
The Cell Surface Heparan Sulfate Proteoglycan Syndecan-3 Promotes Ovarian Cancer Pathogenesis

 , , ,
, , ,  ,
, 
Abstract
:1. Introduction
2. Results
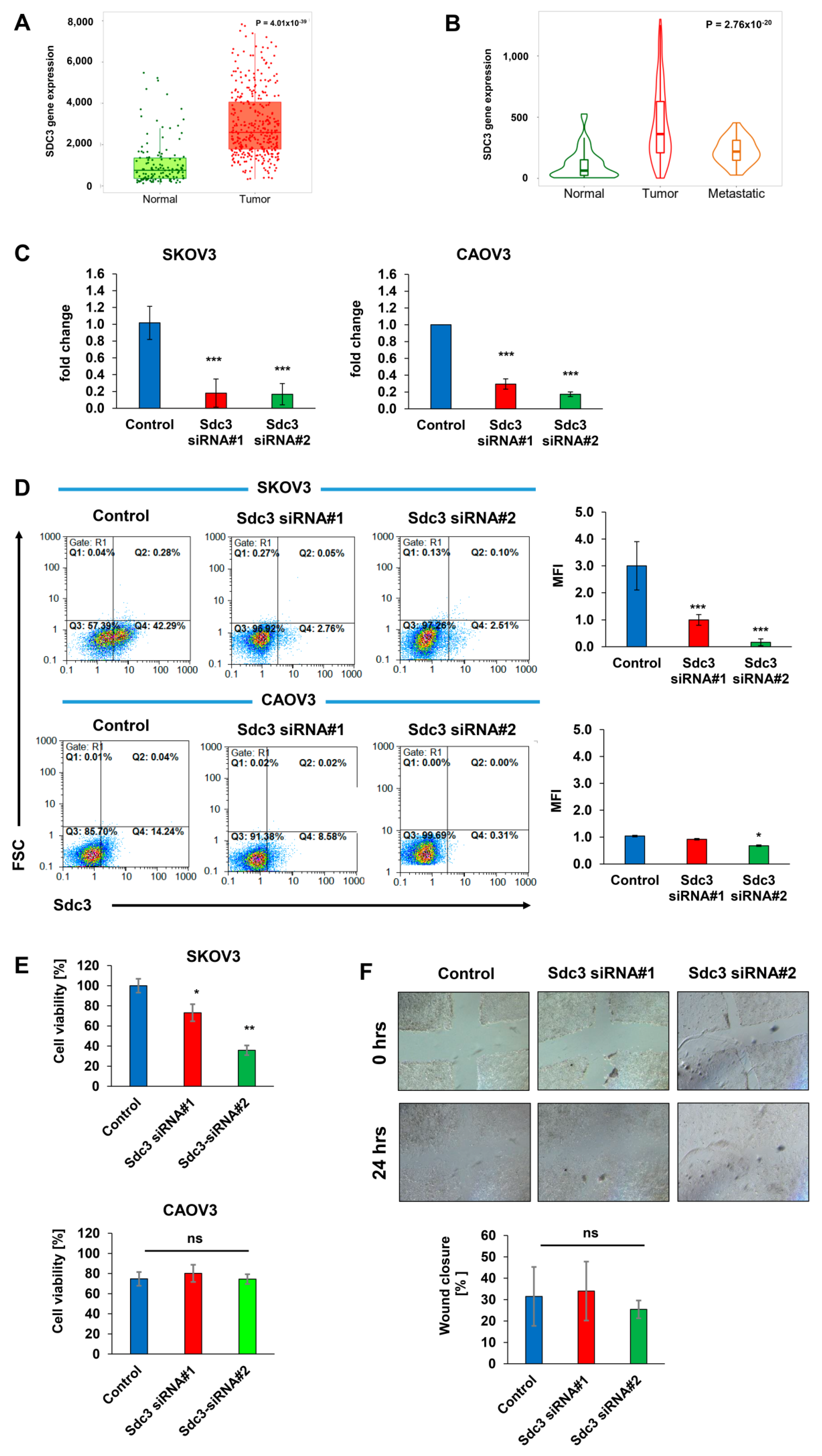
2.1. Syndecan-3 Is Overexpressed in Ovarian Cancer
2.2. Syndecan-3 Depletion Affects Colony Formation and 3D Spheroid Growth of Human SKOV3 and CAOV3 Ovarian Cancer Cells
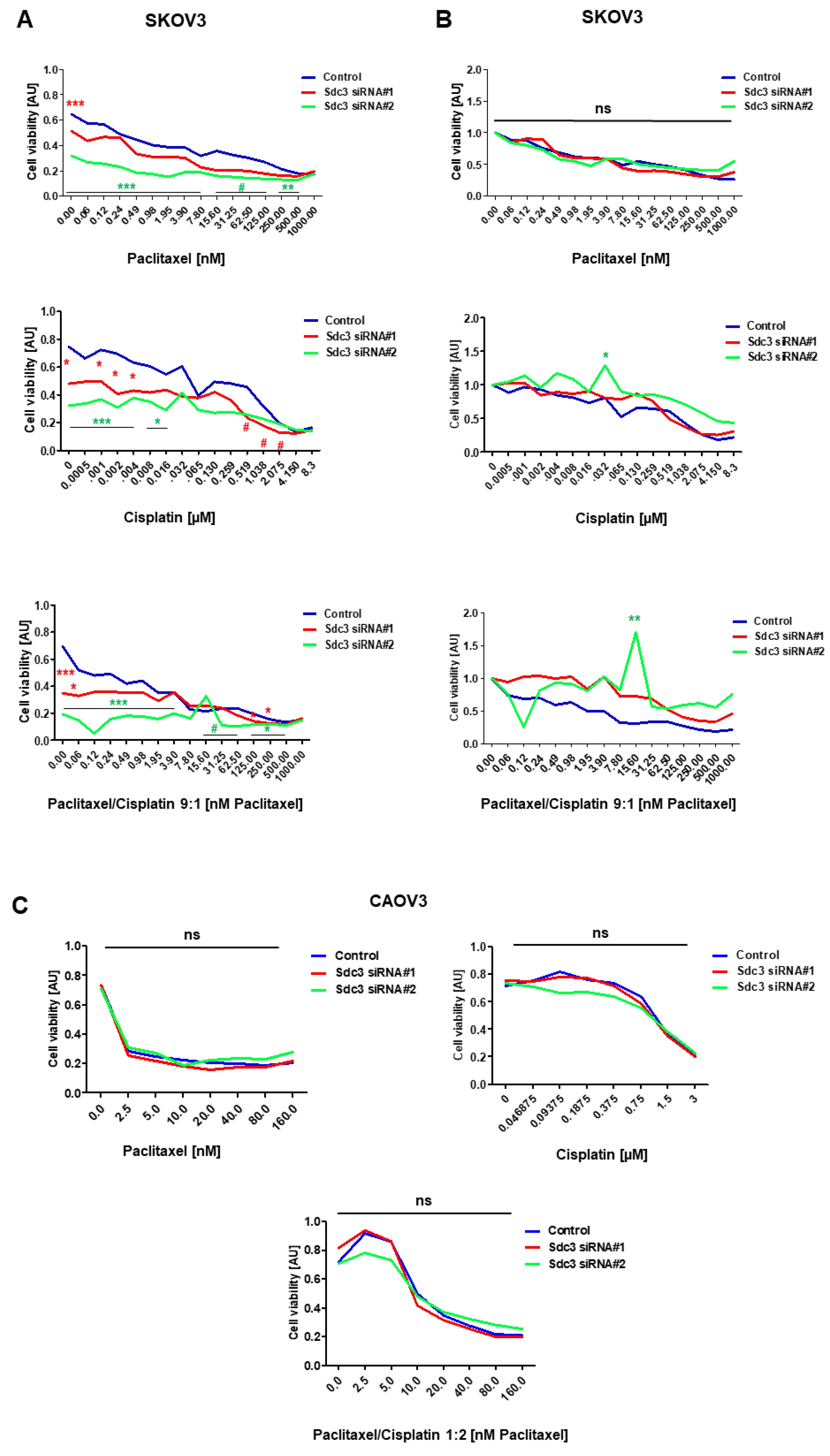
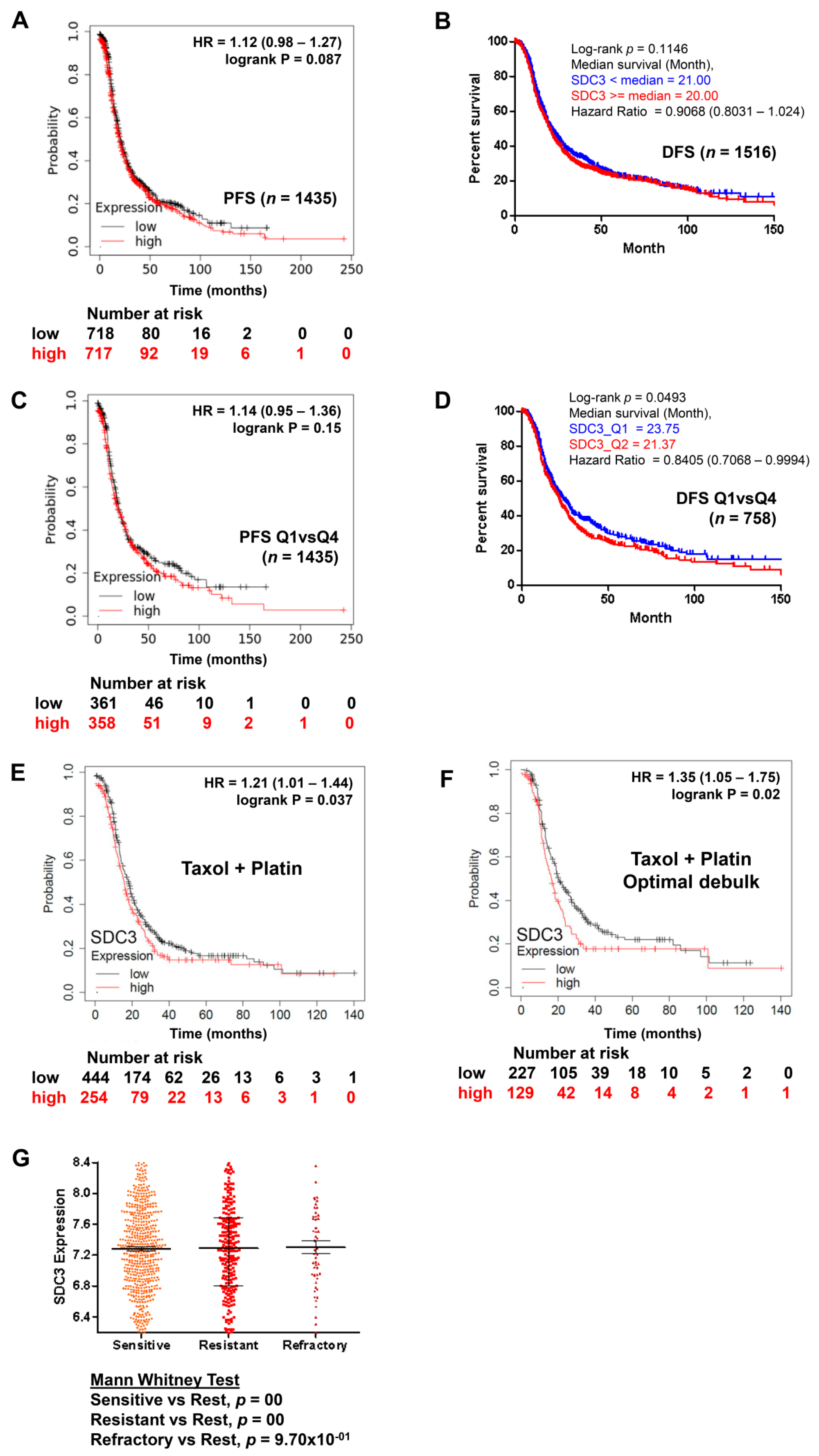
2.3. Impact of Syndecan-3 on the Chemotherapy Response in Ovarian Cancer
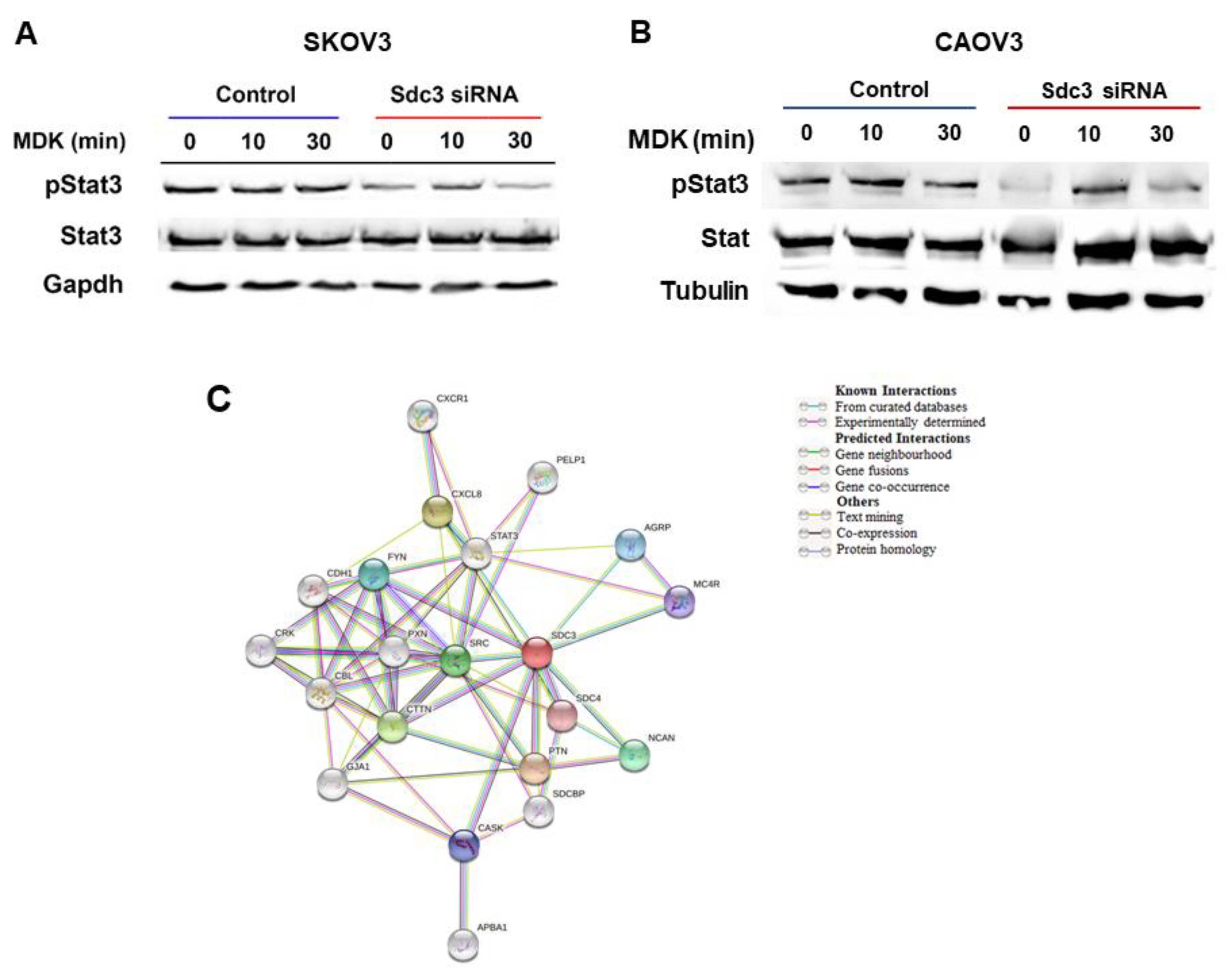
2.4. Syndecan-3 Shows Functional Interactions with Stat3 Signaling and Is Associated with Stemness-Associated Signaling Pathways in STRING Analysis
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. siRNA Transfection
4.3. Total RNA Extraction and cDNA Synthesis
4.4. Quantitative Real-Time PCR
4.5. Western Blotting
4.6. Flow Cytometry
4.7. MTT Cell Viability and Chemosensitivity Assay
4.8. Scratch Wound Assay
4.9. Hanging Drop Assay for 3D Sphere Formation
4.10. Cell Colony Formation
4.11. TNM, Kaplan–Meier Plot and STRING Analysis
4.12. Microarray Gene Expression Database of Epithelial Ovarian Cancer Subtype CSIOVDB
4.13. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bernfield, M.; Götte, M.; Park, P.W.; Reizes, O.; Fitzgerald, M.L.; Lincecum, J.; Zako, M. Functions of Cell Surface Heparan Sulfate Proteoglycans. Annu. Rev. Biochem. 1999, 68, 729–777. [Google Scholar] [CrossRef]
- Karamanos, N.K.; Piperigkou, Z.; Theocharis, A.D.; Watanabe, H.; Franchi, M.; Baud, S.; Brézillon, S.; Götte, M.; Passi, A.; Vigetti, D.; et al. Proteoglycan Chemical Diversity Drives Multifunctional Cell Regulation and Therapeutics. Chem. Rev. 2018, 118, 9152–9232. [Google Scholar] [CrossRef]
- Espinoza-Sánchez, N.A.; Götte, M. Role of cell surface proteoglycans in cancer immunotherapy. Semin. Cancer Biol. 2020, 62, 48–67. [Google Scholar] [CrossRef]
- Gondelaud, F.; Ricard-Blum, S. Structures and interactions of syndecans. FEBS J. 2019, 286, 2994–3007. [Google Scholar] [CrossRef] [Green Version]
- Piperigkou, Z.; Mohr, B.; Karamanos, N.; Götte, M. Shed proteoglycans in tumor stroma. Cell Tissue Res. 2016, 365, 643–655. [Google Scholar] [CrossRef]
- Hassan, N.; Greve, B.; Espinoza-Sánchez, N.A.; Götte, M. Cell-surface heparan sulfate proteoglycans as multifunctional integrators of signaling in cancer. Cell. Signal. 2020, 77, 109822. [Google Scholar] [CrossRef]
- Vitale, D.L.; Katakam, S.K.; Greve, B.; Jang, B.; Oh, E.; Alaniz, L.; Götte, M. Proteoglycans and glycosaminoglycans as regulators of cancer stem cell function and therapeutic resistance. FEBS J. 2019, 286, 2870–2882. [Google Scholar] [CrossRef] [Green Version]
