
Nimodipine Treatment Protects Auditory Hair Cells from Cisplatin-Induced Cell Death Accompanied by Upregulation of LMO4


Abstract
:1. Introduction
2. Results
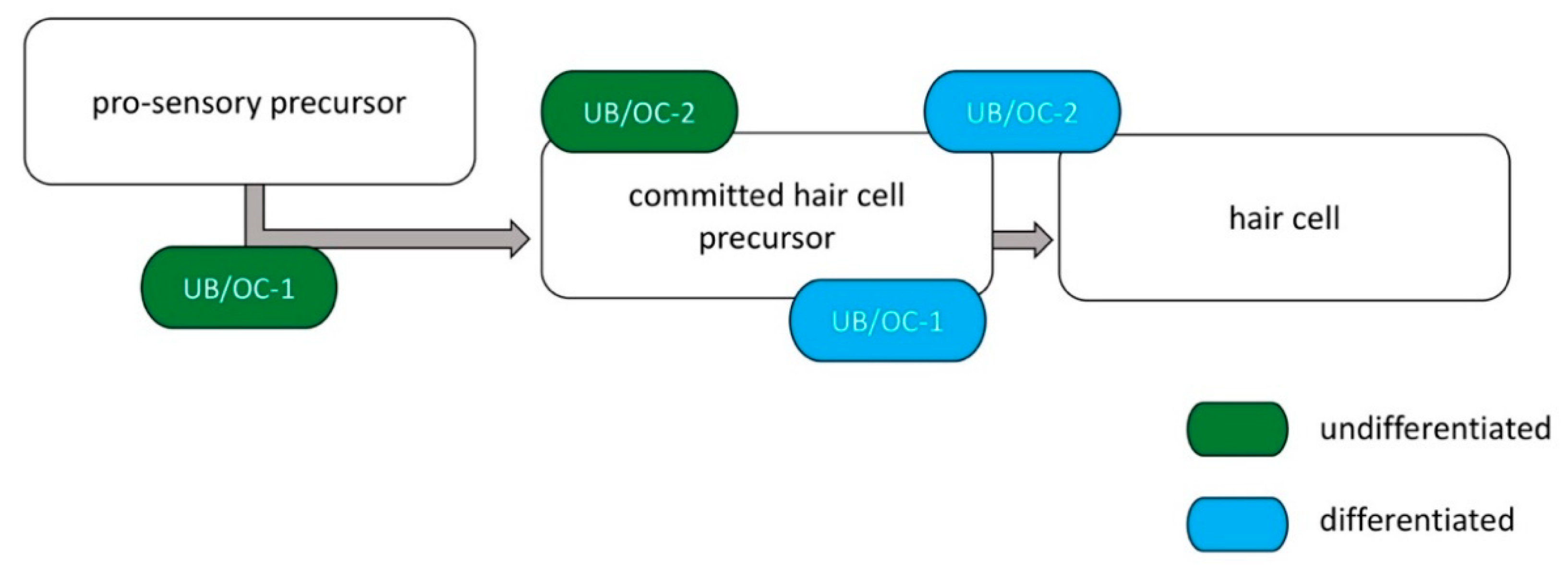
2.1. Increase in Specific Hair Cell Markers after Differentiation of UB/OC−1 and UB/OC−2
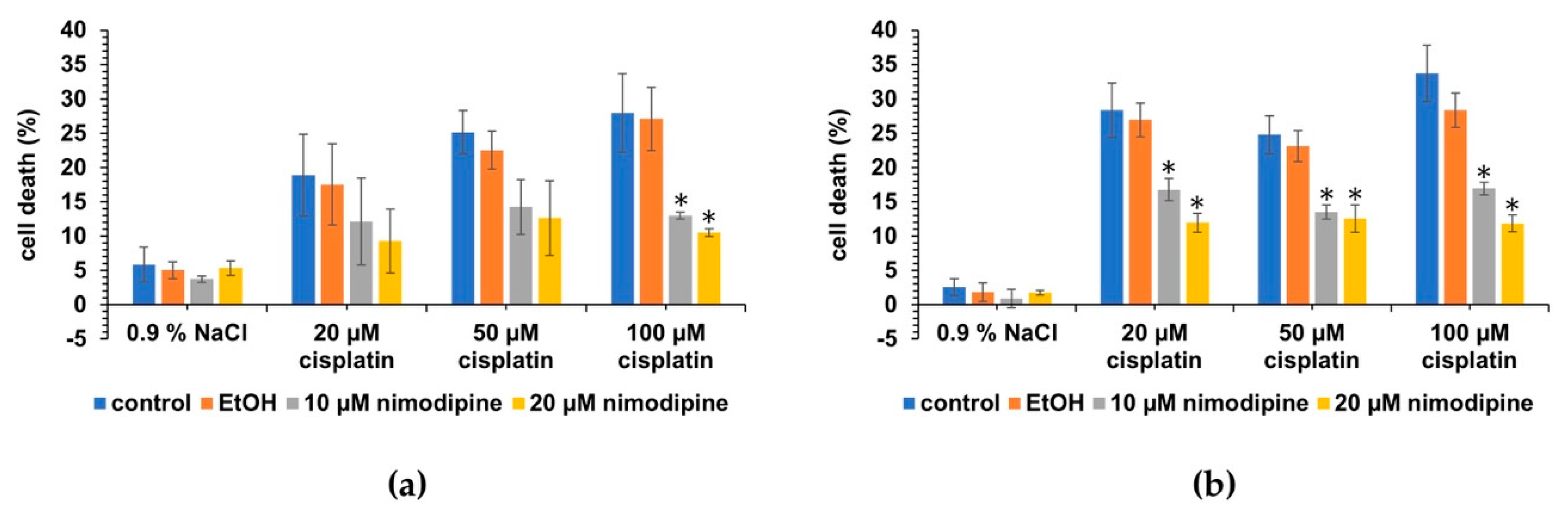
2.2. Nimodipine Decreases Cisplatin-Induced Cytotoxicity in Undifferentiated Hair Cells
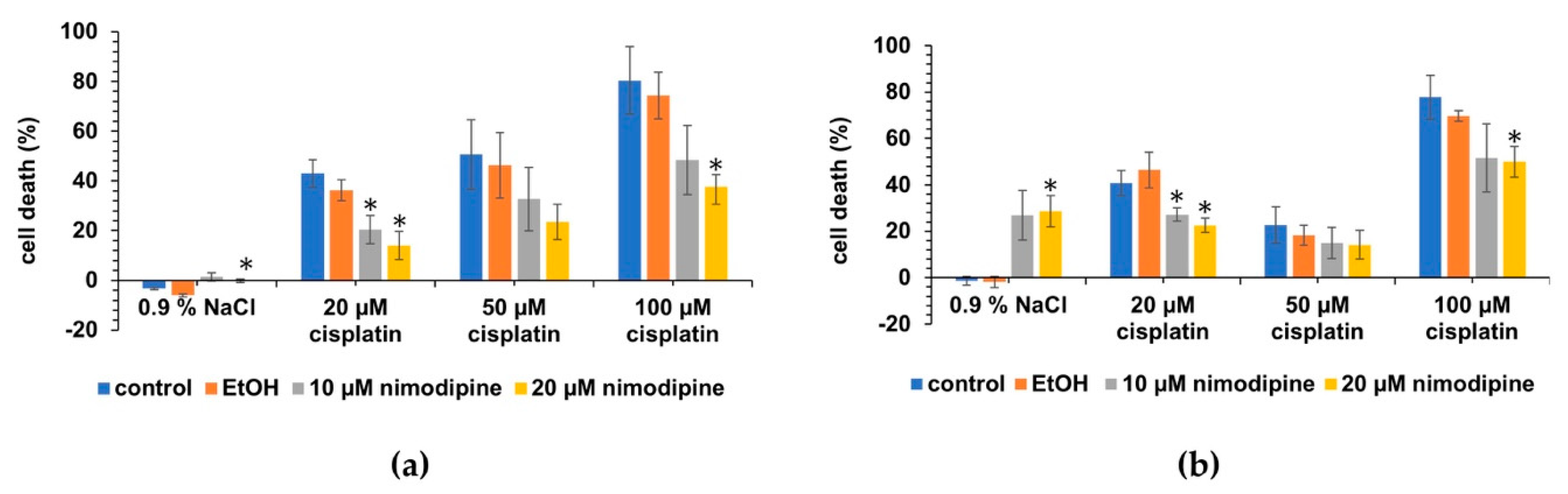
2.3. Nimodipine Decreases Cisplatin-Induced Cytotoxicity in Differentiated Hair Cells
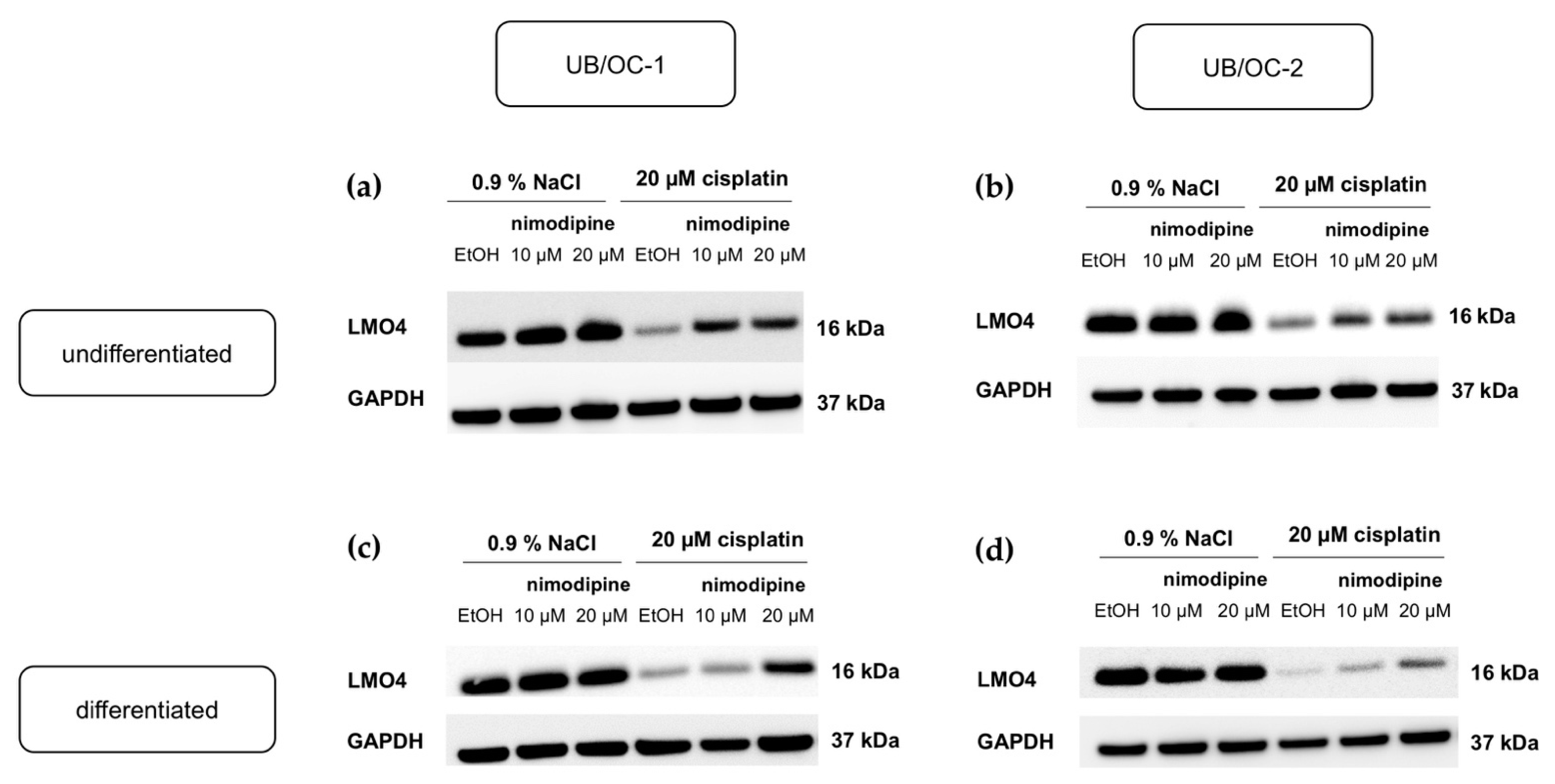
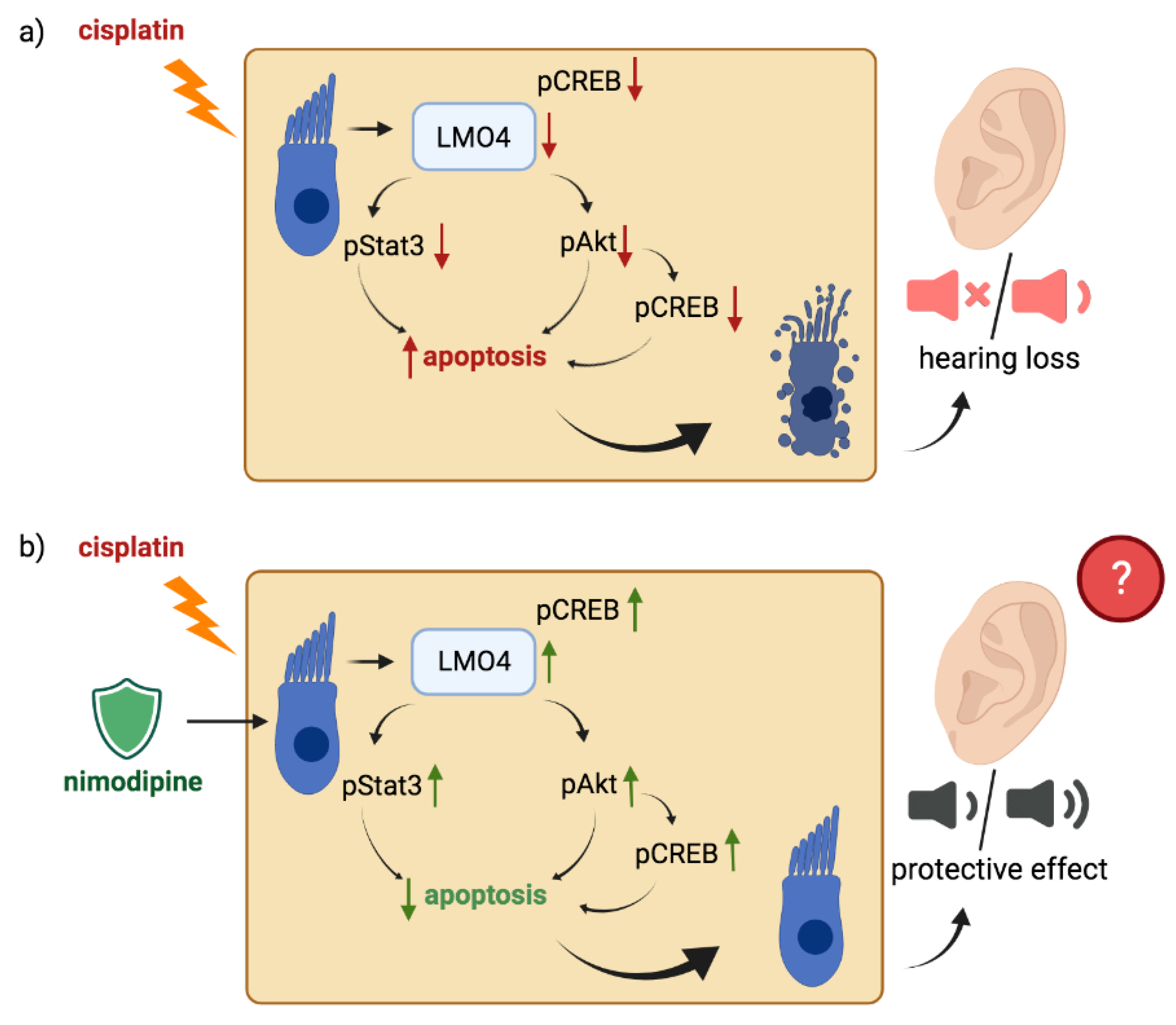
2.4. Nimodipine Counteracts the Downregulation of LMO4 by Cisplatin in Undifferentiated and Differentiated State of Hair Cells
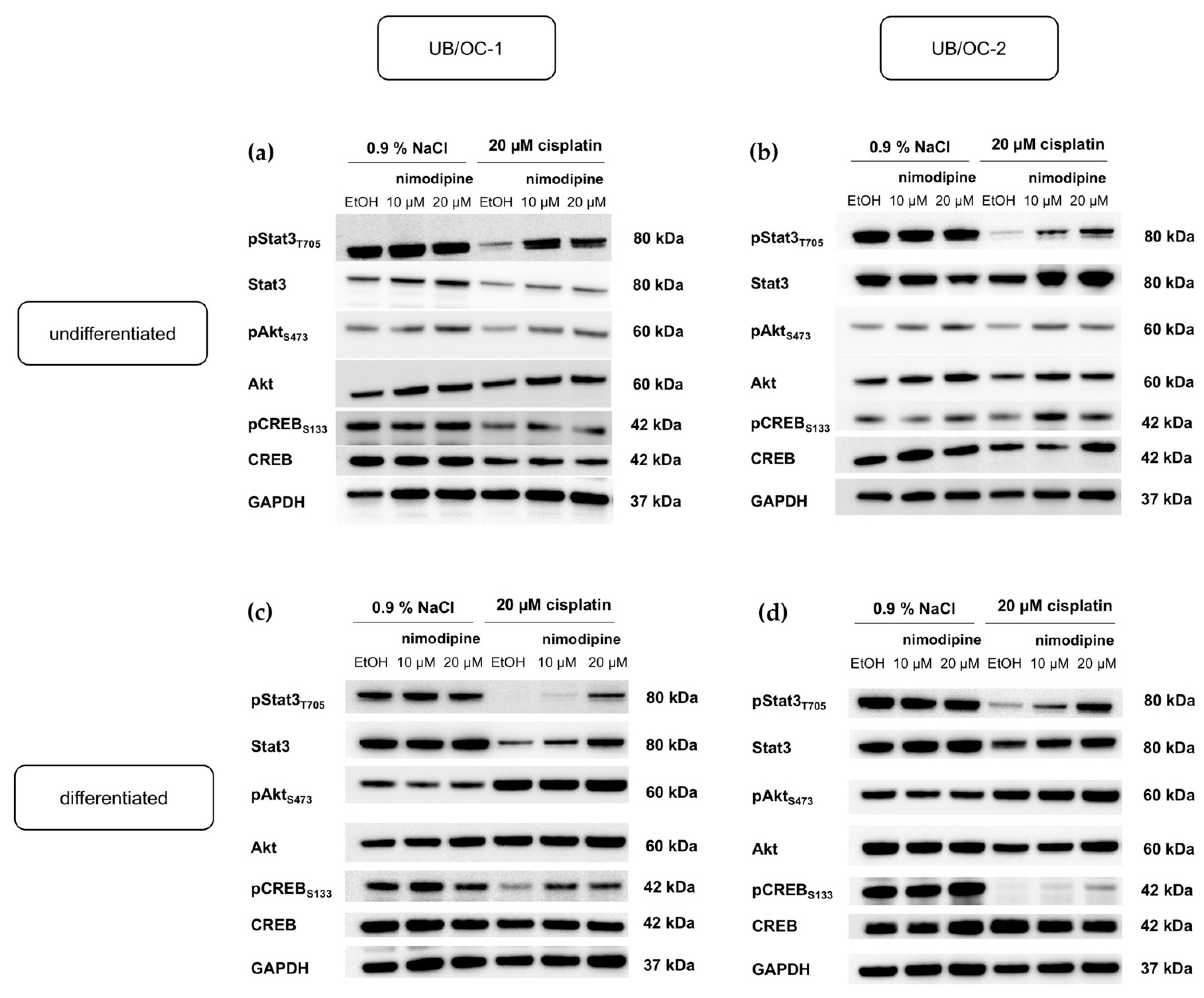
2.5. Activation of Anti-Apoptotic Pathways by Nimodipine under Chemotherapy with Cisplatin in Undifferentiated and Differentiated State of Hair Cells
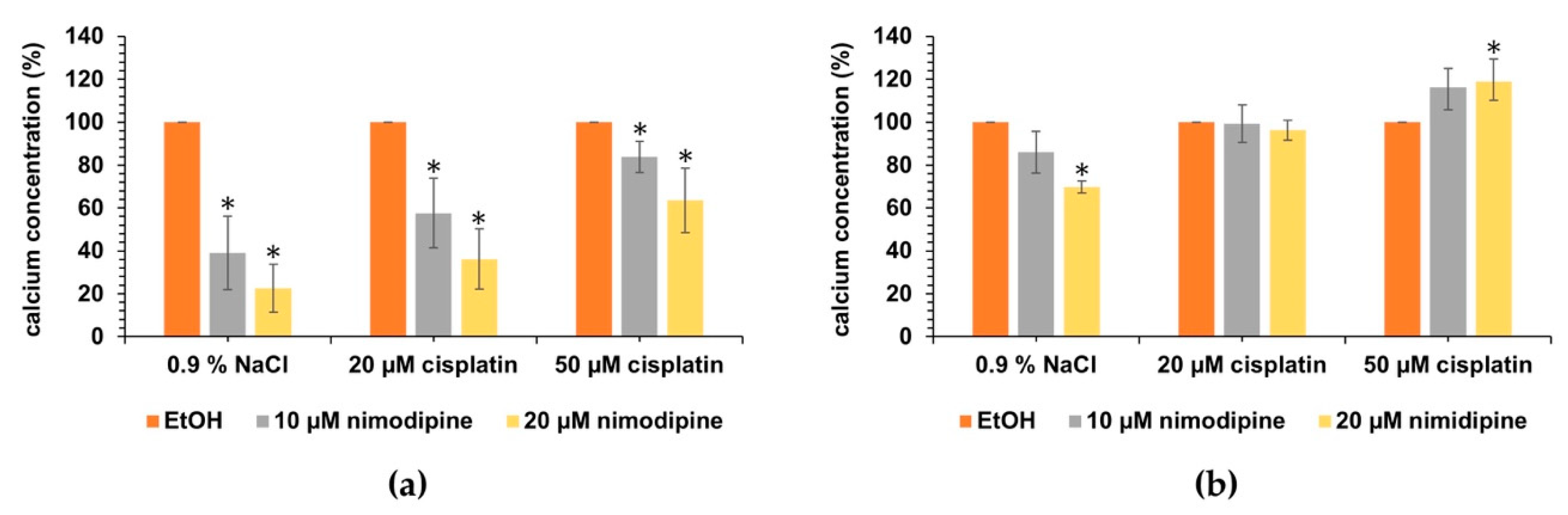
2.6. Influence of Nimodipine on the Intracellular Calcium Concentration of Auditory Hair Cells under Cisplatin Treatment
3. Discussion
4. Materials and Methods
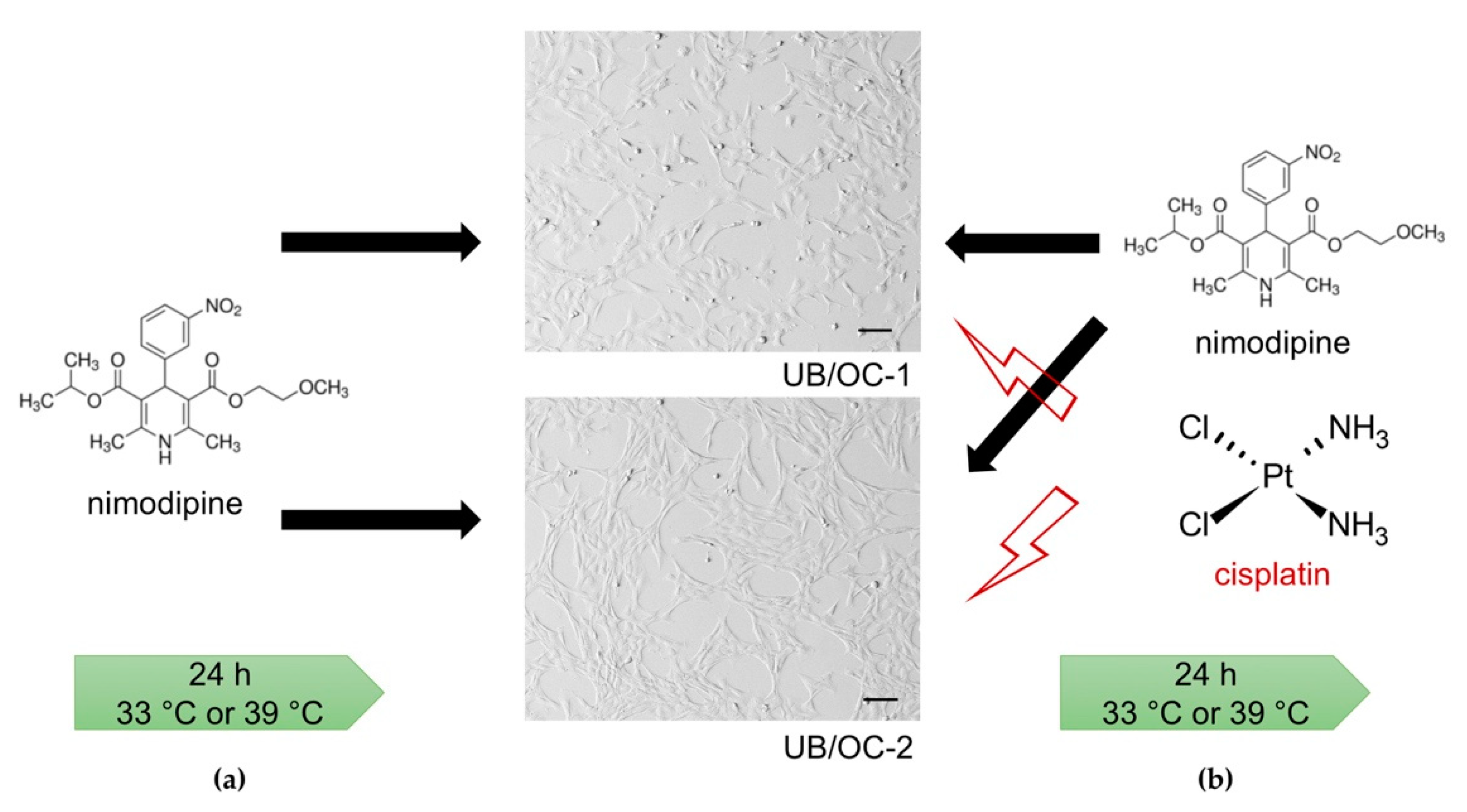
4.1. Cell Lines
4.2. Cell Treatment
4.3. Cytotoxicity Measurement
4.4. Western Blot
4.5. Intracellular Calcium Measurement
4.6. RNA Isolation and Real-Time Quantitative PCR
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carlson, A.P.; Hänggi, D.; Macdonald, R.L.; Shuttleworth, C.W. Nimodipine reappraised: An old drug with a future. Curr. Neuropharmacol. 2019, 18, 65–82. [Google Scholar] [CrossRef] [PubMed]
- Monzani, D.; Genovese, E.; Pini, L.A.; Di Berardino, F.; Alicandri Ciufelli, M.; Galeazzi, G.M.; Presutti, L. Nimodipine in otolaryngology: From past evidence to clinical perspectives. Acta Otorhinolaryngol. Ital. 2015, 35, 135–145. [Google Scholar] [PubMed]
- Mahmoud, S.H.; Ji, X.; Isse, F.A. Nimodipine Pharmacokinetic Variability in Various Patient Populations. Drugs R D 2020, 20, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Scheller, C.; Wienke, A.; Tatagiba, M.; Gharabaghi, A.; Ramina, K.F.; Ganslandt, O.; Bischoff, B.; Zenk, J.; Engelhorn, T.; Matthies, C.; et al. Prophylactic nimodipine treatment for cochlear and facial nerve preservation after vestibular schwannoma surgery: A randomized multicenter phase III trial. J. Neurosurg. 2016, 124, 657–664. [Google Scholar] [CrossRef] [Green Version]
