
4-Pyridone-3-carboxamide-1-β-D-ribonucleoside (4PYR)—A Novel Oncometabolite Modulating Cancer-Endothelial Interactions in Breast Cancer Metastasis

 , , , , ,
, , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
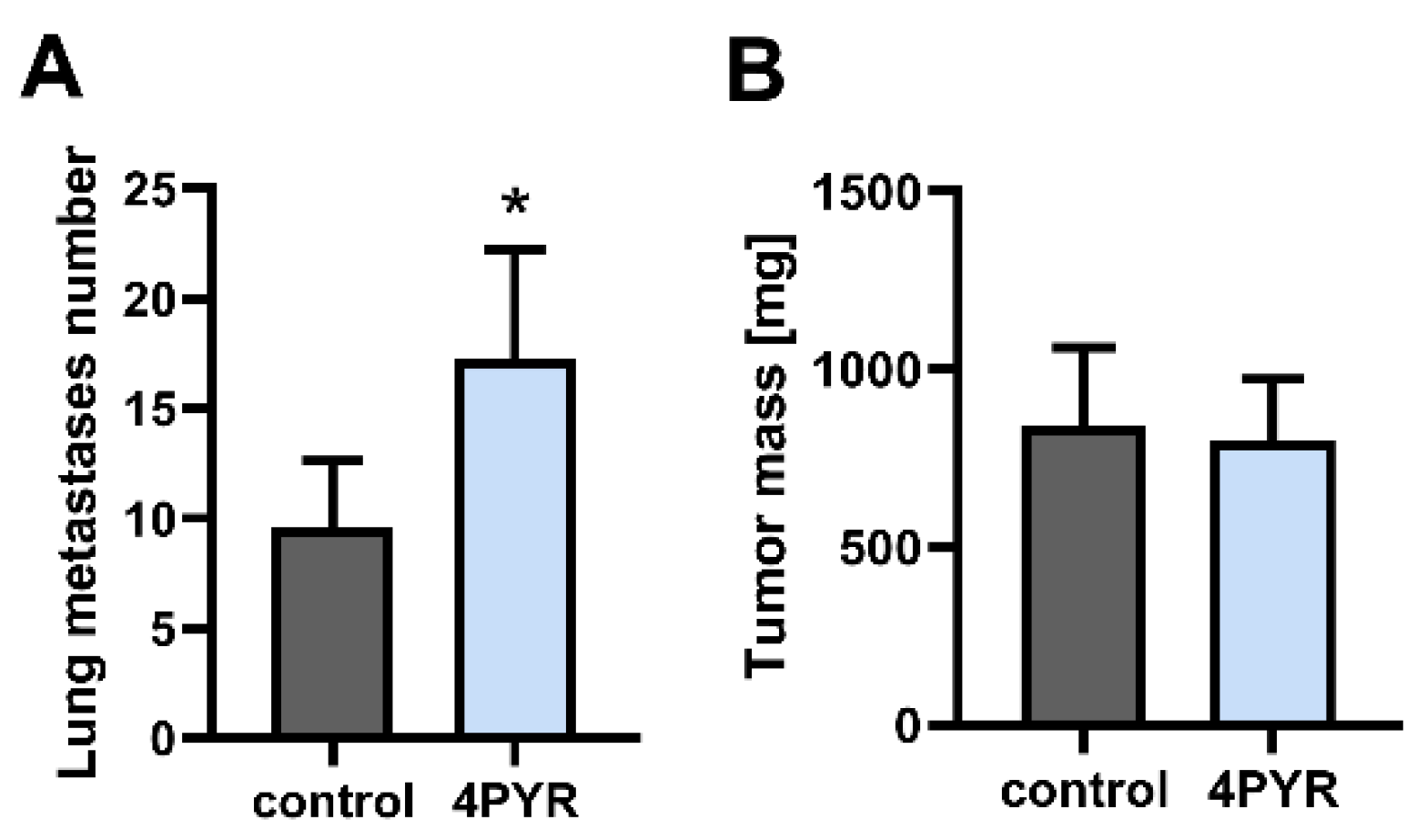
2.1. 4PYR Enhances Lung Metastasis Formation but Does Not Affect Primary Tumor Growth
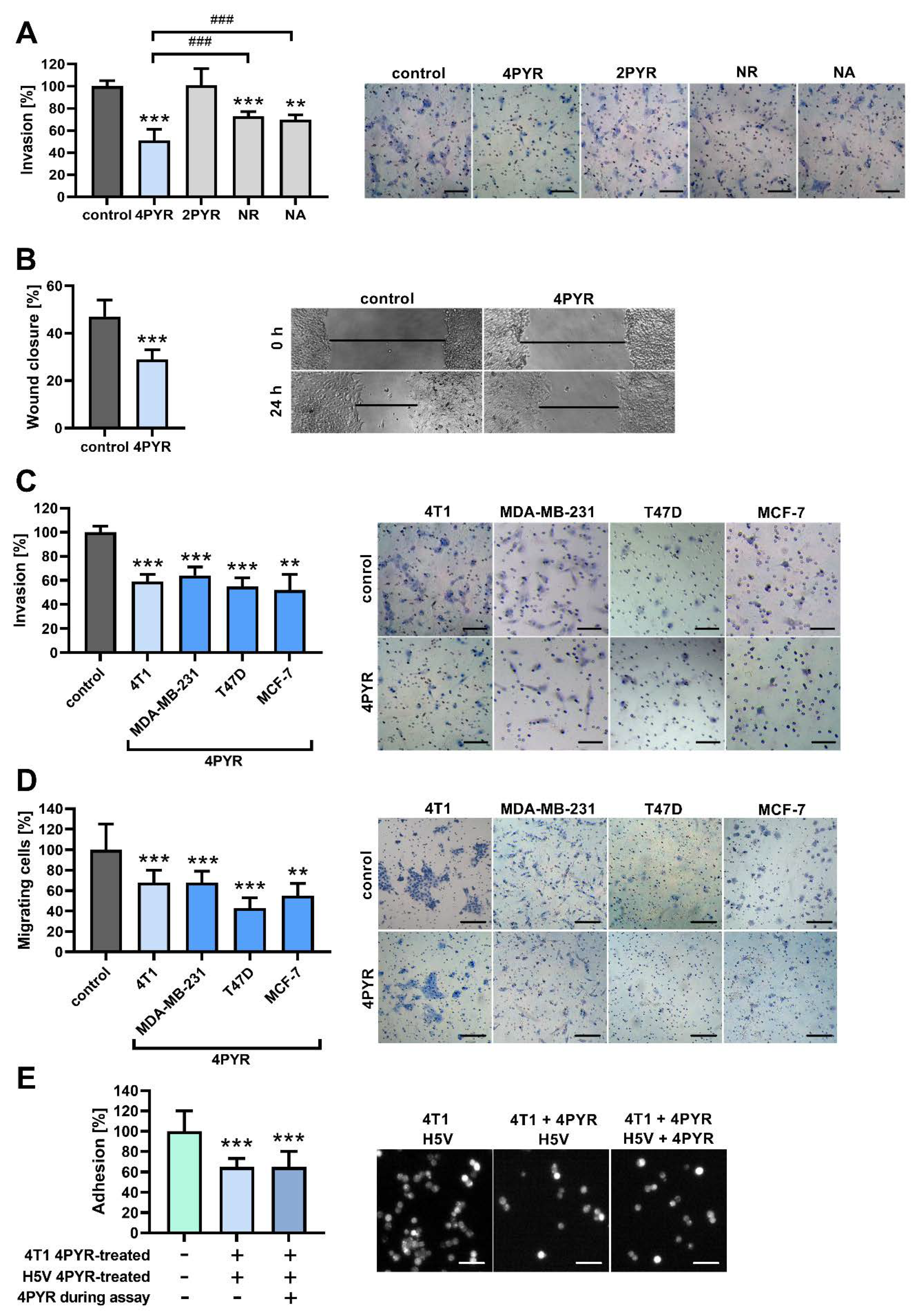
2.2. 4PYR Inhibits the Invasive Potential of 4T1 Breast Cancer Cells In Vitro
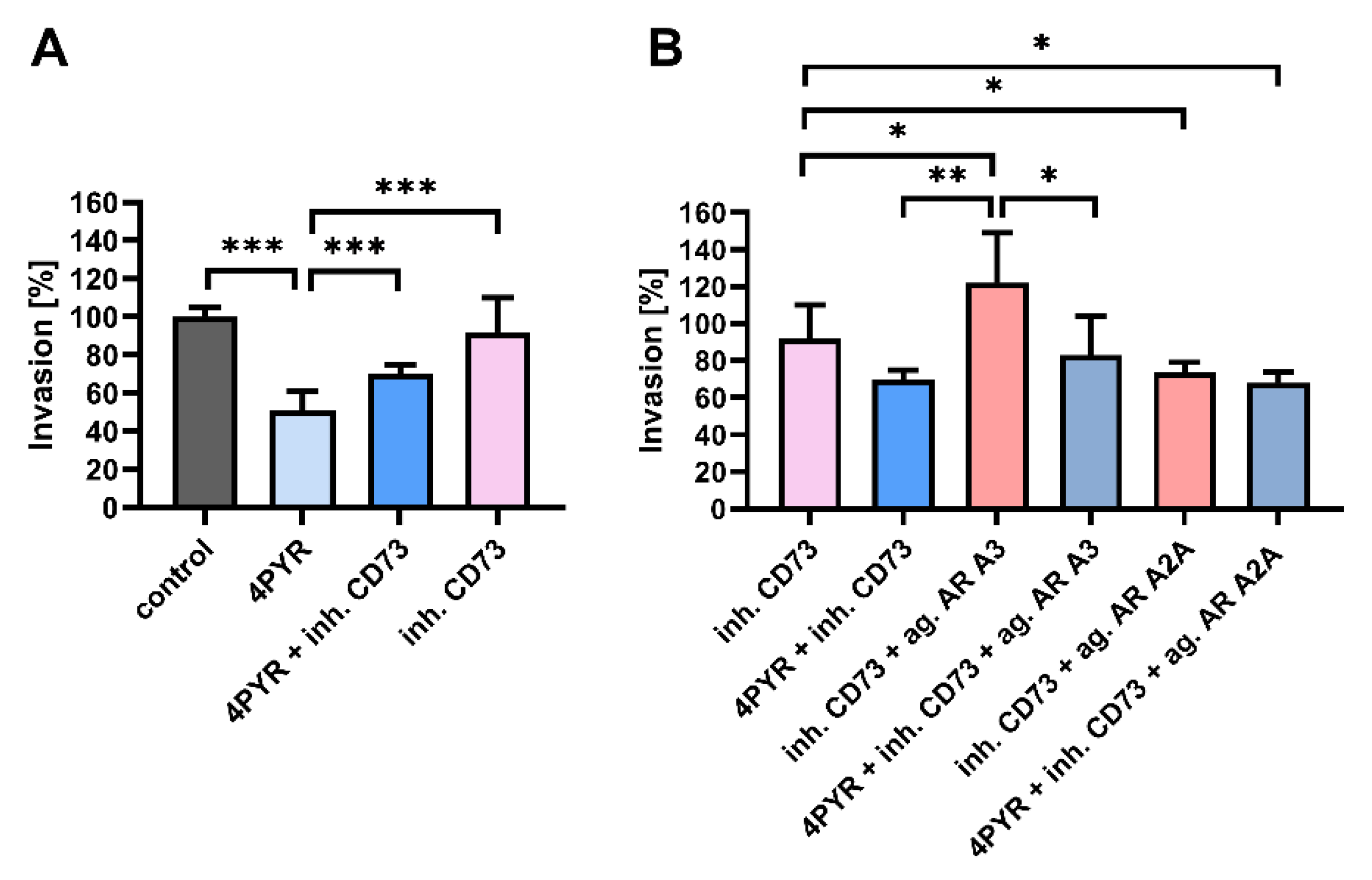
2.3. 4PYR-Induced Modulation of the Invasive Potential of 4T1 Cells Is Dependent on CD73 Activity and Mediated through Adenosine Receptors
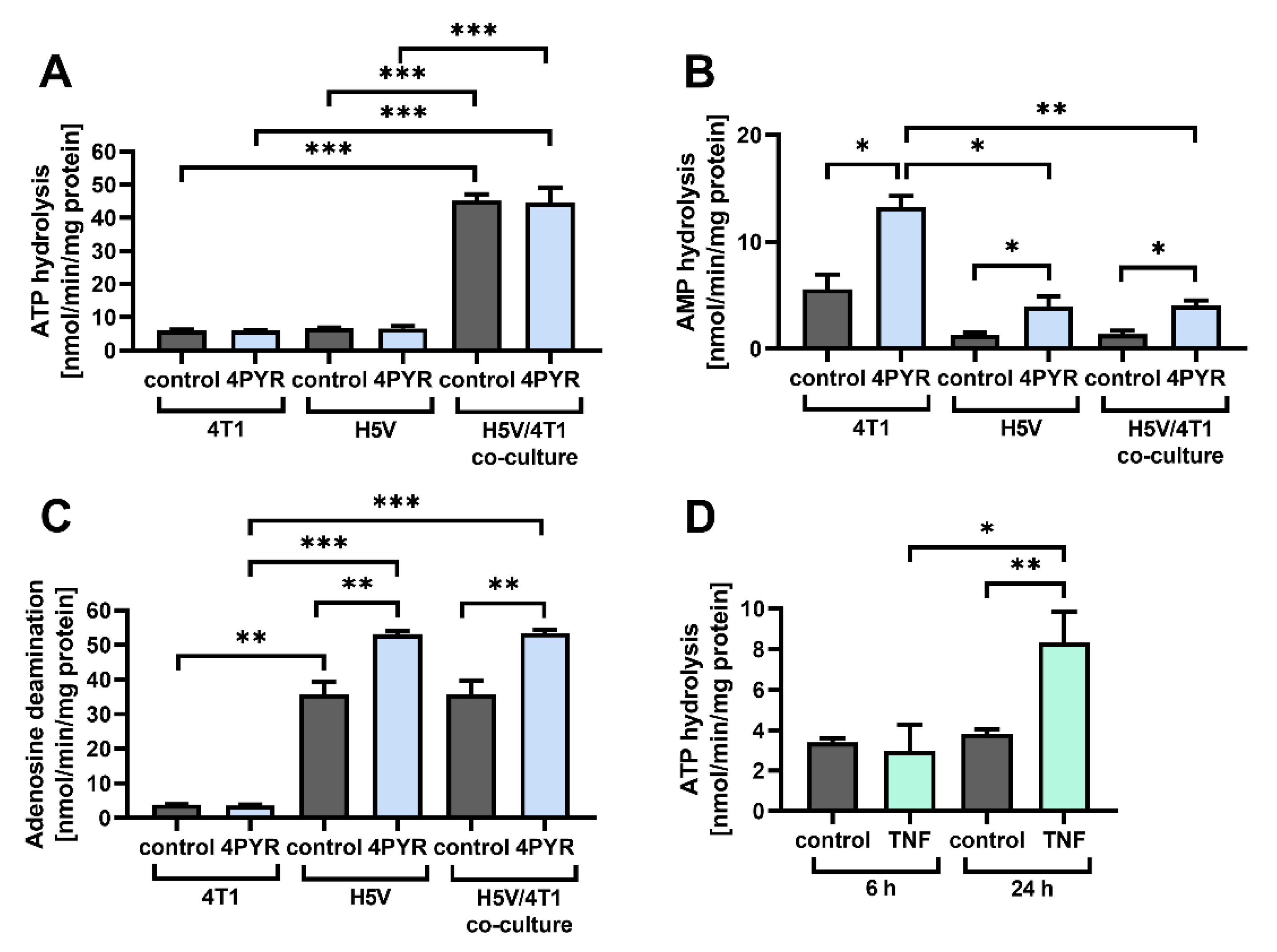
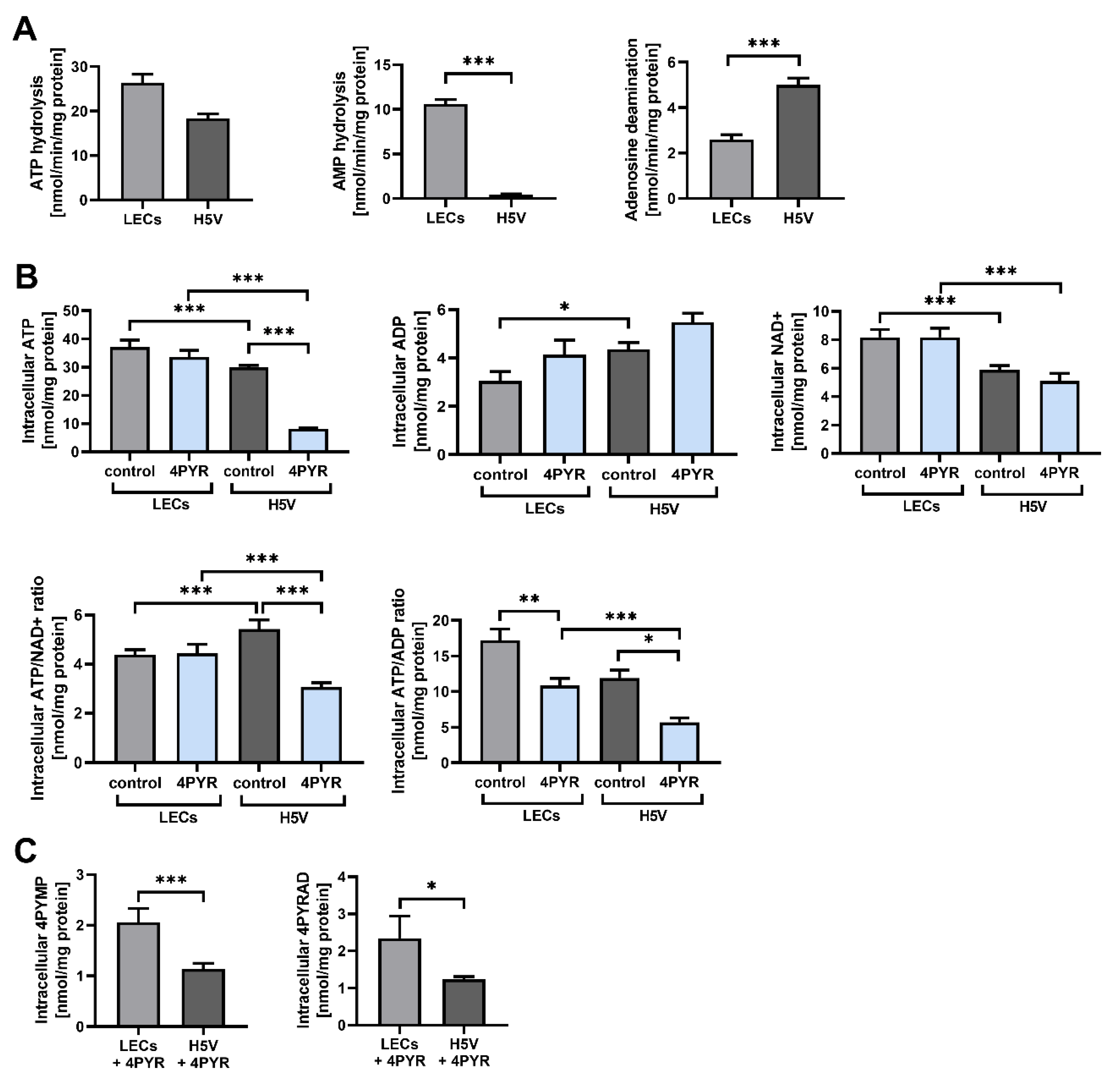
2.4. 4PYR Up-Regulates AMP Hydrolysis and Adenosine Deamination during Paracrine Interactions between Cancer and Endothelial Cells, Leading to a Significant Accumulation of Extracellular Inosine
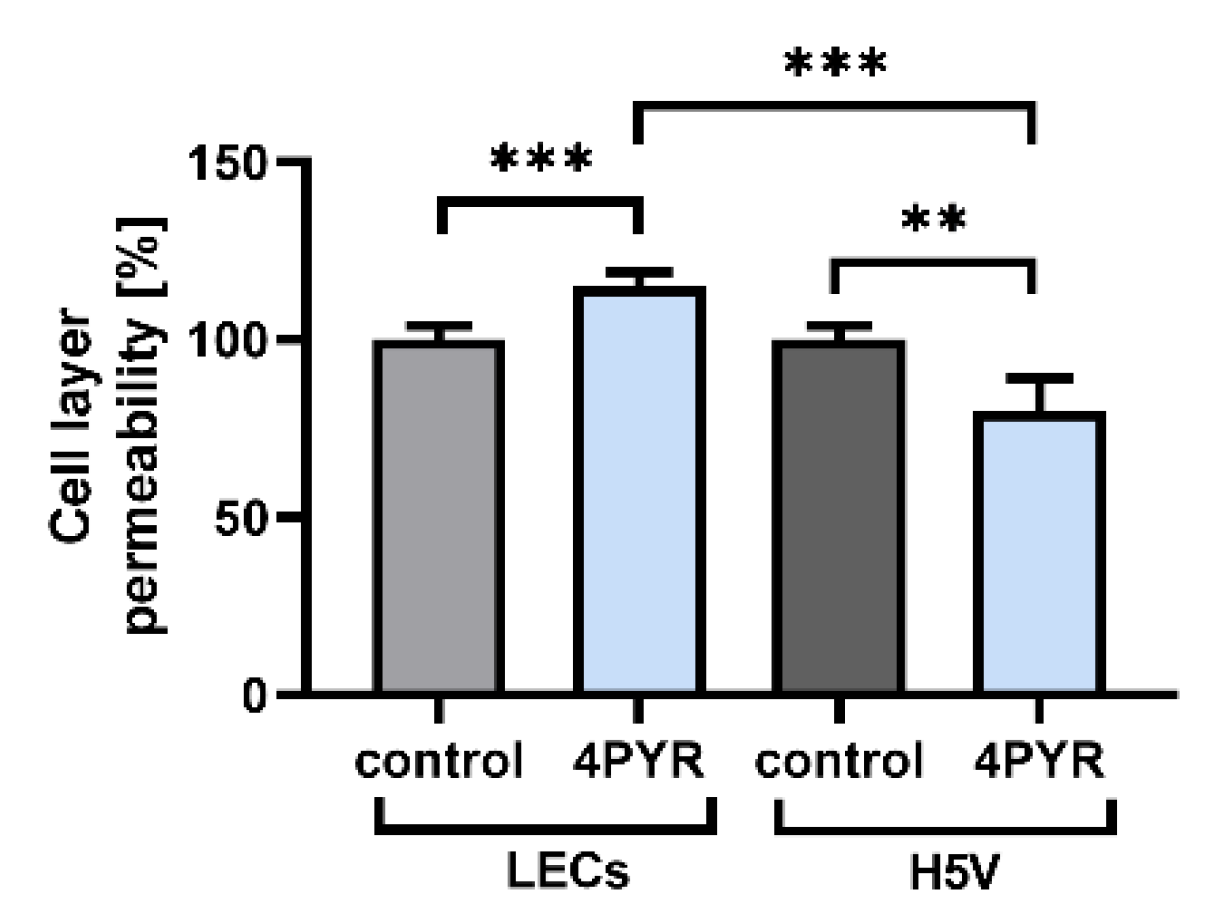
2.5. 4PYR Can Induce Contradictory Alterations in the Barrier Function of Endothelial Cells in Correlation with Differences in Their Extracellular Adenine Nucleotide Metabolism, Energy Status, and Oxidoreductive Potential
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture Conditions
4.3. Cell Viability Assay
4.4. Wound Healing Assay
4.5. Migration and Invasion Assays
4.6. Cell Adhesion Assay
4.7. Murine Model of 4T1 Breast Cancer
4.8. Isolation and Culture of Murine Lung Endothelial Cells
4.9. In Vitro Endothelial Permeability Assay
4.10. Tumor/Endothelial Cells Co-Culture
4.11. Determination of Intracellular Nucleotide and 4PYR Derivatives Levels and Extracellular Nucleotide Metabolism
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Slominska, E.M.; Carrey, E.A.; Foks, H.; Orlewska, C.; Wieczerzak, E.; Sowinski, P.; Yacoub, M.H.; Marinaki, A.M.; Simmonds, H.A.; Smolenski, R.T. A Novel Nucleotide Found in Human Erythrocytes, 4-Pyridone-3-Carboxamide-1-β-d-Ribonucleoside Triphosphate. J. Biol. Chem. 2006, 281, 32057–32064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mrochek, J.E.; Dinsmore, S.R.; Waalkes, T.P. Analytic Techniques in the Separation and Identification of Specific Purine and Pyrimidine Degradation Products of TRNA: Application to Urine Samples from Cancer Patients. J. Natl. Cancer Inst. 1974, 53, 1553–1563. [Google Scholar] [PubMed]
- Dutta, S.P.; Crain, P.F.; McCloskey, J.A.; Chheda, G.B. Isolation and Characterization of 1-β--Ribofuranosylpyridin-4-One-3-Carboxamide from Human Urine. Life Sci. 1979, 24, 1381–1388. [Google Scholar] [CrossRef]
- Speer, J.; Gehrke, C.W.; Kuo, K.C.; Waalkes, T.P.; Borek, E. TRNA Breakdown Products as Markers for Cancer. Cancer 1979, 44, 2120–2123. [Google Scholar] [CrossRef]