- SEER. Cancer of the Ovary—Cancer Stat Facts. Available online: https://seer.cancer.gov/statfacts/html/ovary.html (accessed on 11 June 2021).
- Cancer Facts & Figures 2020. Available online: https://www.cancer.org/content/dam/cancer-org/research/cancer-facts-and-statistics/annual-cancer-facts-and-figures/2020/cancer-facts-and-figures-2020.pdf (accessed on 12 June 2021).
- Armstrong, D.K.; Alvarez, R.D.; Bakkum-Gamez, J.N.; Barroilhet, L.; Behbakht, K.; Berchuck, A.; Chen, L.-M.; Cristea, M.; DeRosa, M.; Eisenhauer, E.L.; et al. Ovarian Cancer, Version 2.2020, NCCN Clinical Practice Guidelines in Oncology. J. Natl. Compr. Cancer Netw. 2021, 19, 191–226. [Google Scholar] [CrossRef]
- Burger, R.A.; Brady, M.F.; Bookman, M.A.; Fleming, G.F.; Monk, B.J.; Huang, H.; Mannel, R.S.; Homesley, H.D.; Fowler, J.; Greer, B.E.; et al. Incorporation of Bevacizumab in the Primary Treatment of Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2473–2483. [Google Scholar] [CrossRef] [Green Version]
- Perren, T.J.; Swart, A.M.; Pfisterer, J.; Ledermann, J.A.; Pujade-Lauraine, E.; Kristensen, G.; Carey, M.S.; Beale, P.; Cervantes, A.; Kurzeder, C.; et al. A Phase 3 Trial of Bevacizumab in Ovarian Cancer. N. Engl. J. Med. 2011, 365, 2484–2496. [Google Scholar] [CrossRef] [Green Version]
- Moore, K.; Colombo, N.; Scambia, G.; Kim, B.-G.; Oaknin, A.; Friedlander, M.; Lisyanskaya, A.; Floquet, A.; Leary, A.; Sonke, G.S.; et al. Maintenance Olaparib in Patients with Newly Diagnosed Advanced Ovarian Cancer. N. Engl. J. Med. 2018, 379, 2495–2505. [Google Scholar] [CrossRef]
- Herzog, T.J.; Tewari, K.S. Evolution of Chemosensitivity and Resistance Assays as Predictors of Clinical Outcomes in Epithelial Ovarian Cancer Patients. Curr. Pharm. Des. 2016, 22, 4717–4728. [Google Scholar] [CrossRef]
- Davies, E.J.; Blackhall, F.; Shanks, J.H.; David, G.; McGown, A.T.; Swindell, R.; Slade, R.J.; Martin-Hirsch, P.; Gallagher, J.T.; Jayson, G.C. Distribution and Clinical Significance of Heparan Sulfate Proteoglycans in Ovarian Cancer. Clin. Cancer Res. 2004, 10, 5178–5186. [Google Scholar] [CrossRef] [Green Version]
- Whitworth, M.K.; Backen, A.C.; Clamp, A.R.; Wilson, G.; Mc Vey, R.; Friedl, A.; Rapraeger, A.C.; David, G.; McGown, A.T.; Slade, R.J.; et al. Regulation of Fibroblast Growth Factor-2 Activity by Human Ovarian Cancer Tumor Endothelium. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2005, 11, 4282–4288. [Google Scholar] [CrossRef] [Green Version]
- Kulbe, H.; Otto, R.; Darb-Esfahani, S.; Lammert, H.; Abobaker, S.; Welsch, G.; Chekerov, R.; Schäfer, R.; Dragun, D.; Hummel, M.; et al. Discovery and Validation of Novel Biomarkers for Detection of Epithelial Ovarian Cancer. Cells 2019, 8, 713. [Google Scholar] [CrossRef] [Green Version]