- Scheller, C.; Rampp, S.; Leisz, S.; Tatagiba, M.; Gharabaghi, A.; Ramina, K.F.; Ganslandt, O.; Matthies, C.; Westermaier, T.; Antoniadis, G.; et al. Prophylactic nimodipine treatment improves hearing outcome after vestibular schwannoma surgery in men: A subgroup analysis of a randomized multicenter phase III trial. Neurosurg. Rev. 2020, 44, 1729–1735. [Google Scholar] [CrossRef]
- Leisz, S.; Simmermacher, S.; Prell, J.; Strauss, C.; Scheller, C. Nimodipine-dependent protection of schwann cells, astrocytes and neuronal cells from osmotic, oxidative and heat stress is associated with the activation of AKT and CREB. Int. J. Mol. Sci. 2019, 20, 4578. [Google Scholar] [CrossRef] [Green Version]
- Scheller, C.; Herzfeld, E.; Strauss, C. Neuroprotective medication in vestibular schwannoma surgery. HNO 2017, 65, 714–718. [Google Scholar] [CrossRef]
- Scheller, C.; Wienke, A.; Tatagiba, M.; Gharabaghi, A.; Ramina, K.F.; Ganslandt, O.; Bischoff, B.; Zenk, J.; Engelhorn, T.; Matthies, C.; et al. Prophylactic nimodipine treatment and improvement in hearing outcome after vestibular schwannoma surgery: A combined analysis of a randomized, multicenter, phase III trial and its pilot study. J. Neurosurg. 2017, 127, 1376–1383. [Google Scholar] [CrossRef] [Green Version]
- Bork, K.; Wurm, F.; Haller, H.; Strauss, C.; Scheller, C.; Gnanapragassam, V.S.; Horstkorte, R. Neuroprotective and neuroregenerative effects of nimodipine in a model system of neuronal differentiation and neurite outgrowth. Molecules 2015, 20, 1003–1013. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.K.; Lee, G.-J.; Choi, S.; Kim, Y.J.; Park, H.-K.; Park, B.J. Neuroprotective effects by nimodipine treatment in the experimental global ischemic rat model: Real time estimation of glutamate. J. Korean Neurosurg. Soc. 2011, 49, 1–7. [Google Scholar] [CrossRef]
- Tang, Y.-D.; Zheng, X.-S.; Ying, T.-T.; Yuan, Y.; Li, S.-T. Nimodipine-mediated re-myelination after facial nerve crush injury in rats. J. Clin. Neurosci. 2015, 22, 1661–1668. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Wang, J.; Li, Q.-Y.; Yu, J.-Z.; Ma, C.-G.; Wang, X.; Lu, C.-Z.; Xiao, B.-G. Neuroprotection and CD131/GDNF/AKT pathway of carbamylated erythropoietin in hypoxic neurons. Mol. Neurobiol. 2017, 54, 5051–5060. [Google Scholar] [CrossRef] [PubMed]