- Jablonska, P.; Mierzejewska, P.; Kutryb-Zajac, B.; Rzyman, W.; Dziadziuszko, R.; Polanska, J.; Sitkiewicz, M.; Smolenski, R.T.; Slominska, E.M. Increased Plasma Concentration of 4-Pyridone-3-Carboxamide-1-ß-D-Ribonucleoside (4PYR) in Lung Cancer. Preliminary Studies. Nucleosides Nucleotides Nucleic Acids 2019, 38, 781–787. [Google Scholar] [CrossRef]
- Synesiou, E.; Fairbanks, L.D.; Simmonds, H.A.; Slominska, E.M.; Smolenski, R.T.; Carrey, E.A. 4-Pyridone-3-Carboxamide-1-β-d-Ribonucleoside Triphosphate (4PyTP), a Novel NAD+ Metabolite Accumulating in Erythrocytes of Uremic Children: A Biomarker for a Toxic NAD+ Analogue in Other Tissues? Toxins 2011, 3, 520–537. [Google Scholar] [CrossRef] [Green Version]
- Pelikant-Malecka, I.; Sielicka, A.; Kaniewska, E.; Smolenski, R.T.; Slominska, E.M. Endothelial Toxicity of Unusual Nucleotide Metabolites. Pharmacol. Rep. 2015, 67, 818–822. [Google Scholar] [CrossRef]
- Kurosaki, M.; Demontis, S.; Barzago, M.M.; Garattini, E.; Terao, M. Molecular Cloning of the CDNA Coding for Mouse Aldehyde Oxidase: Tissue Distribution and Regulation in Vivo by Testosterone. Biochem. J. 1999, 341 Pt 1, 71–80. [Google Scholar] [CrossRef]
- Dalvie, D.; Di, L. Aldehyde Oxidase and Its Role as a Drug Metabolizing Enzyme. Pharmacol. Ther. 2019, 201, 137–180. [Google Scholar] [CrossRef]
- Stagg, J.; Divisekera, U.; Duret, H.; Sparwasser, T.; Teng, M.W.L.; Darcy, P.K.; Smyth, M.J. CD73-Deficient Mice Have Increased Antitumor Immunity and Are Resistant to Experimental Metastasis. Cancer Res. 2011, 71, 2892–2900. [Google Scholar] [CrossRef] [Green Version]
- Gao, Z.; Dong, K.; Zhang, H. The Roles of CD73 in Cancer. BioMed Res. Int. 2014, 2014, 460654. [Google Scholar] [CrossRef] [PubMed]
- Krüger, K.; Thompson, L.; Kaufmann, M.; Möller, P. Expression of Ecto-5′-Nucleotidase (CD73) in Normal Mammary Gland and in Breast Carcinoma. Br. J. Cancer 1991, 63, 114–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, H.; Zebisch, M.; Sträter, N. Cellular Function and Molecular Structure of Ecto-Nucleotidases. Purinergic Signal. 2012, 8, 437–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fishman, P.; Bar-Yehuda, S.; Synowitz, M.; Powell, J.D.; Klotz, K.N.; Gessi, S.; Borea, P.A. Adenosine Receptors and Cancer. In Adenosine Receptors in Health and Disease; Wilson, C.N., Mustafa, S.J., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 193, pp. 399–441. ISBN 978-3-540-89614-2. [Google Scholar]
- Garavaglia, S.; Bruzzone, S.; Cassani, C.; Canella, L.; Allegrone, G.; Sturla, L.; Mannino, E.; Millo, E.; De Flora, A.; Rizzi, M. The High-Resolution Crystal Structure of Periplasmic Haemophilus Influenzae NAD Nucleotidase Reveals a Novel Enzymatic Function of Human CD73 Related to NAD Metabolism. Biochem. J. 2012, 441, 131–141. [Google Scholar] [CrossRef]