- Bartha, Á.; Győrffy, B. TNMplot.com: A Web Tool for the Comparison of Gene Expression in Normal, Tumor and Metastatic Tissues. Int. J. Mol. Sci. 2021, 22, 2622. [Google Scholar] [CrossRef]
- Fogh, G.; Trempe, J. New Human Tumor Cell Lines. In Human Tumor Cells In Vitro; Fogh, J., Ed.; Springer: Boston, MA, USA, 1975. [Google Scholar] [CrossRef]
- Valla, S.; Hassan, N.; Vitale, D.; Madanes, D.; Spinelli, F.; Teixeira, F.; Greve, B.; Espinoza-Sánchez, N.; Cristina, C.; Alaniz, L.; et al. Syndecan-1 Depletion Has a Differential Impact on Hyaluronic Acid Metabolism and Tumor Cell Behavior in Luminal and Triple-Negative Breast Cancer Cells. Int. J. Mol. Sci. 2021, 22, 5874. [Google Scholar] [CrossRef]
- Teixeira, F.C.O.B.; Kumar, A.V.; Katakam, S.K.; Cocola, C.; Pelucchi, P.; Graf, M.; Kiesel, L.; Reinbold, R.; Pavão, M.S.G.; Greve, B.; et al. The Heparan Sulfate Sulfotransferases HS2ST1 and HS3ST2 Are Novel Regulators of Breast Cancer Stem-Cell Properties. Front. Cell Dev. Biol. 2020, 8, 559554. [Google Scholar] [CrossRef]
- Katakam, S.K.; Tria, V.; Sim, W.; Yip, G.W.; Molgora, S.; Karnavas, T.; Elghonaimy, E.A.; Pelucchi, P.; Piscitelli, E.; Ibrahim, S.A.; et al. The heparan sulfate proteoglycan syndecan-1 regulates colon cancer stem cell function via a focal adhesion kinase—Wnt signaling axis. FEBS J. 2021, 288, 486–506. [Google Scholar] [CrossRef]
- Katakam, S.K.; Pelucchi, P.; Cocola, C.; Reinbold, R.; Vlodavsky, I.; Greve, B.; Götte, M. Syndecan-1-Dependent Regulation of Heparanase Affects Invasiveness, Stem Cell Properties, and Therapeutic Resistance of Caco2 Colon Cancer Cells. Front. Oncol. 2020, 10, 774. [Google Scholar] [CrossRef]
- Stejskalová, A.; Fincke, V.; Nowak, M.; Schmidt, Y.; Borrmann, K.; von Wahlde, M.-K.; Schäfer, S.D.; Kiesel, L.; Greve, B.; Götte, M. Collagen I triggers directional migration, invasion and matrix remodeling of stroma cells in a 3D spheroid model of endometriosis. Sci. Rep. 2021, 11, 4115. [Google Scholar] [CrossRef]
- Kuehn, J.; Espinoza-Sanchez, N.A.; Teixeira, F.C.O.B.; Pavão, M.S.G.; Kiesel, L.; Győrffy, B.; Greve, B.; Götte, M. Prognostic significance of hedgehog signaling network-related gene expression in breast cancer patients. J. Cell. Biochem. 2021, 122, 577–597. [Google Scholar] [CrossRef]
- Kinnunen, T.K.; Kaksonen, M.; Saarinen, J.; Kalkkinen, N.; Peng, H.B.; Rauvala, H. Cortactin-Src Kinase Signaling Pathway Is Involved in N-syndecan-dependent Neurite Outgrowth. J. Biol. Chem. 1998, 273, 10702–10708. [Google Scholar] [CrossRef] [Green Version]
- Gyorffy, B.; Lánczky, A.; Szallasi, Z. Implementing an online tool for genome-wide validation of survival-associated biomarkers in ovarian-cancer using microarray data from 1287 patients. Endocr. Relat. Cancer 2012, 19, 197–208. [Google Scholar] [CrossRef] [Green Version]
