
The Long-Term Culture of Human Fibroblasts Reveals a Spectroscopic Signature of Senescence

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
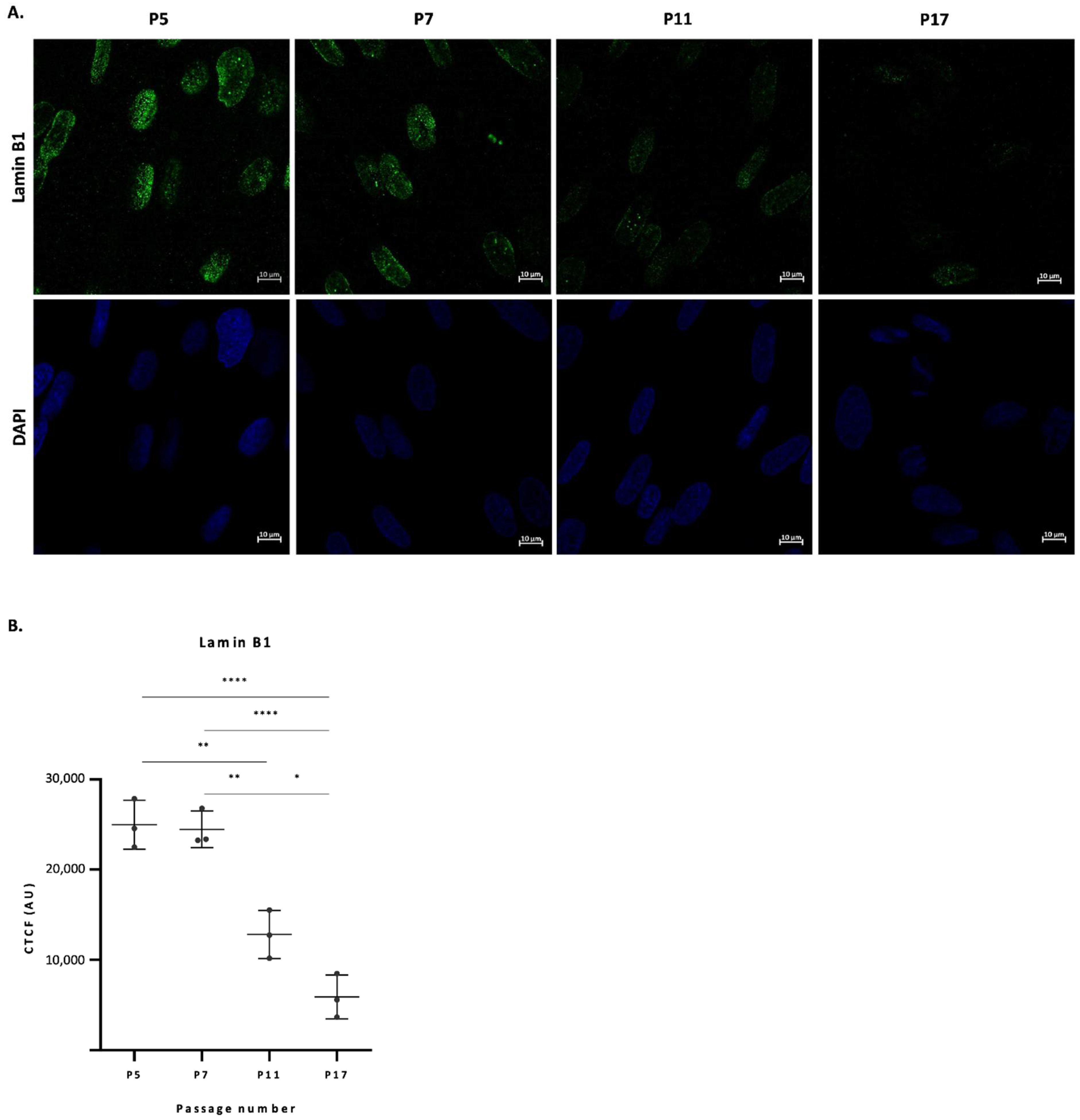
2.1. Lamin B1 Levels in Fibroblast Nuclei Drop from Passage 5 to Passage 17
2.2. FTIR Spectra Overview
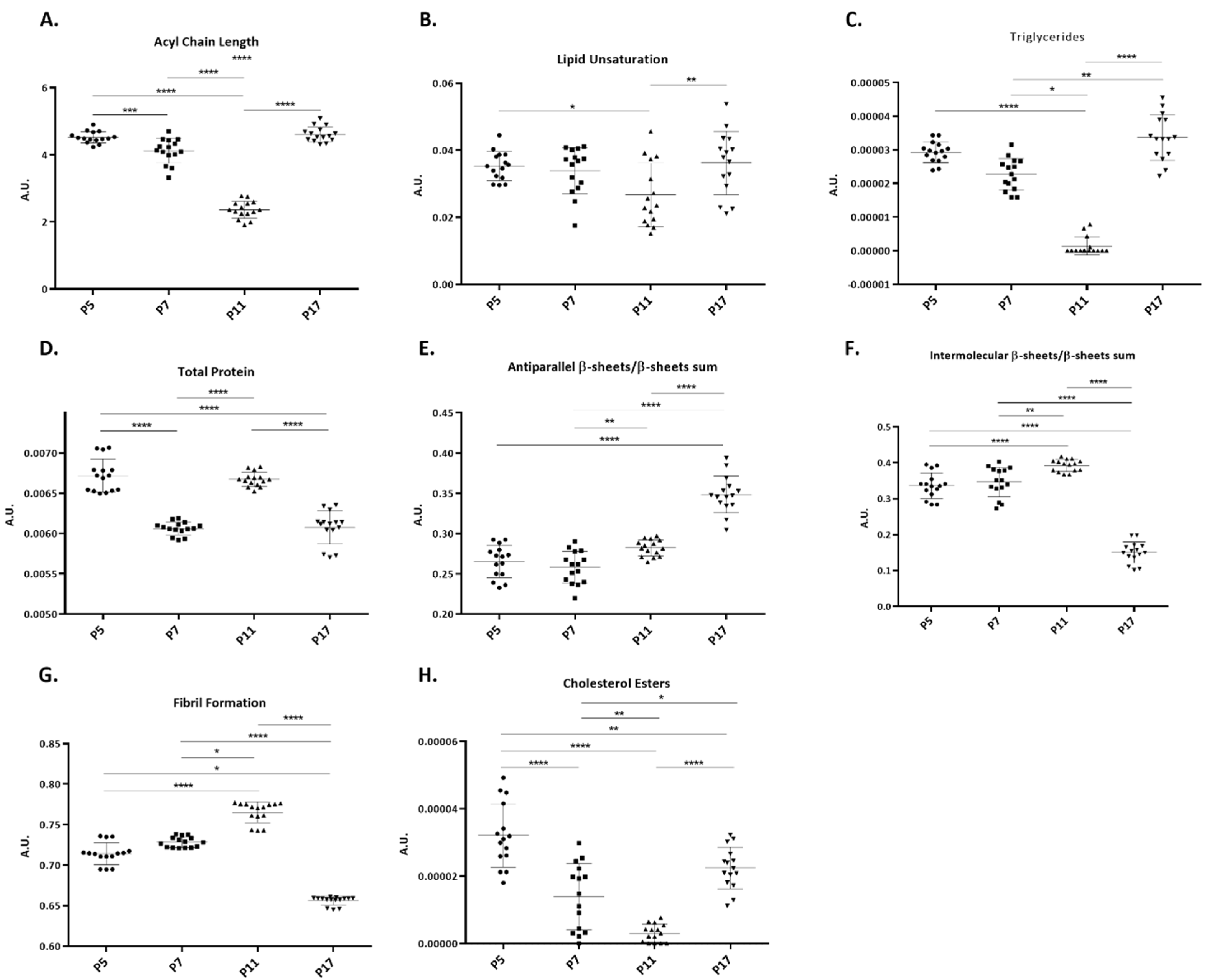
2.3. Intensity of FTIR Spectral Bands
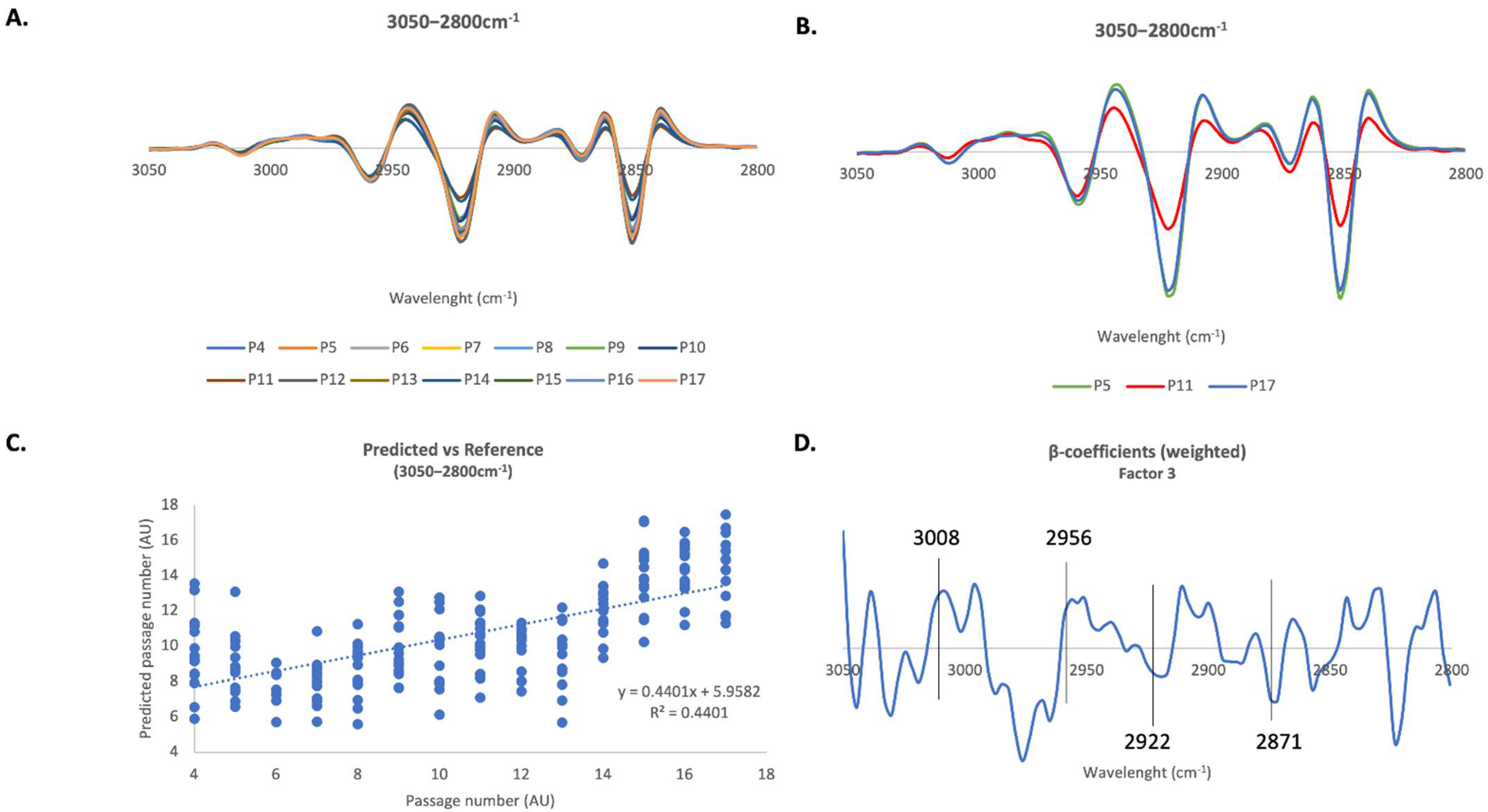
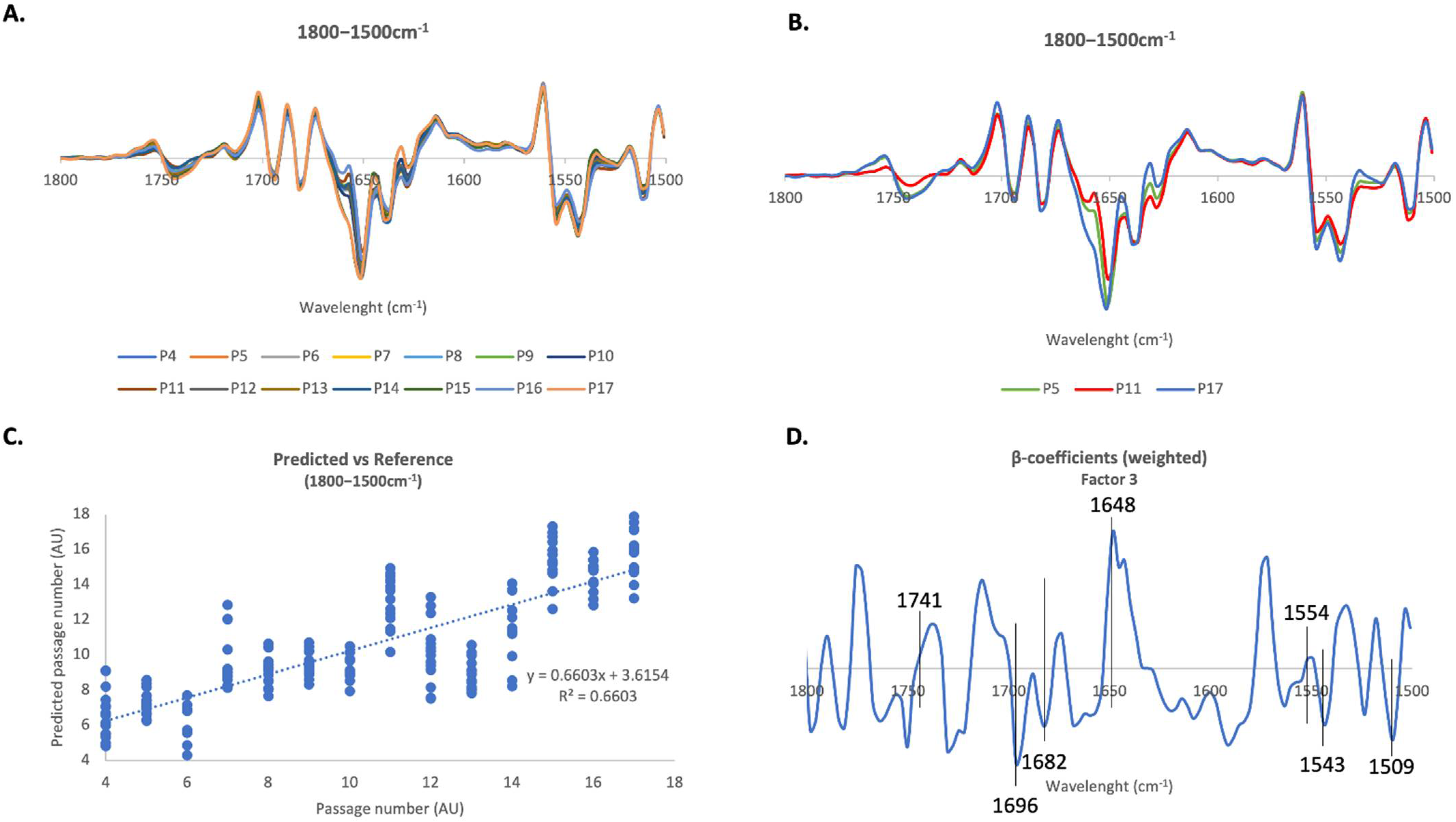
2.4. PLS-R Multivariate Analysis
2.4.1. 3050–2800 cm−1 Region
2.4.2. 1800–1500 cm−1 Region
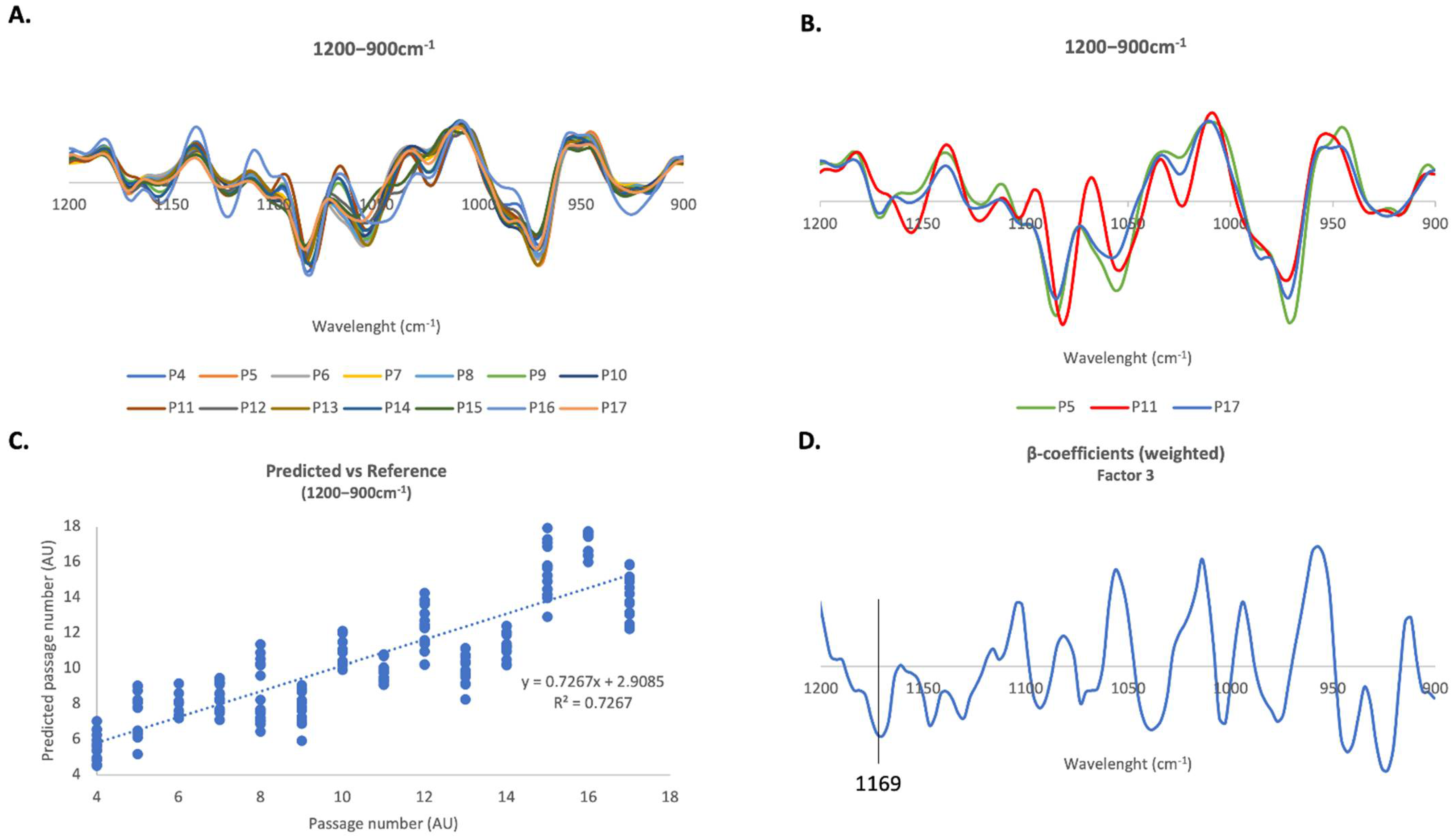
2.4.3. 1200–900 cm−1 Region
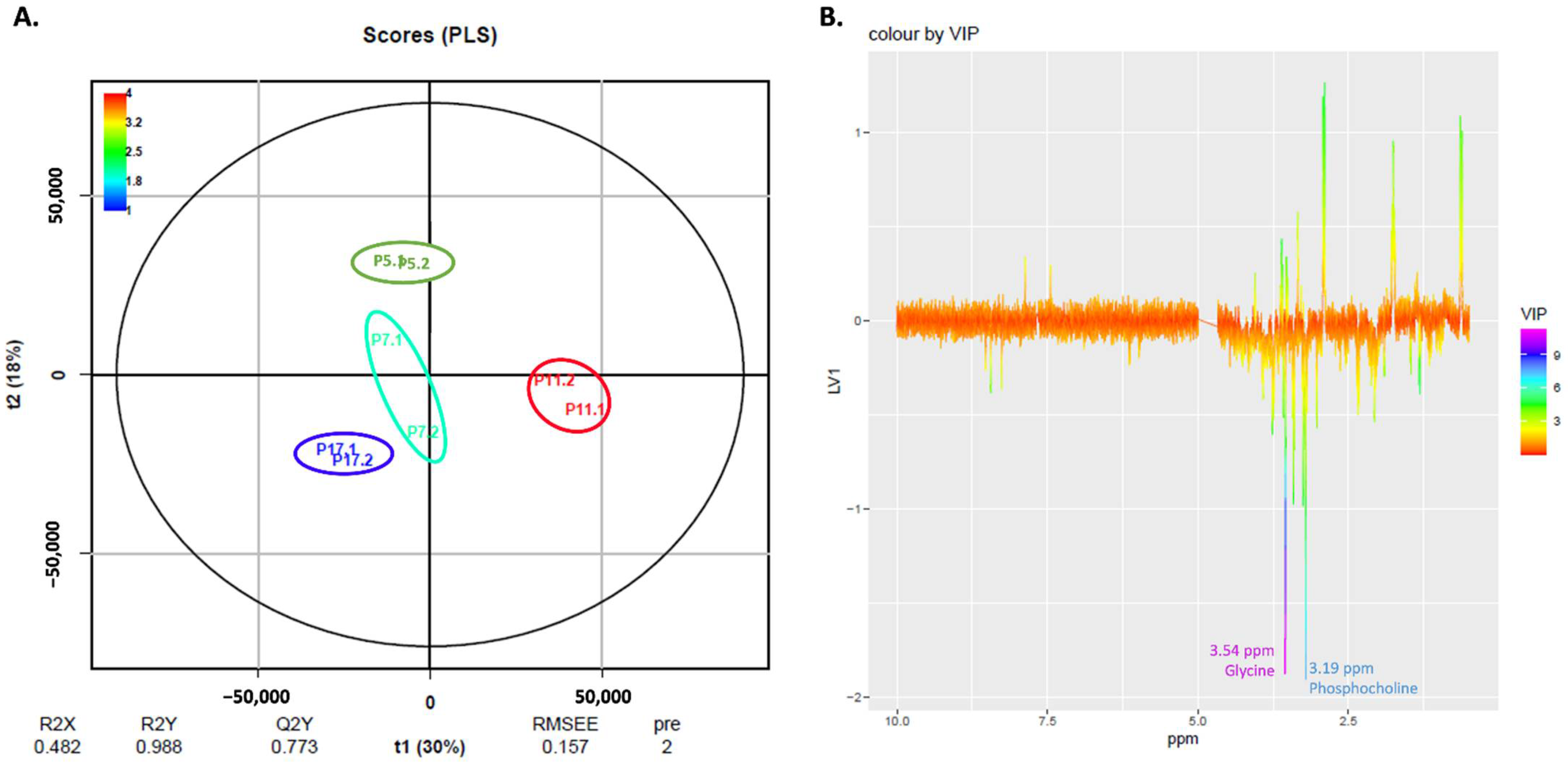
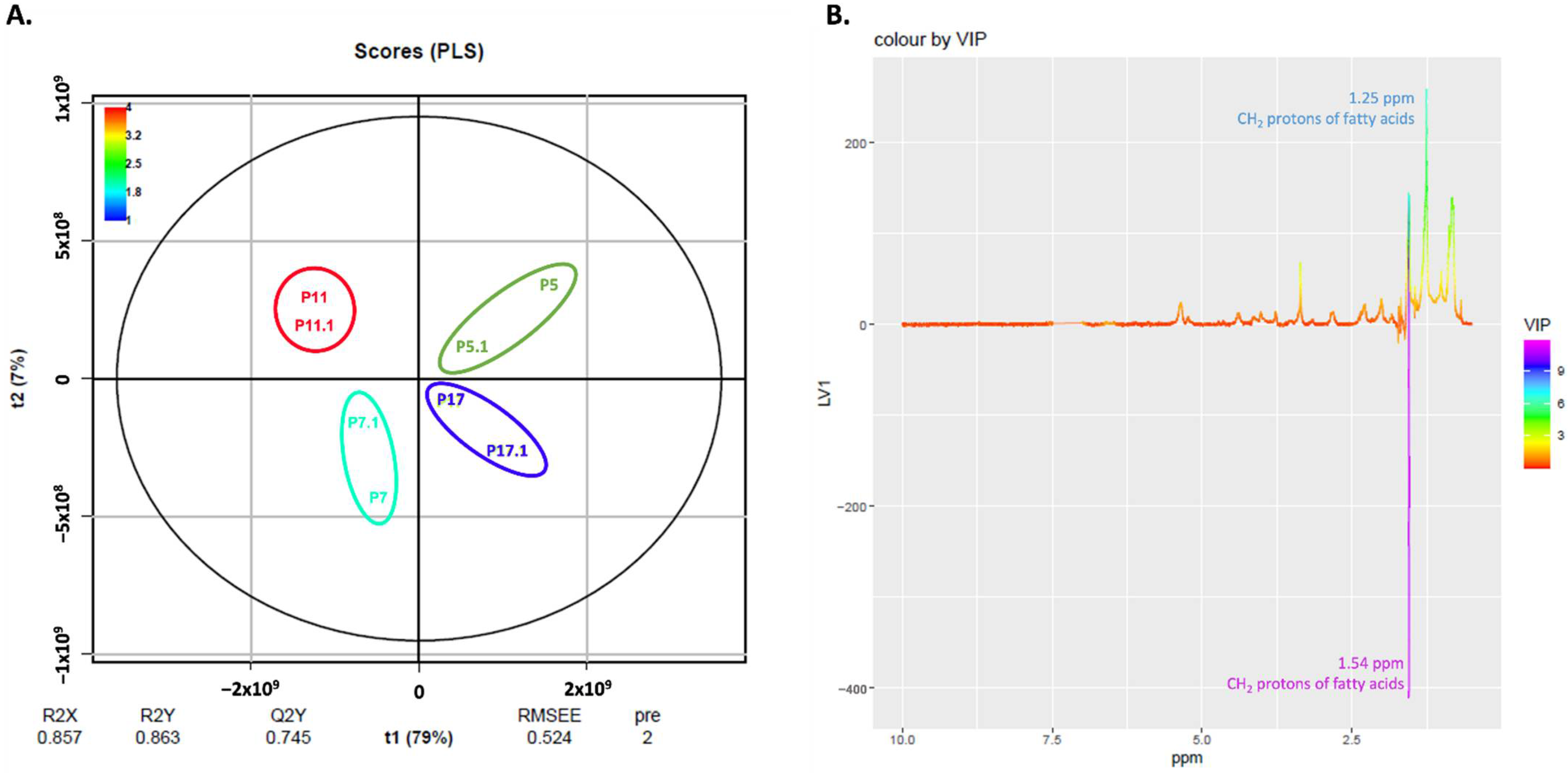
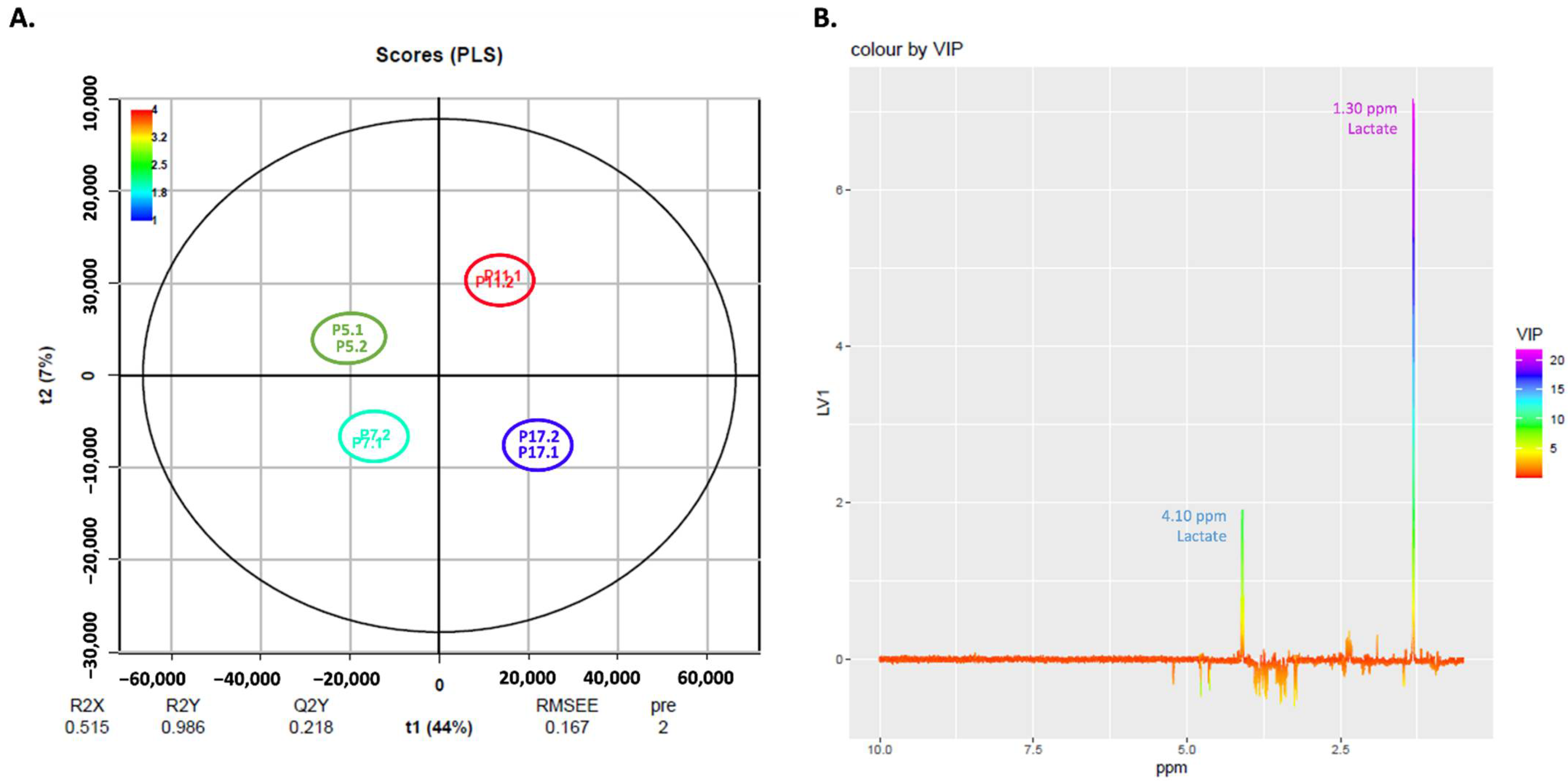
2.5. NMR Exploratory Analysis of Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture Procedures
4.2. Immunocytochemistry (ICC)
4.3. Confocal Microscopy
4.4. Fourier-Transform InfraRed (FTIR) Spectroscopy
4.4.1. Cell Pellet Preparation
4.4.2. FTIR Spectra Measurements and Spectra Preprocessing
4.4.3. PLS-R Multivariate Analysis
4.4.4. Analysis of Peak Intensities
4.5. NMR Spectroscopy
4.5.1. Quenching and Extraction of the Cell Extracts: Endo- and Exometabolome
4.5.2. NMR Spectral Processing
4.5.3. Multivariate Analysis
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- López-Otín, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The Hallmarks of Aging. Cell 2016, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powers, E.T.; Morimoto, R.I.; Dillin, A.; Kelly, J.W.; Balch, W.E. Biological and Chemical Approaches to Diseases of Proteostasis Deficiency. Annu. Rev. Biochem. 2009, 78, 959–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labbadia, J.; Morimoto, R.I. The Biology of Proteostasis in Aging and Disease. Annu. Rev. Biochem. 2015, 84, 435–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magalhaes, S.; Goodfellow, B.J.; Nunes, A. Aging and Proteins: What Does Proteostasis Have to Do with Age? Curr. Mol. Med. 2018, 18, 178–189. [Google Scholar] [CrossRef]