- Grozio, A.; Sociali, G.; Sturla, L.; Caffa, I.; Soncini, D.; Salis, A.; Raffaelli, N.; De Flora, A.; Nencioni, A.; Bruzzone, S. CD73 Protein as a Source of Extracellular Precursors for Sustained NAD+ Biosynthesis in FK866-Treated Tumor Cells. J. Biol. Chem. 2013, 288, 25938–25949. [Google Scholar] [CrossRef] [Green Version]
- Slominska, E.M.; Orlewska, C.; Yuen, A.; Osman, L.; Romaszko, P.; Sokolowska, E.; Foks, H.; Simmonds, H.A.; Yacoub, M.H.; Smolenski, R.T. Metabolism of 4-Pyridone-3-Carboxamide-1-ß-D-Ribonucleoside Triphosphate and Its Nucleoside Precursor in the Erythrocytes. Nucleosides Nucleotides Nucleic Acids 2008, 27, 830–834. [Google Scholar] [CrossRef]
- Pelikant-Małecka, I.; Sielicka, A.; Kaniewska, E.; Smoleński, R.T.; Słomińska, E.M. Influence of 4-Pyridone-3-Carboxamide-1Β-D-Ribonucleoside (4PYR) on Activities of Extracellular Enzymes in Endothelial Human Cells. Nucleosides Nucleotides Nucleic Acids 2016, 35, 732–736. [Google Scholar] [CrossRef]
- Pelikant-Malecka, I.; Kaniewska-Bednarczuk, E.; Szrok, S.; Sielicka, A.; Sledzinski, M.; Orlewska, C.; Smolenski, R.T.; Slominska, E.M. Metabolic Pathway of 4-Pyridone-3-Carboxamide-1β-d-Ribonucleoside and Its Effects on Cellular Energetics. Int. J. Biochem. Cell Biol. 2017, 88, 31–43. [Google Scholar] [CrossRef]
- Scimeca, M.; Urbano, N.; Bonfiglio, R.; Duggento, A.; Toschi, N.; Schillaci, O.; Bonanno, E. Novel Insights into Breast Cancer Progression and Metastasis: A Multidisciplinary Opportunity to Transition from Biology to Clinical Oncology. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2019, 1872, 138–148. [Google Scholar] [CrossRef]
- Corrado, M.; Scorrano, L.; Campello, S. Changing Perspective on Oncometabolites: From Metabolic Signature of Cancer to Tumorigenic and Immunosuppressive Agents. Oncotarget 2016, 7, 46692–46706. [Google Scholar] [CrossRef] [Green Version]
- Collins, R.R.J.; Patel, K.; Putnam, W.C.; Kapur, P.; Rakheja, D. Oncometabolites: A New Paradigm for Oncology, Metabolism, and the Clinical Laboratory. Clin. Chem. 2017, 63, 1812–1820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaur, P.; Nagaraja, G.M.; Zheng, H.; Gizachew, D.; Galukande, M.; Krishnan, S.; Asea, A. A Mouse Model for Triple-Negative Breast Cancer Tumor-Initiating Cells (TNBC-TICs) Exhibits Similar Aggressive Phenotype to the Human Disease. BMC Cancer 2012, 12, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allard, B.; Turcotte, M.; Stagg, J. CD73-Generated Adenosine: Orchestrating the Tumor-Stroma Interplay to Promote Cancer Growth. J. Biomed. Biotechnol. 2012, 2012, 485156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse Models of Metastasis: Progress and Prospects. Dis. Models Mech. 2017, 10, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schram, K.H. Urinary Nucleosides. Mass Spectrom. Rev. 1998, 17, 131–251. [Google Scholar] [CrossRef]