- Walton, M.R.; Dragunow, I.M. Is CREB a key to neuronal survival? Trends Neurosci. 2000, 23, 48–53. [Google Scholar] [CrossRef]
- Xu, Y.-Q.; Long, L.; Yan, J.-Q.; Wei, L.; Pan, M.-Q.; Gao, H.-M.; Zhou, P.; Liu, M.; Zhu, C.-S.; Tang, B.-S.; et al. Simvastatin induces neuroprotection in 6-OHDA-lesioned PC12 via the PI3K/AKT/caspase 3 pathway and anti-inflammatory responses. CNS Neurosci. Ther. 2013, 19, 170–177. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, R.; Tong, Y.; Chen, P.; Shen, Y.; Miao, S.; Liu, X. Neuroprotection by dihydrotestosterone in LPS-induced neuroinflammation. Neurobiol. Dis. 2020, 140, 104814. [Google Scholar] [CrossRef]
- Sangaran, P.G.; Ibrahim, Z.A.; Chik, Z.; Mohamed, Z.; Ahmadiani, A. LPS preconditioning attenuates apoptosis mechanism by inhibiting NF-κB and caspase-3 activity: TLR4 Pre-activation in the signaling pathway of LPS-induced neuroprotection. Mol. Neurobiol. 2021, 58, 2407–2422. [Google Scholar] [CrossRef]
- Zajączkowska, R.; Kocot-Kępska, M.; Leppert, W.; Wrzosek, A.; Mika, J.; Wordliczek, J. Mechanisms of Chemotherapy-Induced Peripheral Neuropathy. Int. J. Mol. Sci. 2019, 20, 1451. [Google Scholar] [CrossRef] [Green Version]
- Rosati, R.; Shahab, M.; Ramkumar, V.; Jamesdaniel, S. Lmo4 deficiency enhances susceptibility to cisplatin-induced cochlear apoptosis and hearing loss. Mol. Neurobiol. 2021, 58, 2019–2029. [Google Scholar] [CrossRef]
- Yu, D.; Gu, J.; Chen, Y.; Kang, W.; Wang, X.; Wu, H. Current strategies to combat cisplatin-induced ototoxicity. Front. Pharmacol. 2020, 11, 999. [Google Scholar] [CrossRef]
- Gentilin, E.; Simoni, E.; Candito, M.; Cazzador, D.; Astolfi, L. Cisplatin-induced ototoxicity: Updates on molecular targets. Trends Mol. Med. 2019, 25, 1123–1132. [Google Scholar] [CrossRef]
- Ciarimboli, G. Membrane transporters as mediators of cisplatin side-effects. Anticancer Res. 2014, 34, 547–550. [Google Scholar] [CrossRef]
- Tang, Q.; Wang, X.; Jin, H.; Mi, Y.; Liu, L.; Dong, M.; Chen, Y.; Zou, Z. Cisplatin-induced ototoxicity: Updates on molecular mechanisms and otoprotective strategies. Eur. J. Pharm. Biopharm. 2021, 163, 60–71. [Google Scholar] [CrossRef] [PubMed]
- Rathinam, R.; Ghosh, S.; Neumann, W.L.; Jamesdaniel, S. Cisplatin-induced apoptosis in auditory, renal, and neuronal cells is associated with nitration and downregulation of LMO4. Cell Death Discov. 2015, 1, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, S.; Egashira, N. Drug repositioning for the prevention and treatment of chemotherapy-induced peripheral neuropathy: A mechanism- and screening-based strategy. Front. Pharmacol. 2020, 11, 607780. [Google Scholar] [CrossRef] [PubMed]