- Hayflick, L.; Moorhead, P.S. The serial cultivation of human diploid cell strains. Exp. Cell Res. 1961, 25, 585–621. [Google Scholar] [CrossRef]
- Calcinotto, A.; Kohli, J.; Zagato, E.; Pellegrini, L.; Demaria, M.; Alimonti, A. Cellular Senescence: Aging, Cancer, and Injury. Physiol. Rev. 2019, 99, 1047–1078. [Google Scholar] [CrossRef]
- Yu, G.; Hyun, S. Proteostasis-associated aging: Lessons from a Drosophila model. Genes Genom. 2021, 43, 1–9. [Google Scholar] [CrossRef]
- Guo, S.; Wang, X.; Kang, L. Special Significance of Non-Drosophila Insects in Aging. Front. Cell Dev. Biol. 2020, 8, 576571. [Google Scholar] [CrossRef]
- Walther, D.M.; Kasturi, P.; Zheng, M.; Pinkert, S.; Vecchi, G.; Ciryam, P.; Morimoto, R.I.; Dobson, C.M.; Vendruscolo, M.; Mann, M.; et al. Widespread proteome remodeling and aggregation in aging C. elegans. Cell 2015, 161, 919–932. [Google Scholar] [CrossRef] [Green Version]
- Magalhães, S.; Almeida, I.; Martins, F.; Camões, F.; Soares, A.R.; Goodfellow, B.J.; Rebelo, S.; Nunes, A. FTIR Spectroscopy as a Tool to Study Age-Related Changes in Cardiac and Skeletal Muscle of Female C57BL/6J Mice. Molecules 2021, 26, 6410. [Google Scholar] [CrossRef]
- Rocha, A.; Magalhães, S.; Nunes, A. Cell Culture Studies: A Promising Approach to the Metabolomic Study of Human Aging. Curr. Metabolomics Syst. Biol. 2021, 8, 1–26. [Google Scholar] [CrossRef]
- Schneider, E.L. Aging and Cultured Human Skin in Fibroblasts. J. Invest Dermatol. 1979, 73, 15–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stacey, G. Primary Cell Cultures and Immortal Cell Lines. In eLS; Wiley: Hoboken, NJ, USA, 2006. [Google Scholar]