- Tan, T.Z.; Yang, H.; Ye, J.; Low, J.; Choolani, M.; Tan, D.; Thiery, J.-P.; Huang, R.Y.-J. CSIOVDB: A microarray gene expression database of epithelial ovarian cancer subtype. Oncotarget 2015, 6, 43843–43852. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, T.; Kadomatsu, K.; Okamoto, T.; Ichihara-Tanaka, K.; Kojima, T.; Saito, H.; Tomoda, Y.; Muramatsu, T. Expression of syndecan-1 and -3 during embryogenesis of the central nervous system in relation to binding with midkine. J. Biochem. 1997, 121, 197–205. [Google Scholar]
- Cernkovich, E.R.; Deng, J.; Hua, K.; Harp, J.B. Midkine Is an Autocrine Activator of Signal Transducer and Activator of Transcription 3 in 3T3-L1 Cells. Endocrinology 2007, 148, 1598–1604. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING v11: Protein–protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Prieto-Fernández, E.; Egia-Mendikute, L.; Bosch, A.; Del Río, A.G.; Jimenez-Lasheras, B.; Antoñana-Vildosola, A.; Lee, S.Y.; Palazon, A. Hypoxia Promotes Syndecan-3 Expression in the Tumor Microenvironment. Front. Immunol. 2020, 11, 586977. [Google Scholar] [CrossRef]
- Yao, J.; Zhang, L.-L.; Huang, X.-M.; Li, W.-Y.; Gao, S.-G. Pleiotrophin and N-syndecan promote perineural invasion and tumor progression in an orthotopic mouse model of pancreatic cancer. World J. Gastroenterol. 2017, 23, 3907–3914. [Google Scholar] [CrossRef]
- Eustace, A.D.; McNaughton, E.F.; King, S.; Kehoe, O.; Kungl, A.; Mattey, D.; Nobbs, A.H.; Williams, N.; Middleton, J. Soluble syndecan-3 binds chemokines, reduces leukocyte migration in vitro and ameliorates disease severity in models of rheumatoid arthritis. Arthritis Res. Ther. 2019, 21, 172. [Google Scholar] [CrossRef] [Green Version]
- Bespalov, M.M.; Sidorova, Y.A.; Tumova, S.; Ahonen-Bishopp, A.; Magalhães, A.C.; Kulesskiy, E.; Paveliev, M.; Rivera, C.; Rauvala, H.; Saarma, M. Heparan sulfate proteoglycan syndecan-3 is a novel receptor for GDNF, neurturin, and artemin. J. Cell Biol. 2011, 192, 153–169. [Google Scholar] [CrossRef] [Green Version]
- Kurosawa, N.; Chen, G.-Y.; Kadomatsu, K.; Ikematsu, S.; Sakuma, S.; Muramatsu, T. Glypican-2 binds to midkine: The role of glypican-2 in neuronal cell adhesion and neurite outgrowth. Glycoconj. J. 2001, 18, 499–507. [Google Scholar] [CrossRef]
- Yao, J.; Li, W.-Y.; Li, S.-G.; Feng, X.-S.; Gao, S.-G. Midkine promotes perineural invasion in human pancreatic cancer. World J. Gastroenterol. 2014, 20, 3018–3024. [Google Scholar] [CrossRef]