- Al-Gayyar, M.M.H.; Bagalagel, A.; Noor, A.O.; Almasri, D.M.; Diri, R. The Therapeutic Effects of Nicotinamide in Hepatocellular Carcinoma through Blocking IGF-1 and Effecting the Balance between Nrf2 and PKB. Biomed. Pharmacother. 2019, 112, 108653. [Google Scholar] [CrossRef] [PubMed]
- Honn, K.V.; Tang, D.G. Adhesion Molecules and Tumor Cell Interaction with Endothelium and Subendothelial Matrix. Cancer Metast. Rev. 1992, 11, 353–375. [Google Scholar] [CrossRef]
- Garlanda, C.; Parravicini, C.; Sironi, M.; De Rossi, M.; Wainstok de Calmanovici, R.; Carozzi, F.; Bussolino, F.; Colotta, F.; Mantovani, A.; Vecchi, A. Progressive Growth in Immunodeficient Mice and Host Cell Recruitment by Mouse Endothelial Cells Transformed by Polyoma Middle-Sized T Antigen: Implications for the Pathogenesis of Opportunistic Vascular Tumors. Proc. Natl. Acad. Sci. USA 1994, 91, 7291–7295. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Z.; Li, X.; Kang, N.; Yang, Y.; Chen, C.; Wu, T.; Zhao, M.; Liu, Y.; Ji, X. A Novel Specific Anti-CD73 Antibody Inhibits Triple-Negative Breast Cancer Cell Motility by Regulating Autophagy. Int. J. Mol. Sci. 2019, 20, 1057. [Google Scholar] [CrossRef] [Green Version]
- Koszałka, P.; Gołuńska, M.; Stanisławowski, M.; Urban, A.; Stasiłojć, G.; Majewski, M.; Wierzbicki, P.; Składanowski, A.C.; Bigda, J. CD73 on B16F10 Melanoma Cells in CD73-Deficient Mice Promotes Tumor Growth, Angiogenesis, Neovascularization, Macrophage Infiltration and Metastasis. Int. J. Biochem. Cell Biol. 2015, 69, 1–10. [Google Scholar] [CrossRef]
- Alefishat, E.; Alexander, S.P.H.; Ralevic, V. Effects of NAD at Purine Receptors in Isolated Blood Vessels. Purinergic Signal. 2015, 11, 47–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutryb-Zajac, B.; Koszalka, P.; Mierzejewska, P.; Bulinska, A.; Zabielska, M.A.; Brodzik, K.; Skrzypkowska, A.; Zelazek, L.; Pelikant-Malecka, I.; Slominska, E.M.; et al. Adenosine Deaminase Inhibition Suppresses Progression of 4T1 Murine Breast Cancer by Adenosine Receptor-Dependent Mechanisms. J. Cell Mol. Med. 2018, 22, 5939–5954. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. Extracellular ATP and Other Nucleotides—Ubiquitous Triggers of Intercellular Messenger Release. Purinergic Signal. 2016, 12, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Vandamme, W.; Braet, K.; Cabooter, L.; Leybaert, L. Tumour Necrosis Factor Alpha Inhibits Purinergic Calcium Signalling in Blood-Brain Barrier Endothelial Cells: TNF-α and Endothelial Calcium Signalling. J. Neurochem. 2003, 88, 411–421. [Google Scholar] [CrossRef]