- Phipps, S.M.O.; Berletch, J.B.; Andrews, L.G.; Tollefsbol, T.O. Aging Cell Culture. Methods Mol. Biol. 2007, 371, 9–19. [Google Scholar] [PubMed]
- Schneider, E.L.; Mitsui, Y. The relationship between in vitro cellular aging and in vivo human age. Proc. Natl. Acad. Sci. USA 1976, 73, 3584–3588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čuperlović-Culf, M.; Barnett, D.A.; Culf, A.S.; Chute, I. Cell culture metabolomics: Applications and future directions. Drug Discov. Today 2010, 15, 610–621. [Google Scholar] [CrossRef]
- Eberhardt, K.; Beleites, C.; Marthandan, S.; Matthäus, C.; Diekmann, S.; Popp, J. Raman and Infrared Spectroscopy Distinguishing Replicative Senescent from Proliferating Primary Human Fibroblast Cells by Detecting Spectral Differences Mainly Due to Biomolecular Alterations. Anal. Chem. 2017, 89, 2937–2947. [Google Scholar] [CrossRef]
- Dreesen, O.; Ong, P.F.; Chojnowski, A.; Colman, A. The contrasting roles of lamin B1 in cellular aging and human disease. Nucleus 2013, 4, 283–290. [Google Scholar] [CrossRef] [Green Version]
- Viegas, D.; Pereira, C.D.; Martins, F.; Mateus, T.; da Cruz e Silva, O.A.B.; Herdeiro, M.T.; Rebelo, S. Nuclear Envelope Alterations in Myotonic Dystrophy Type 1 Patient-Derived Fibroblasts. Int. J. Mol. Sci. 2022, 23, 522. [Google Scholar] [CrossRef]
- Inoue, K. CRISPR-activated patient fibroblasts for modeling of familial Alzheimer’s disease. Neurosci. Res. 2021, 172, 7–12. [Google Scholar] [CrossRef]
- Matloka, M.; Klein, A.F.; Rau, F.; Furling, D. Cells of Matter—In Vitro Models for Myotonic Dystrophy. Front. Neurol. 2018, 9, 361. [Google Scholar] [CrossRef]
- Rocha, A.; Magalhães, S.; Nunes, A. Study Aging by Fibroblasts Metabolome. Curr. Mol. Med. 2021, 21, 300–317. [Google Scholar] [CrossRef] [PubMed]