- Ponandai-Srinivasan, S.; Saare, M.; Boggavarapu, N.R.; Frisendahl, C.; Ehrström, S.; Riethmüller, C.; García-Uribe, P.A.; Rettkowski, J.; Iyengar, A.; Salumets, A.; et al. Syndecan-1 modulates the invasive potential of endometrioma via TGF-β signalling in a subgroup of women with endometriosis. Hum. Reprod. 2020, 35, 2280–2293. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Gadalla, R.; El-Ghonaimy, E.A.; Samir, O.; Mohamed, H.T.; Hassan, H.; Greve, B.; El-Shinawi, M.; Mohamed, M.M.; Götte, M. Syndecan-1 is a novel molecular marker for triple negative inflammatory breast cancer and modulates the cancer stem cell phenotype via the IL-6/STAT3, Notch and EGFR signaling pathways. Mol. Cancer 2017, 16, 57. [Google Scholar] [CrossRef] [Green Version]
- Jones, F.K.; Stefan, A.; Kay, A.G.; Hyland, M.; Morgan, R.; Forsyth, N.R.; Pisconti, A.; Kehoe, O. Syndecan-3 regulates MSC adhesion, ERK and AKT signalling in vitro and its deletion enhances MSC efficacy in a model of inflammatory arthritis in vivo. Sci. Rep. 2020, 10, 20487. [Google Scholar] [CrossRef]
- Arokiasamy, S.; Balderstone, M.J.M.; De Rossi, G.; Whiteford, J.R. Syndecan-3 in Inflammation and Angiogenesis. Front. Immunol. 2019, 10, 3031. [Google Scholar] [CrossRef] [Green Version]
- Escalona, R.M.; Bilandzic, M.; Western, P.; Kadife, E.; Kannourakis, G.; Findlay, J.K.; Ahmed, N. TIMP-2 regulates proliferation, invasion and STAT3-mediated cancer stem cell-dependent chemoresistance in ovarian cancer cells. BMC Cancer 2020, 20, 960. [Google Scholar] [CrossRef]
- Martincuks, A.; Li, P.-C.; Zhao, Q.; Zhang, C.; Li, Y.-J.; Yu, H.; Rodriguez-Rodriguez, L. CD44 in Ovarian Cancer Progression and Therapy Resistance—A Critical Role for STAT3. Front. Oncol. 2020, 10, 589601. [Google Scholar] [CrossRef]
- Gong, T.; Zhang, P.; Jia, L.; Pan, Y. Suppression of ovarian cancer by low-intensity ultrasound through depletion of IL-6/STAT3 inflammatory pathway-maintained cancer stemness. Biochem. Biophys. Res. Commun. 2020, 526, 820–826. [Google Scholar] [CrossRef]
- Ibrahim, S.A.; Hassan, H.; Vilardo, L.; Kumar, S.K.; Kumar, A.V.; Kelsch, R.; Schneider, C.; Kiesel, L.; Eich, H.T.; Zucchi, I.; et al. Syndecan-1 (CD138) Modulates Triple-Negative Breast Cancer Stem Cell Properties via Regulation of LRP-6 and IL-6-Mediated STAT3 Signaling. PLoS ONE 2013, 8, e85737. [Google Scholar] [CrossRef]
- Pisconti, A.; Cornelison, D.; Olguín, H.C.; Antwine, T.L.; Olwin, B.B. Syndecan-3 and Notch cooperate in regulating adult myogenesis. J. Cell Biol. 2010, 190, 427–441. [Google Scholar] [CrossRef] [Green Version]
- Casar, J.C.; Cabello-Verrugio, C.; Olguin, H.; Aldunate, R.; Inestrosa, N.C.; Brandan, E. Heparan sulfate proteoglycans are increased during skeletal muscle regeneration: Requirement of syndecan-3 for successful fiber formation. J. Cell Sci. 2004, 117 Pt 1, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Winkler, S.; Stahl, R.C.; Carey, D.J.; Bansal, R. Syndecan-3 and perlecan are differentially expressed by progenitors and mature oligodendrocytes and accumulate in the extracellular matrix. J. Neurosci. Res. 2002, 69, 477–487. [Google Scholar] [CrossRef]