- Corriden, R.; Insel, P.A. Basal Release of ATP: An Autocrine-Paracrine Mechanism for Cell Regulation. Sci. Signal. 2010, 3, re1. [Google Scholar] [CrossRef] [Green Version]
- Haskó, G.; Sitkovsky, M.V.; Szabó, C. Immunomodulatory and Neuroprotective Effects of Inosine. Trends Pharmacol. Sci. 2004, 25, 152–157. [Google Scholar] [CrossRef]
- Mierke, C.T.; Zitterbart, D.P.; Kollmannsberger, P.; Raupach, C.; Schlötzer-Schrehardt, U.; Goecke, T.W.; Behrens, J.; Fabry, B. Breakdown of the Endothelial Barrier Function in Tumor Cell Transmigration. Biophys. J. 2008, 94, 2832–2846. [Google Scholar] [CrossRef] [Green Version]
- Eltzschig, H.K.; Weissmüller, T.; Mager, A.; Eckle, T. Nucleotide Metabolism and Cell-Cell Interactions. In Cell-Cell Interactions in Health and Disease; Humana Press: Totowa, NJ, USA, 2006; Volume 341, pp. 73–88. ISBN 978-1-59745-113-0. [Google Scholar]
- Jambusaria, A.; Hong, Z.; Zhang, L.; Srivastava, S.; Jana, A.; Toth, P.T.; Dai, Y.; Malik, A.B.; Rehman, J. Endothelial Heterogeneity across Distinct Vascular Beds during Homeostasis and Inflammation. eLife 2020, 9, e51413. [Google Scholar] [CrossRef]
- Gao, Y.; Bado, I.; Wang, H.; Zhang, W.; Rosen, J.M.; Zhang, X.H.-F. Metastasis Organotropism: Redefining the Congenial Soil. Dev. Cell 2019, 49, 375–391. [Google Scholar] [CrossRef]
- Hiratsuka, S.; Goel, S.; Kamoun, W.S.; Maru, Y.; Fukumura, D.; Duda, D.G.; Jain, R.K. Endothelial Focal Adhesion Kinase Mediates Cancer Cell Homing to Discrete Regions of the Lungs via E-Selectin up-Regulation. Proc. Natl. Acad. Sci. USA 2011, 108, 3725–3730. [Google Scholar] [CrossRef] [Green Version]
- Noll, T.; Muhs, A.; Besselmann, M.; Watanabe, H.; Piper, H.M. Initiation of Hyperpermeability in Energy-Depleted Coronary Endothelial Monolayers. Am. J. Physiol.-Heart Circ. Physiol. 1995, 268, H1462–H1470. [Google Scholar] [CrossRef] [PubMed]
- Friedl, J.; Puhlmann, M.; Bartlett, D.L.; Libutti, S.K.; Turner, E.N.; Gnant, M.F.X.; Alexander, H.R. Induction of Permeability across Endothelial Cell Monolayers by Tumor Necrosis Factor (TNF) Occurs via a Tissue Factor-Dependent Mechanism: Relationship between the Procoagulant and Permeability Effects of TNF. Blood 2002, 100, 1334–1339. [Google Scholar] [PubMed] [Green Version]