- Tigges, J.; Krutmann, J.; Fritsche, E.; Haendeler, J.; Schaal, H.; Fischer, J.W.; Kalfalah, F.; Reinke, H.; Reifenberger, G.; Stühler, K.; et al. The hallmarks of fibroblast ageing. Mech. Ageing Dev. 2014, 138, 26–44. [Google Scholar] [CrossRef] [PubMed]
- Porter, N.M.; Thibault, O.; Thibault, V.; Chen, K.-C.; Landfield, P.W. Calcium Channel Density and Hippocampal Cell Death with Age in Long-Term Culture. J. Neurosci. 1997, 17, 5629–5639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benazzoug, Y.; Borchiellini, C.; Labat-Robert, J.; Robert, L.; Kern, P. Effect of high-glucose concentrations on the expression of collagens and fibronectin by fibroblasts in culture. Exp. Gerontol. 1998, 33, 445–455. [Google Scholar] [CrossRef]
- Raffetto, J.D.; Mendez, M.V.; Phillips, T.J.; Park, H.-Y.; Menzoian, J.O. The effect of passage number on fibroblast cellular senescence in patients with chronic venous insufficiency with and without ulcer. Am. J. Surg. 1999, 178, 107–112. [Google Scholar] [CrossRef]
- Yang, Y.-H.K.; Ogando, C.R.; Wang See, C.; Chang, T.-Y.; Barabino, G.A. Changes in phenotype and differentiation potential of human mesenchymal stem cells aging in vitro. Stem Cell Res. Ther. 2018, 9, 131. [Google Scholar] [CrossRef] [Green Version]
- Atkuru, S.; Muniraj, G.; Sudhaharan, T.; Chiam, K.; Wright, G.D.; Sriram, G. Cellular ageing of oral fibroblasts differentially modulates extracellular matrix organization. J. Periodontal Res. 2021, 56, 108–120. [Google Scholar] [CrossRef]
- Srisongkram, T.; Weerapreeyakul, N.; Thumanu, K. Evaluation of Melanoma (SK-MEL-2) Cell Growth between Three-Dimensional (3D) and Two-Dimensional (2D) Cell Cultures with Fourier Transform Infrared (FTIR) Microspectroscopy. Int. J. Mol. Sci. 2020, 21, 4141. [Google Scholar] [CrossRef]