- Inatani, M.; Haruta, M.; Honjo, M.; Oohira, A.; Kido, N.; Takahashi, M.; Honda, Y.; Tanihara, H. Upregulated expression of N-syndecan, a transmembrane heparan sulfate proteoglycan, in differentiated neural stem cells. Brain Res. 2001, 920, 217–221. [Google Scholar] [CrossRef]
- Huang, R.-L.; Chen, H.-J.; Chen, L.-Y.; Chao, T.-K.; Lin, W.-Y.; Liew, P.-L.; Su, P.-H.; Weng, Y.-C.; Wang, Y.-C.; Liao, C.-C.; et al. Epigenetic loss of heparan sulfate 3-O-sulfation sensitizes ovarian carcinoma to oncogenic signals and predicts prognosis. Int. J. Cancer 2018, 143, 1943–1953. [Google Scholar] [CrossRef]
- Kumar, A.V.; Gassar, E.S.; Spillmann, D.; Stock, C.; Sen, Y.-P.; Zhang, T.; Van Kuppevelt, T.H.; Hülsewig, C.; Koszlowski, E.O.; Pavao, M.S.; et al. HS3ST2 modulates breast cancer cell invasiveness via MAP kinase- and Tcf4 (Tcf7l2)-dependent regulation of protease and cadherin expression. Int. J. Cancer 2014, 135, 2579–2592. [Google Scholar] [CrossRef]
- Kamimura, K.; Rhodes, J.M.; Ueda, R.; McNeely, M.; Shukla, D.; Kimata, K.; Spear, P.G.; Shworak, N.W.; Nakato, H. Regulation of Notch signaling by Drosophila heparan sulfate 3-O sulfotransferase. J. Cell Biol. 2004, 166, 1069–1079. [Google Scholar] [CrossRef] [Green Version]
- Steg, A.D.; Bevis, K.S.; Katre, A.A.; Ziebarth, A.; Dobbin, Z.C.; Alvarez, R.D.; Zhang, K.; Conner, M.; Landen, C.N. Stem Cell Pathways Contribute to Clinical Chemoresistance in Ovarian Cancer. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2011, 18, 869–881. [Google Scholar] [CrossRef] [Green Version]
- Prasetyanti, P.R.; Medema, J.P. Intra-tumor heterogeneity from a cancer stem cell perspective. Mol. Cancer 2017, 16, 41. [Google Scholar] [CrossRef] [Green Version]
- Ottevanger, P.B. Ovarian cancer stem cells more questions than answers. Semin. Cancer Biol. 2017, 44, 67–71. [Google Scholar] [CrossRef]
- Yang-Hartwich, Y.; Soteras, M.G.; Lin, Z.P.; Holmberg, J.; Sumi, N.; Craveiro, V.; Liang, M.; Romanoff, E.; Bingham, J.; Garofalo, F.; et al. p53 protein aggregation promotes platinum resistance in ovarian cancer. Oncogene 2015, 34, 3605–3616. [Google Scholar] [CrossRef]
- Patch, A.-M.; Christie, E.L.; Etemadmoghadam, D.; Garsed, D.W.; George, J.; Fereday, S.; Nones, K.; Cowin, P.; Alsop, K.; Bailey, P.J.; et al. Whole–genome characterization of chemoresistant ovarian cancer. Nature 2015, 521, 489–494. [Google Scholar] [CrossRef]
- Barakat, B.M.; Wang, Q.-E.; Han, C.; Milum, K.; Yin, D.-T.; Zhao, Q.; Wani, G.; Arafa, E.-S.A.; El-Mahdy, M.A.; Wani, A.A. Overexpression of DDB2 enhances the sensitivity of human ovarian cancer cells to cisplatin by augmenting cellular apoptosis. Int. J. Cancer 2010, 127, 977–988. [Google Scholar] [CrossRef]
