www.R-project.org/ (accessed on 1 September 2021).






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tanaka, S.; Zmora, N.; Levavi-Sivan, B.; Zohar, Y. Chemogenetic Depletion of Hypophysiotropic GnRH Neurons Does Not Affect Fertility in Mature Female Zebrafish. Int. J. Mol. Sci. 2022, 23, 5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105596
Tanaka S, Zmora N, Levavi-Sivan B, Zohar Y. Chemogenetic Depletion of Hypophysiotropic GnRH Neurons Does Not Affect Fertility in Mature Female Zebrafish. International Journal of Molecular Sciences. 2022; 23(10):5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105596
Chicago/Turabian StyleTanaka, Sakura, Nilli Zmora, Berta Levavi-Sivan, and Yonathan Zohar. 2022. "Chemogenetic Depletion of Hypophysiotropic GnRH Neurons Does Not Affect Fertility in Mature Female Zebrafish" International Journal of Molecular Sciences 23, no. 10: 5596. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105596