- Wehbe, K.; Vezzalini, M.; Cinque, G. Detection of mycoplasma in contaminated mammalian cell culture using FTIR microspectroscopy. Anal. Bioanal. Chem. 2018, 410, 3003–3016. [Google Scholar] [CrossRef] [Green Version]
- Poonprasartporn, A.; Chan, K.L.A. Live-cell ATR-FTIR spectroscopy as a novel bioanalytical tool for cell glucose metabolism research. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 119024. [Google Scholar] [CrossRef]
- Romanò, S.; Di Giacinto, F.; Primiano, A.; Mazzini, A.; Panzetta, C.; Papi, M.; Di Gaspare, A.; Ortolani, M.; Gervasoni, J.; De Spirito, M.; et al. Fourier Transform Infrared Spectroscopy as a useful tool for the automated classification of cancer cell-derived exosomes obtained under different culture conditions. Anal. Chim. Acta 2020, 1140, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Montoliu, I.; Scherer, M.; Beguelin, F.; Dasilva, L.; Mari, D.; Martin, F.P.J.; Capri, M.; Bucci, L.; Ostan, R.; Monti, D.; et al. Serum profiling of healthy aging identifies phospho - and sphingolipid species as markers of human longevity. Aging 2014, 6, 9–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, I.; Magalhães, S.; Nunes, A. Lipids: Biomarkers of healthy aging. Biogerontology 2021, 22, 273–295. [Google Scholar] [CrossRef] [PubMed]
- Jobson, R.W.; Nabholz, B.; Galtier, N. An Evolutionary Genome Scan for Longevity-Related Natural Selection in Mammals. Mol. Biol. Evol. 2010, 27, 840–847. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Covarrubias, V.; Beekman, M.; Uh, H.; Dane, A.; Troost, J.; Paliukhovich, I.; Kloet, F.M.; Houwing-Duistermaat, J.; Vreeken, R.J.; Hankemeier, T.; et al. Lipidomics of familial longevity. Aging Cell 2013, 12, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Hulbert, A.J.; Pamplona, R.; Buffenstein, R.; Buttemer, W.A. Life and death: Metabolic rate, membrane composition, and life span of animals. Physiol. Rev. 2007, 87, 1175–1213. [Google Scholar] [CrossRef]