- Wasilewski, A.; Mohile, N. Meet the expert: How I treat chemotherapy-induced peripheral neuropathy. J. Geriatr. Oncol. 2021, 12, 1–5. [Google Scholar] [CrossRef]
- Jamesdaniel, S.; Rathinam, R.; Neumann, W.L. Targeting nitrative stress for attenuating cisplatin-induced downregulation of cochlear LIM domain only 4 and ototoxicity. Redox Biol. 2016, 10, 257–265. [Google Scholar] [CrossRef] [Green Version]
- Sang, M.; Ma, L.; Zhou, X.; Gao, W.; Geng, C. LIM-domain-only proteins: Multifunctional nuclear transcription coregulators that interacts with diverse proteins. Mol. Biol. Rep. 2014, 41, 1067–1073. [Google Scholar] [CrossRef]
- Kashani, A.H.; Qiu, Z.; Jurata, L.; Lee, S.K.; Pfaff, S.; Goebbels, S.; Nave, K.A.; Ghosh, A. Calcium activation of the LMO4 transcription complex and its role in the patterning of thalamocortical connections. J. Neurosci. 2006, 26, 8398–8408. [Google Scholar] [CrossRef] [Green Version]
- Rathinam, R.; Rosati, R.; Jamesdaniel, S. CRISPR/Cas9-mediated knockout of Lim-domain only four retards organ of corti cell growth. J. Cell. Biochem. 2018, 119, 3545–3553. [Google Scholar] [CrossRef]
- Deng, M.; Luo, X.J.; Pan, L.; Yang, H.; Xie, X.; Liang, G.; Huang, L.; Hu, F.; Kiernan, A.E.; Gan, L. LMO4 functions as a negative regulator of sensory organ formation in the mammalian cochlea. J. Neurosci. 2014, 34, 10072–10077. [Google Scholar] [CrossRef]
- Qin, Z.; Zhou, X.; Gomez-Smith, M.; Pandey, N.R.; Lee, K.F.; Lagace, D.C.; Béïque, J.C.; Chen, H.H. LIM domain only 4 (LMO4) regulates calcium-induced calcium release and synaptic plasticity in the hippocampus. J. Neurosci. 2012, 32, 4271–4283. [Google Scholar] [CrossRef] [Green Version]
- Jamesdaniel, S. Downstream targets of Lmo4 are modulated by cisplatin in the inner ear of Wistar rats. PLoS ONE 2014, 9, e115263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosati, R.; Shahab, M.; Neumann, W.L.; Jamesdaniel, S. Inhibition of protein nitration prevents cisplatin-induced inactivation of STAT3 and promotes anti-apoptotic signaling in organ of Corti cells. Exp. Cell Res. 2019, 381, 105–111. [Google Scholar] [CrossRef]