- Seidel, A.; Brunner, S.; Seidel, P.; Fritz, G.I.; Herbarth, O. Modified Nucleosides: An Accurate Tumour Marker for Clinical Diagnosis of Cancer, Early Detection and Therapy Control. Br. J. Cancer 2006, 94, 1726–1733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Auchampach, J.A. Adenosine Receptors and Angiogenesis. Circ. Res. 2007, 101, 1075–1077. [Google Scholar] [CrossRef] [PubMed]
- Schulte, G.; Fredholm, B.B. Signalling from Adenosine Receptors to Mitogen-Activated Protein Kinases. Cell. Signal. 2003, 15, 813–827. [Google Scholar] [CrossRef]
- Koszałka, P.; Pryszlak, A.; Gołuńska, M.; Kolasa, J.; Stasiłojć, G.; Składanowski, A.C.; Bigda, J.J. Inhibition of CD73 Stimulates the Migration and Invasion of B16F10 Melanoma Cells in Vitro, but Results in Impaired Angiogenesis and Reduced Melanoma Growth in Vivo. Oncol. Rep. 2014, 31, 819–827. [Google Scholar] [CrossRef] [Green Version]
- Gregório, A.C.; Fonseca, N.A.; Moura, V.; Lacerda, M.; Figueiredo, P.; Simões, S.; Dias, S.; Moreira, J.N. Inoculated Cell Density as a Determinant Factor of the Growth Dynamics and Metastatic Efficiency of a Breast Cancer Murine Model. PLoS ONE 2016, 11, e0165817. [Google Scholar] [CrossRef]
- Eales, K.L.; Hollinshead, K.E.R.; Tennant, D.A. Hypoxia and Metabolic Adaptation of Cancer Cells. Oncogenesis 2016, 5, e190. [Google Scholar] [CrossRef] [Green Version]
- Fredholm, B.B. Adenosine—A Physiological or Pathophysiological Agent? J. Mol. Med. 2014, 92, 201–206. [Google Scholar] [CrossRef]
- Kalsi, K.; Lawson, C.; Dominguez, M.; Taylor, P.; Yacoub, M.H.; Smolenski, R.T. Regulation of Ecto-5′-Nucleotidase by TNF-α in Human Endothelial Cells. Mol. Cell. Biochem. 2002, 232, 113–119. [Google Scholar] [CrossRef]
- Di Virgilio, F.; Adinolfi, E. Extracellular Purines, Purinergic Receptors and Tumor Growth. Oncogene 2017, 36, 293–303. [Google Scholar] [CrossRef] [PubMed]
- Fredholm, B.B.; Irenius, E.; Kull, B.; Schulte, G. Comparison of the Potency of Adenosine as an Agonist at Human Adenosine Receptors Expressed in Chinese Hamster Ovary Cells11Abbreviations: CAMP, Cyclic Adenosine 3′,5′-Monophosphate; CHO, Chinese Hamster Ovary; NBMPR, Nitrobenzylthioinosine; and NECA, 5′-N-Ethyl Carboxamido Adenosine. Biochem. Pharmacol. 2001, 61, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Stagg, J.; Divisekera, U.; McLaughlin, N.; Sharkey, J.; Pommey, S.; Denoyer, D.; Dwyer, K.M.; Smyth, M.J. Anti-CD73 Antibody Therapy Inhibits Breast Tumor Growth and Metastasis. Proc. Natl. Acad. Sci. USA 2010, 107, 1547–1552. [Google Scholar] [CrossRef] [Green Version]