- Jové, M.; Naudí, A.; Gambini, J.; Borras, C.; Cabré, R.; Portero-Otín, M.; Viña, J.; Pamplona, R. A Stress-Resistant Lipidomic Signature Confers Extreme Longevity to Humans. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2017, 72, 30–37. [Google Scholar] [CrossRef]
- Pradas, I.; Jové, M.; Huynh, K.; Puig, J.; Ingles, M.; Borras, C.; Viña, J.; Meikle, P.; Pamplona, R. Exceptional human longevity is associated with a specific plasma phenotype of ether lipids. Redox Biol. 2019, 21, 101127. [Google Scholar] [CrossRef]
- Gey, C.; Seeger, K. Metabolic changes during cellular senescence investigated by proton NMR-spectroscopy. Mech. Ageing Dev. 2013, 134, 130–138. [Google Scholar] [CrossRef]
- Dong, A.; Huang, P.; Caughey, W.S. Protein Secondary Structures in Water from Second-Derivative Amide I Infrared Spectra. Biochemistry 1990, 29, 3303–3308. [Google Scholar] [CrossRef]
- Kaushik, S.; Cuervo, A.M. Proteostasis and aging. Nat. Med. 2015, 21, 1406–1415. [Google Scholar] [CrossRef] [PubMed]
- Roeters, S.J.; Iyer, A.; Pletikapić, G.; Kogan, V.; Subramaniam, V.; Woutersen, S. Evidence for Intramolecular Antiparallel Beta-Sheet Structure in Alpha-Synuclein Fibrils from a Combination of Two-Dimensional Infrared Spectroscopy and Atomic Force Microscopy. Sci. Rep. 2017, 7, 41051. [Google Scholar] [CrossRef] [PubMed]
- Chiti, F.; Dobson, C.M. Protein Misfolding, Amyloid Formation, and Human Disease: A Summary of Progress Over the Last Decade. Annu. Rev. Biochem. 2017, 86, 27–68. [Google Scholar] [CrossRef] [PubMed]