- Rivolta, M.N.; Grix, N.; Lawlor, P.; Ashmore, J.F.; Jagger, D.J.; Holley, M.C. Auditory hair cell precursors immortalized from the mammalian inner ear. Proc. Biol. Sci. 1998, 265, 1595–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R. Hot spots and future directions of research on the neuroprotective effects of nimodipine. Neural Regen. Res. 2014, 9, 1933–1938. [Google Scholar] [CrossRef] [PubMed]
- Herzfeld, E.; Speh, L.; Strauss, C.; Scheller, C. Nimodipine but not nifedipine promotes expression of fatty acid 2-hydroxylase in a surgical stress model based on neuro2a cells. Int. J. Mol. Sci. 2017, 18, 964. [Google Scholar] [CrossRef] [Green Version]
- Herzfeld, E.; Strauss, C.; Simmermacher, S.; Bork, K.; Horstkorte, R.; Dehghani, F.; Scheller, C. Investigation of the neuroprotective impact of nimodipine on neuro2a cells by means of a surgery-like stress model. Int. J. Mol. Sci. 2014, 15, 18453–18465. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Dong, Q.; Zhou, X.N. LMO4 promotes the invasion and proliferation of gastric cancer by activating PI3K-Akt-mTOR signaling. Am. J. Transl. Res. 2019, 11, 6534–6543. [Google Scholar]
- Wang, L.; Yin, Z.; Wang, F.; Han, Z.; Wang, Y.; Huang, S.; Hu, T.; Guo, M.; Lei, P. Hydrogen exerts neuroprotection by activation of the miR-21/PI3K/AKT/GSK-3β pathway in an in vitro model of traumatic brain injury. J. Cell. Mol. Med. 2020, 24, 4061–4071. [Google Scholar] [CrossRef] [Green Version]
- Pregi, N.; Belluscio, L.M.; Berardino, B.G.; Castillo, D.S.; Cánepa, E.T. Oxidative stress-induced CREB upregulation promotes DNA damage repair prior to neuronal cell death protection. Mol. Cell. Biochem. 2017, 425, 9–24. [Google Scholar] [CrossRef]
- Petersen, O.H.; Verkhratsky, A. Calcium and ATP control multiple vital functions. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unel, C.C.; Erol, K. The role of ionic homeostasis in cisplatin-induced neurotoxicity: A preliminary study. Eurasian J. Med. 2018, 50, 81–85. [Google Scholar] [CrossRef] [PubMed]
- Duchen, M.R. Mitochondria, calcium-dependent neuronal death and neurodegenerative disease. Pflügers Arch.-Eur. J. Physiol. 2012, 464, 111–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petremann, M.; Tran Van Ba, C.; Broussy, A.; Romanet, C.; Dyhrfjeld-Johnsen, J. Oral administration of clinical stage drug candidate SENS-401 effectively reduces cisplatin-induced hearing loss in rats. Otol. Neurotol. 2017, 38, 1355–1361. [Google Scholar] [CrossRef]