- Ireson, C.R.; Alavijeh, M.S.; Palmer, A.M.; Fowler, E.R.; Jones, H.J. The Role of Mouse Tumour Models in the Discovery and Development of Anticancer Drugs. Br. J. Cancer 2019, 121, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Azevedo, A.S.; Follain, G.; Patthabhiraman, S.; Harlepp, S.; Goetz, J.G. Metastasis of Circulating Tumor Cells: Favorable Soil or Suitable Biomechanics, or Both? Cell Adhes. Migr. 2015, 9, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Labuschagne, C.F.; Cheung, E.C.; Blagih, J.; Domart, M.-C.; Vousden, K.H. Cell Clustering Promotes a Metabolic Switch That Supports Metastatic Colonization. Cell Metab. 2019, 30, 720–734.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slominska, E.M.; Borkowski, T.; Rybakowska, I.; Abramowicz-Glinka, M.; Orlewska, C.; Smolenski, R.T. In Vitro and Cellular Effects of 4-Pyridone-3-Carboxamide Riboside on Enzymes of Nucleotide Metabolism. Nucleosides Nucleotides Nucleic Acids 2014, 33, 353–357. [Google Scholar] [CrossRef]
- Zabielska, M.A.; Borkowski, T.; Slominska, E.M.; Smolenski, R.T. Inhibition of AMP Deaminase as Therapeutic Target in Cardiovascular Pathology. Pharmacol. Rep. 2015, 67, 682–688. [Google Scholar] [CrossRef]
- Tang, Z.; Ye, W.; Chen, H.; Kuang, X.; Guo, J.; Xiang, M.; Peng, C.; Chen, X.; Liu, H. Role of Purines in Regulation of Metabolic Reprogramming. Purinergic Signal. 2019, 15, 423–438. [Google Scholar] [CrossRef]
- Eyles, J.; Puaux, A.-L.; Wang, X.; Toh, B.; Prakash, C.; Hong, M.; Tan, T.G.; Zheng, L.; Ong, L.C.; Jin, Y.; et al. Tumor Cells Disseminate Early, but Immunosurveillance Limits Metastatic Outgrowth, in a Mouse Model of Melanoma. J. Clin. Investig. 2010, 120, 2030–2039. [Google Scholar] [CrossRef]
- Massagué, J.; Obenauf, A.C. Metastatic Colonization by Circulating Tumour Cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef] [PubMed]
- Furlow, P.W.; Zhang, S.; Soong, T.D.; Halberg, N.; Goodarzi, H.; Mangrum, C.; Wu, Y.G.; Elemento, O.; Tavazoie, S.F. Mechanosensitive Pannexin-1 Channels Mediate Microvascular Metastatic Cell Survival. Nat. Cell Biol. 2015, 17, 943–952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allard, D.; Chrobak, P.; Allard, B.; Messaoudi, N.; Stagg, J. Targeting the CD73-Adenosine Axis in Immuno-Oncology. Immunol. Lett. 2019, 205, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Haskó, G.; Kuhel, D.G.; Németh, Z.H.; Mabley, J.G.; Stachlewitz, R.F.; Virág, L.; Lohinai, Z.; Southan, G.J.; Salzman, A.L.; Szabó, C. Inosine Inhibits Inflammatory Cytokine Production by a Posttranscriptional Mechanism and Protects Against Endotoxin-Induced Shock. J. Immunol. 2000, 164, 1013–1019. [Google Scholar] [CrossRef]
- Junqueira, S.C.; dos Santos Coelho, I.; Lieberknecht, V.; Cunha, M.P.; Calixto, J.B.; Rodrigues, A.L.S.; Santos, A.R.S.; Dutra, R.C. Inosine, an Endogenous Purine Nucleoside, Suppresses Immune Responses and Protects Mice from Experimental Autoimmune Encephalomyelitis: A Role for A2A Adenosine Receptor. Mol. Neurobiol. 2017, 54, 3271–3285. [Google Scholar] [CrossRef]