- Ho, P.W.-L.; Leung, C.-T.; Liu, H.; Pang, S.Y.-Y.; Lam, C.S.-C.; Xian, J.; Li, L.; Kung, M.H.-W.; Ramsden, D.B.; Ho, S.-L. Age-dependent accumulation of oligomeric SNCA/α-synuclein from impaired degradation in mutant LRRK2 knockin mouse model of Parkinson disease: Role for therapeutic activation of chaperone-mediated autophagy (CMA). Autophagy 2020, 16, 347–370. [Google Scholar] [CrossRef] [PubMed]
- Walton, C.C.; Begelman, D.; Nguyen, W.; Andersen, J.K. Senescence as an Amyloid Cascade: The Amyloid Senescence Hypothesis. Front. Cell. Neurosci. 2020, 14, 129. [Google Scholar] [CrossRef]
- Wang, J.; Zheng, B.; Yang, S.; Wang, F.; Wang, Z.; Wang, J. The protective effects of Agomelatine against Aβ1-42 oligomers-induced cellular senescence mediated by SIRT6 and Agomelatine’s potential in AD treatment. Hum. Cell 2021, 34, 1734–1743. [Google Scholar] [CrossRef]
- Birch, J.; Gil, J. Senescence and the SASP: Many therapeutic avenues. Genes Dev. 2020, 34, 1565–1576. [Google Scholar] [CrossRef]
- Gasek, N.S.; Kuchel, G.A.; Kirkland, J.L.; Xu, M. Strategies for targeting senescent cells in human disease. Nat. Aging 2021, 1, 870–879. [Google Scholar] [CrossRef]
- Certo, M.; Tsai, C.-H.; Pucino, V.; Ho, P.-C.; Mauro, C. Lactate modulation of immune responses in inflammatory versus tumour microenvironments. Nat. Rev. Immunol. 2021, 21, 151–161. [Google Scholar] [CrossRef]
- Proia, P.; Di Liegro, C.; Schiera, G.; Fricano, A.; Di Liegro, I. Lactate as a Metabolite and a Regulator in the Central Nervous System. Int. J. Mol. Sci. 2016, 17, 1450. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Covarrubias, V. Lipidomics in longevity and healthy aging. Biogerontology 2013, 14, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; DeZonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [PubMed]
- Rasband, W.S. ImageJ. Version 1.8.0; NIH; Bethesda, Rockville, MD, USA; 1997. Available online: https://imagej.nih.gov/ij/ (accessed on 9 March 2022).