- Petremann, M.; Romanet, C.; Broussy, A.; Van Ba, C.T.; Poli, S.; Dyhrfjeld-Johnsen, J. SENS-401 effectively reduces severe acoustic trauma-induced hearing loss in male rats with twice daily administration delayed up to 96 hours. Otol. Neurotol. 2019, 40, 254–263. [Google Scholar] [CrossRef]
- Fouladi, M.; Chintagumpala, M.; Ashley, D.; Kellie, S.; Gururangan, S.; Hassall, T.; Gronewold, L.; Stewart, C.F.; Wallace, D.; Broniscer, A.; et al. Amifostine protects against cisplatin-induced ototoxicity in children with average-risk medulloblastoma. J. Clin. Oncol. 2008, 26, 3749–3755. [Google Scholar] [CrossRef] [Green Version]
- Gurney, J.G.; Bass, J.K.; Onar-Thomas, A.; Huang, J.; Chintagumpala, M.; Bouffet, E.; Hassall, T.; Gururangan, S.; Heath, J.A.; Kellie, S.; et al. Evaluation of amifostine for protection against cisplatin-induced serious hearing loss in children treated for average-risk or high-risk medulloblastoma. Neuro Oncol. 2014, 16, 848–855. [Google Scholar] [CrossRef] [Green Version]
- Freyer, D.R.; Brock, P.R.; Chang, K.W.; Dupuis, L.L.; Epelman, S.; Knight, K.; Mills, D.; Phillips, R.; Potter, E.; Risby, D.; et al. Prevention of cisplatin-induced ototoxicity in children and adolescents with cancer: A clinical practice guideline. Lancet Child. Adolesc. Health 2020, 4, 141–150. [Google Scholar] [CrossRef]
- Fox, E.; Levin, K.; Zhu, Y.; Segers, B.; Balamuth, N.; Womer, R.; Bagatell, R.; Balis, F. Pantoprazole, an inhibitor of the organic cation transporter 2, does not ameliorate cisplatin-related ototoxicity or nephrotoxicity in children and adolescents with newly diagnosed osteosarcoma treated with methotrexate, doxorubicin, and cisplatin. Oncologist 2018, 23, e762–e779. [Google Scholar] [CrossRef] [Green Version]
- Waissbluth, S.; Salehi, P.; He, X.; Daniel, S.J. Systemic dexamethasone for the prevention of cisplatin-induced ototoxicity. Eur. Arch. Otorhinolaryngol. 2013, 270, 1597–1605. [Google Scholar] [CrossRef]
- Simsek, G.; Taş, B.M.; Muluk, N.B.; Azman, M.; Kılıç, R. Comparison of the protective efficacy between intratympanic dexamethasone and resveratrol treatments against cisplatin-induced ototoxicity: An experimental study. Eur. Arch. Oto-Rhino-Laryngol. 