- Thompson, L.F.; Eltzschig, H.K.; Ibla, J.C.; Van De Wiele, C.J.; Resta, R.; Morote-Garcia, J.C.; Colgan, S.P. Crucial Role for Ecto-5′-Nucleotidase (CD73) in Vascular Leakage during Hypoxia. J. Exp. Med. 2004, 200, 1395–1405. [Google Scholar] [CrossRef]
- Bowser, J.L.; Blackburn, M.R.; Shipley, G.L.; Molina, J.G.; Dunner, K.; Broaddus, R.R. Loss of CD73-Mediated Actin Polymerization Promotes Endometrial Tumor Progression. J. Clin. Investig. 2015, 126, 220–238. [Google Scholar] [CrossRef] [Green Version]
- Bowser, J.L.; Broaddus, R.R. CD73s Protection of Epithelial Integrity: Thinking beyond the Barrier. Tissue Barriers 2016, 4, e1224963. [Google Scholar] [CrossRef] [Green Version]
- Mierzejewska, P.; Kunc, M.; Zabielska-Kaczorowska, M.A.; Kutryb-Zajac, B.; Pelikant-Malecka, I.; Braczko, A.; Jablonska, P.; Romaszko, P.; Koszalka, P.; Szade, J.; et al. An Unusual Nicotinamide Derivative, 4-Pyridone-3-Carboxamide Ribonucleoside (4PYR), Is a Novel Endothelial Toxin and Oncometabolite. Exp. Mol. Med. 2021, 53, 1402–1412. [Google Scholar] [CrossRef]
- Kutryb-Zajac, B.; Yuen, A.H.Y.; Khalpey, Z.; Zukowska, P.; Slominska, E.M.; Taylor, P.M.; Goldstein, S.; Heacox, A.E.; Lavitrano, M.; Chester, A.H.; et al. Nucleotide Catabolism on the Surface of Aortic Valve Xenografts; Effects of Different Decellularization Strategies. J. Cardiovasc. Trans. Res. 2016, 9, 119–126. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koszalka, P.; Kutryb-Zajac, B.; Mierzejewska, P.; Tomczyk, M.; Wietrzyk, J.; Serafin, P.K.; Smolenski, R.T.; Slominska, E.M. 4-Pyridone-3-carboxamide-1-β-D-ribonucleoside (4PYR)—A Novel Oncometabolite Modulating Cancer-Endothelial Interactions in Breast Cancer Metastasis. Int. J. Mol. Sci. 2022, 23, 5774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105774
Koszalka P, Kutryb-Zajac B, Mierzejewska P, Tomczyk M, Wietrzyk J, Serafin PK, Smolenski RT, Slominska EM. 4-Pyridone-3-carboxamide-1-β-D-ribonucleoside (4PYR)—A Novel Oncometabolite Modulating Cancer-Endothelial Interactions in Breast Cancer Metastasis. International Journal of Molecular Sciences. 2022; 23(10):5774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105774
Chicago/Turabian StyleKoszalka, Patrycja, Barbara Kutryb-Zajac, Paulina Mierzejewska, Marta Tomczyk, Joanna Wietrzyk, Pawel K. Serafin, Ryszard T. Smolenski, and Ewa M. Slominska. 2022. "4-Pyridone-3-carboxamide-1-β-D-ribonucleoside (4PYR)—A Novel Oncometabolite Modulating Cancer-Endothelial Interactions in Breast Cancer Metastasis" International Journal of Molecular Sciences 23, no. 10: 5774. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105774