- Pereira, C.D.; Martins, F.; Santos, M.; Müeller, T.; da Cruz e Silva, O.A.B.; Rebelo, S. Nuclear Accumulation of LAP1:TRF2 Complex during DNA Damage Response Uncovers a Novel Role for LAP1. Cells 2020, 9, 1804. [Google Scholar] [CrossRef] [PubMed]
- Mateus, T.; Almeida, I.; Costa, A.; Viegas, D.; Magalhães, S.; Martins, F.; Herdeiro, M.T.; da Cruz e Silva, O.A.B.; Fraga, C.; Alves, I.; et al. Fourier-Transform Infrared Spectroscopy as a Discriminatory Tool for Myotonic Dystrophy Type 1 Metabolism: A Pilot Study. Int. J. Environ. Res. Public Health 2021, 18, 3800. [Google Scholar] [CrossRef]
- Crowley, L.C.; Marfell, B.J.; Christensen, M.E.; Waterhouse, N.J. Measuring Cell Death by Trypan Blue Uptake and Light Microscopy. Cold Spring Harb. Protoc. 2016, 2016, pdb.prot087155. [Google Scholar] [CrossRef]
- OPUS. Version 7.0; Bruker Optics LDA; Billerica, MA, USA. Available online: https://www.bruker.com/en/products-and-solutions/infrared-and-raman/opus-spectroscopy-software.html (accessed on 9 March 2022).
- The Unscrambler X. Version 10.5.1; Camo Analytics; Oslo; Norway. Available online: https://the-unscrambler-x.software.informer.com/10.5/ (accessed on 9 March 2022).
- Magalhães, S.; Goodfellow, B.J.; Nunes, A. FTIR spectroscopy in biomedical research: How to get the most out of its potential. Appl. Spectrosc. Rev. 2021, 56, 1–39. [Google Scholar] [CrossRef]
- Palomino-Schätzlein, M.; Molina-Navarro, M.M.; Tormos-Pérez, M.; Rodríguez-Navarro, S.; Pineda-Lucena, A. Optimised protocols for the metabolic profiling of S. cerevisiae by 1H-NMR and HRMAS spectroscopy. Anal. Bioanal. Chem. 2013, 405, 8431–8441. [Google Scholar] [CrossRef]
- iNMR. Version 6.4.5; Nucleomatica; Molfetta, Italy. 2005. Available online: https://www.inmr.net/downloads/ (accessed on 9 March 2022).
- RStudio. Version 2022.02.2-485; RStudio Inc.; Boston MA USA. 2009. Available online: https://www.rstudio.com/products/rstudio/release-notes/ (accessed on 9 March 2022).
- Beirnaert, C.; Meysman, P.; Vu, T.N.; Hermans, N.; Apers, S.; Pieters, L.; Covaci, A.; Laukens, K. speaq 2.0: A complete workflow for high-throughput 1D NMR spectra processing and quantification. PLoS Comput. Biol. 2018, 14, e1006018. [Google Scholar] [CrossRef] [Green Version]
- Thévenot, E.A.; Roux, A.; Xu, Y.; Ezan, E.; Junot, C. Analysis of the Human Adult Urinary Metabolome Variations with Age, Body Mass Index, and Gender by Implementing a Comprehensive Workflow for Univariate and OPLS Statistical Analyses. J. Proteome Res. 2015, 14, 3322–3335. [Google Scholar] [CrossRef]
- GraphPad Prism. Version 6.0; GraphPad Software; San Diego, CA, USA. 1989. Available online: https://www.graphpad.com/support/prism-6-updates/ (accessed on 9 March 2022).










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young Fibroblasts | Old Fibroblasts |
|---|---|
| ↑ CH2 groups of lipids | ↑ CH3 groups of lipids (except for cells at passage 17) |
| ↑ CH groups of lipids | |
| ↓ triglycerides (except for cells at passage 17) | |
| ↓ cholesterol (except for cells at passage 17) | |
| ↑ antiparallel β-sheet structures of proteins | |
| ↑ intermolecular β-sheet structures of proteins (except for cells at passage 17) | |
| ↑ phosphocholine | |
| ↑ glycine | |
| ↑ extracellular lactate |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Magalhães, S.; Almeida, I.; Pereira, C.D.; Rebelo, S.; Goodfellow, B.J.; Nunes, A. The Long-Term Culture of Human Fibroblasts Reveals a Spectroscopic Signature of Senescence. Int. J. Mol. Sci. 2022, 23, 5830. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105830
Magalhães S, Almeida I, Pereira CD, Rebelo S, Goodfellow BJ, Nunes A. The Long-Term Culture of Human Fibroblasts Reveals a Spectroscopic Signature of Senescence. International Journal of Molecular Sciences. 2022; 23(10):5830. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105830
Chicago/Turabian StyleMagalhães, Sandra, Idália Almeida, Cátia D. Pereira, Sandra Rebelo, Brian J. Goodfellow, and Alexandra Nunes. 2022. "The Long-Term Culture of Human Fibroblasts Reveals a Spectroscopic Signature of Senescence" International Journal of Molecular Sciences 23, no. 10: 5830. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23105830