2019, 276, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Wang, X.; Chen, D.; Lin, X.; Yu, D.; Wu, H. Dexamethasone loaded nanoparticles exert protective effects against cisplatin-induced hearing loss by systemic administration. Neurosci. Lett. 2016, 619, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Gu, J.; Liu, J.; Tong, L.; Shi, F.; Wang, X.; Yu, D.; Wu, H. Dexamethasone-loaded injectable silk-polyethylene glycol hydrogel alleviates cisplatin-induced ototoxicity. Int. J. Nanomed. 2019, 14, 4211–4227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saliba, I.; El Fata, F.; Ouelette, V.; Robitaille, Y. Are intratympanic injections of N-acetylcysteine and methylprednisolone protective against Cisplatin-induced ototoxicity? J. Otolaryngol. Head Neck Surg. 2010, 39, 236–243. [Google Scholar] [PubMed]
- Somdaş, M.A.; Güntürk, İ.; Balcıoğlu, E.; Avcı, D.; Yazıcı, C.; Özdamar, S. Protective effect of N-acetylcysteine against cisplatin ototoxicity in rats: A study with hearing tests and scanning electron microscopy. Braz J. Otorhinolaryngol. 2020, 86, 30–37. [Google Scholar] [CrossRef]
- Esen, E.; Özdoğan, F.; Gürgen, S.G.; Özel, H.E.; Başer, S.; Genç, S.; Selçuk, A. Ginkgo biloba and lycopene are effective on cisplatin induced ototoxicity? J. Int. Adv. Otol. 2018, 14, 22–26. [Google Scholar] [CrossRef] [PubMed]
- Freyer, D.R.; Chen, L.; Krailo, M.D.; Knight, K.; Villaluna, D.; Bliss, B.; Pollock, B.H.; Ramdas, J.; Lange, B.; Van Hoff, D.; et al. Effects of sodium thiosulfate versus observation on development of cisplatin-induced hearing loss in children with cancer (ACCL0431): A multicentre, randomised, controlled, open-label, phase 3 trial. Lancet Oncol. 2017, 18, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Brock, P.R.; Maibach, R.; Childs, M.; Rajput, K.; Roebuck, D.; Sullivan, M.J.; Laithier, V.; Ronghe, M.; Dall’Igna, P.; Hiyama, E.; et al. Sodium thiosulfate for protection from cisplatin-induced hearing loss. N. Engl. J. Med. 2018, 378, 2376–2385. [Google Scholar] [CrossRef] [Green Version]
- Sarpong-Kumankomah, S.; Gailer, J. Application of a novel metallomics tool to probe the fate of metal-based anticancer drugs in blood plasma: Potential, challenges and prospects. Curr. Top. Med. Chem. 2021, 21, 48–58. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UB/OC−1 | UB/OC−2 | |
|---|---|---|
| Gene Name | Factor of Upregulation after Differentation | |
| Brn3.1 | 2.5 ± 0.9 | 2.8 ± 1.9 |
| Myo6 | 1.5 ± 0.9 | 2.7 ± 2.2 |
| Myo7a | 1.1 ± 0.2 | 9.9 ± 4.3 |