- Marchini, S.; Fruscio, R.; Clivio, L.; Beltrame, L.; Porcu, L.; Nerini, I.F.; Cavalieri, D.; Chiorino, G.; Cattoretti, G.; Mangioni, C.; et al. Resistance to platinum-based chemotherapy is associated with epithelial to mesenchymal transition in epithelial ovarian cancer. Eur. J. Cancer 2013, 49, 520–530. [Google Scholar] [CrossRef]
- Chebouti, I.; Kasimir-Bauer, S.; Buderath, P.; Wimberger, P.; Hauch, S.; Kimmig, R.; Kuhlmann, J.D. EMT-like circulating tumor cells in ovarian cancer patients are enriched by platinum-based chemotherapy. Oncotarget 2017, 8, 48820–48831. [Google Scholar] [CrossRef] [Green Version]
- Du, B.; Shim, J.S. Targeting Epithelial–Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules 2016, 21, 965. [Google Scholar] [CrossRef] [Green Version]
- Nieto, M.A. The snail superfamily of zinc-finger transcription factors. Nat. Rev. Mol. Cell Biol. 2002, 3, 155–166. [Google Scholar] [CrossRef]
- Abedini, A.; Sayed, C.; Carter, L.E.; Boerboom, D.; Vanderhyden, B.C. Non-canonical WNT5a regulates Epithelial-to-Mesenchymal Transition in the mouse ovarian surface epithelium. Sci. Rep. 2020, 10, 9695. [Google Scholar] [CrossRef]
- Zhou, W.; Mei, J.; Gu, D.; Xu, J.; Wang, R.; Wang, H.; Liu, C. Wnt5a: A promising therapeutic target in ovarian cancer. Pathol. Res. Pract. 2021, 219, 153348. [Google Scholar] [CrossRef]
- Kotrbová, A.; Ovesná, P.; Gybel’, T.; Radaszkiewicz, T.; Bednaříková, M.; Hausnerová, J.; Jandáková, E.; Minář, L.; Crha, I.; Weinberger, V.; et al. WNT signaling inducing activity in ascites predicts poor outcome in ovarian cancer. Theranostics 2020, 10, 537–552. [Google Scholar] [CrossRef]
- Burston, H.E.; Kent, O.A.; Communal, L.; Udaskin, M.L.; Sun, R.X.; Brown, K.R.; Jung, E.; Francis, K.E.; La Rose, J.; Lowitz, J.K.; et al. Inhibition of relaxin autocrine signaling confers therapeutic vulnerability in ovarian cancer. J. Clin. Investig. 2021, 131, e142677. [Google Scholar] [CrossRef]
- Nikolova, V.; Koo, C.Y.; Ibrahim, S.A.; Wang, Z.; Spillmann, D.; Dreier, R.; Kelsch, R.; Fischgräbe, J.; Smollich, M.; Rossi, L.H.; et al. Differential roles for membrane-bound and soluble syndecan-1 (CD138) in breast cancer progression. Carcinogenesis 2009, 30, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Kato, M.; Saunders, S.; Nguyen, H.; Bernfield, M. Loss of cell surface syndecan-1 causes epithelia to transform into anchorage-independent mesenchyme-like cells. Mol. Biol. Cell 1995, 6, 559–576. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Drug | IC50 [nM] | Combination Index (CI) [nM] | ||||
|---|---|---|---|---|---|---|---|
| Control | Sequence 1 | Sequence 2 | Control | Sequence 1 | Sequence 2 | ||
| SKOV3 | Taxol | 2.909 | 1.679 | 0.4900 | - | - | - |