| α9AChR | 2.4 ± 0.9 | 5.3 ± 2.9 |
| Antibody | Isotype | Dilution | Dilution Buffer | Manufacturer |
|---|---|---|---|---|
| Phospho-Akt (Ser473) (D9E) XP® #4060 | Rabbit IgG | 1:1000 | 5% BSA in TBS-T | Cell Signaling Technology (Danvers, MA, USA) |
| Akt (pan) (40D4) #2920 | Mouse IgG1 | 1:2000 | 5% MP in TBS-T | |
| Phospho-CREB (Ser133) (87G3) #9198 | Rabbit IgG | 1:1000 | 5% MP in TBS-T | |
| CREB (48H2) #9197 | Rabbit IgG | 1:1000 | 5% BSA in TBS-T | |
| Phospho-Stat3 (Tyr705) (3E2) #9138 | Mouse IgG1 | 1:1000 | 5% MP in TBS-T | |
| Stat3 (124H6) #9139 | Mouse IgG2a | 1:1000 | 5% MP in TBS-T | |
| LMO4 (D6V4Z) #81428 | Rabbit IgG | 1:1000 | 5% BSA in TBS-T | |
| GAPDH (14C10) #2118 | Rabbit IgG | 1:1000 | 5% BSA in TBS-T | |
| Anti-mouse IgG, HRP-linked Antibody | Horse | 1:1000 | 2% MP in TBS-T | |
| Anti-rabbit IgG, HRP-linked Antibody | Goat | 1:1000 | 2% MP in TBS-T |
| Gene Name | Oligo Sequence 5′ to 3′ (Forward, Reverse) | Annealing Temperature | Reference Sequence | Species |
|---|---|---|---|---|
| Brn3.1 | TTCAACGGCAGTGAGCGTAA, ACAGAACCAGACCCTCACCA | 60 °C | NM_138945.2 | Mus musculus |
| Myo6 | CATGGCACTCCGAAGAGGT, GGGATTCTGCTGAGGTGAATTG | 60 °C | NM_001039546.2 | |
| Myo7a | ACTGCTCTGTGAGACATCGC, ACCAGGAAGGCCACAACAAA | 60 °C | NM_001256081.1 | |
| α9AChR | TGCACGCTATGAAGCACTGA, CGAATGCCTACCAACCCACT | 60 °C | NM_001081104.1 | |
| GAPDH | GCACAGTCAAGGCCGAGAAT, GCCTTCTCCATGGTGGTGAA | 60 °C | NM_001289726 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fritzsche, S.; Strauss, C.; Scheller, C.; Leisz, S. Nimodipine Treatment Protects Auditory Hair Cells from Cisplatin-Induced Cell Death Accompanied by Upregulation of LMO4. Int. J. Mol. Sci. 2022, 23, 5780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105780
Fritzsche S, Strauss C, Scheller C, Leisz S. Nimodipine Treatment Protects Auditory Hair Cells from Cisplatin-Induced Cell Death Accompanied by Upregulation of LMO4. International Journal of Molecular Sciences. 2022; 23(10):5780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105780
Chicago/Turabian StyleFritzsche, Saskia, Christian Strauss, Christian Scheller, and Sandra Leisz. 2022. "Nimodipine Treatment Protects Auditory Hair Cells from Cisplatin-Induced Cell Death Accompanied by Upregulation of LMO4" International Journal of Molecular Sciences 23, no. 10: 5780. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105780