| Cisplatin | 2.270 | 2.887 | 1.355 | - | - | - | |
| Taxol/Cisplatin | - | - | - | 0.930 | 18.198 | <0.001 | |
| CAOV3 | Taxol | 1.952 | 1.947 | 1.985 | - | - | - |
| Cisplatin | 1.222 | 1.304 | 1.360 | - | - | ||
| Taxol/Cisplatin | - | - | - | 13.013 | 10.649 | 12.774 | |
| mRNA | Sequence (Forward) | Sequence (Reverse) |
|---|---|---|
| ACTB | TCAAGATCATTGCTCCTCCTGAG | ACATCTGCTGGAAGGTGGACA |
| CDH1 | CAAAGCCCAGAATCCCCAAG | CACACCTGGAATTGGGCAAA |
| Gli1 | TTCCTACCAGAGTCCCAAGT | CCCTATGTGAAGCCCTATTT |
| Gli2 | GTCAGAGCCATCAAGACCGAGA | GCATCTCCACGCCACTGTCATT |
| Gli3 | TCAGCAAGTGGCTCCTATGGTC | GCTCTGTTGTCGGCTTAGGATC |
| Hes1 | GTGAAGCACCTCCGGAAC | CGTTCATGCACTCGCTGA |
| Hey2 | TTGCCCATGCCTAAACTAGTGC | TCTCACGTGCTTGATTTCAGCATA |
| Msi1 | AGCTTCCCTCTCCCTCATTCC | GGAATTCCAGGGTCCTGAGC |
| Msi2 | CCTAGTATGCTTGCCTCACAAACG | CGTCATCAGGGAGAAGCACAG |
| Notch1 | GGTGAGACCTGCCTGAATG | GTTGGGGTCCTGGCATC |
| PTCH1 | GGTGGCACAGTCAAGAACA | ACCAAGAGCGAGAAATGG |
| Robo1 | GCATCGCTGGAAGTAGCCATACT | CATGAAATGGTGGGCTCAGGAT |
| SDC3 | TGCCTCAGAAGAGTATCCTG | CTTGTCAGGCTTCTGGTATG |
| SFRP1 | GATGCAGGAGGCTCAGGTGAT | GCTGGCAACAGGTCAGAACG |
| SHH | GAAAGCAGAGAACTCGGTGG | GGAAAGTGAGGAAGTCGCTG |
| Snail | CGAGCCCAGGCAGCTATTTC | CCCGACAAGTGACAGCCATT |
| Src | ACCACCTTTGTGGCCCTCTATG | GCCACCAGTCTCCCTCTGTGTT |
| TCF7L1 | AAGGTGCCTGCCACTTCCTC | CCTGCCACTCTGGGATTGTG |
| TCF7L2 | AAACCAGCTGCCGCTTTTATG | GCAACATCAACATGCCTAGGTTTT |
| Twist | GCGGCCAGGTACATCGACTT | TGCAGCTTGCCATCTTGGAG |
| Vimentin | TCAGCATCACGATGACCTTGAA | CTGCAGAAAGGCACTTGAAAGC |
| Wnt 3A | AGGTCCCACAGCCCTGAGAT | TCCAGGAAAGCGGACCATTT |
| Wnt 5A | TCGTTAGCAGCATCAGTCCACA | GACCTGTGCCTTCGTGCCTA |
| ZEB1 | CAGCCCTGCAGTCCAAGAAC | TTGTCTTTCATCCTGATTTCCATTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hillemeyer, L.; Espinoza-Sanchez, N.A.; Greve, B.; Hassan, N.; Chelariu-Raicu, A.; Kiesel, L.; Götte, M. The Cell Surface Heparan Sulfate Proteoglycan Syndecan-3 Promotes Ovarian Cancer Pathogenesis. Int. J. Mol. Sci. 2022, 23, 5793. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105793
Hillemeyer L, Espinoza-Sanchez NA, Greve B, Hassan N, Chelariu-Raicu A, Kiesel L, Götte M. The Cell Surface Heparan Sulfate Proteoglycan Syndecan-3 Promotes Ovarian Cancer Pathogenesis. International Journal of Molecular Sciences. 2022; 23(10):5793. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105793
Chicago/Turabian StyleHillemeyer, Lara, Nancy Adriana Espinoza-Sanchez, Burkhard Greve, Nourhan Hassan, Anca Chelariu-Raicu, Ludwig Kiesel, and Martin Götte. 2022. "The Cell Surface Heparan Sulfate Proteoglycan Syndecan-3 Promotes Ovarian Cancer Pathogenesis" International Journal of Molecular Sciences 23, no. 10: 5793. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105793