
Retinal Proteomic Analysis in a Mouse Model of Endotoxin-Induced Uveitis Using Data-Independent Acquisition-Based Mass Spectrometry

 , , , and
, , , and
Abstract
:1. Introduction
2. Results
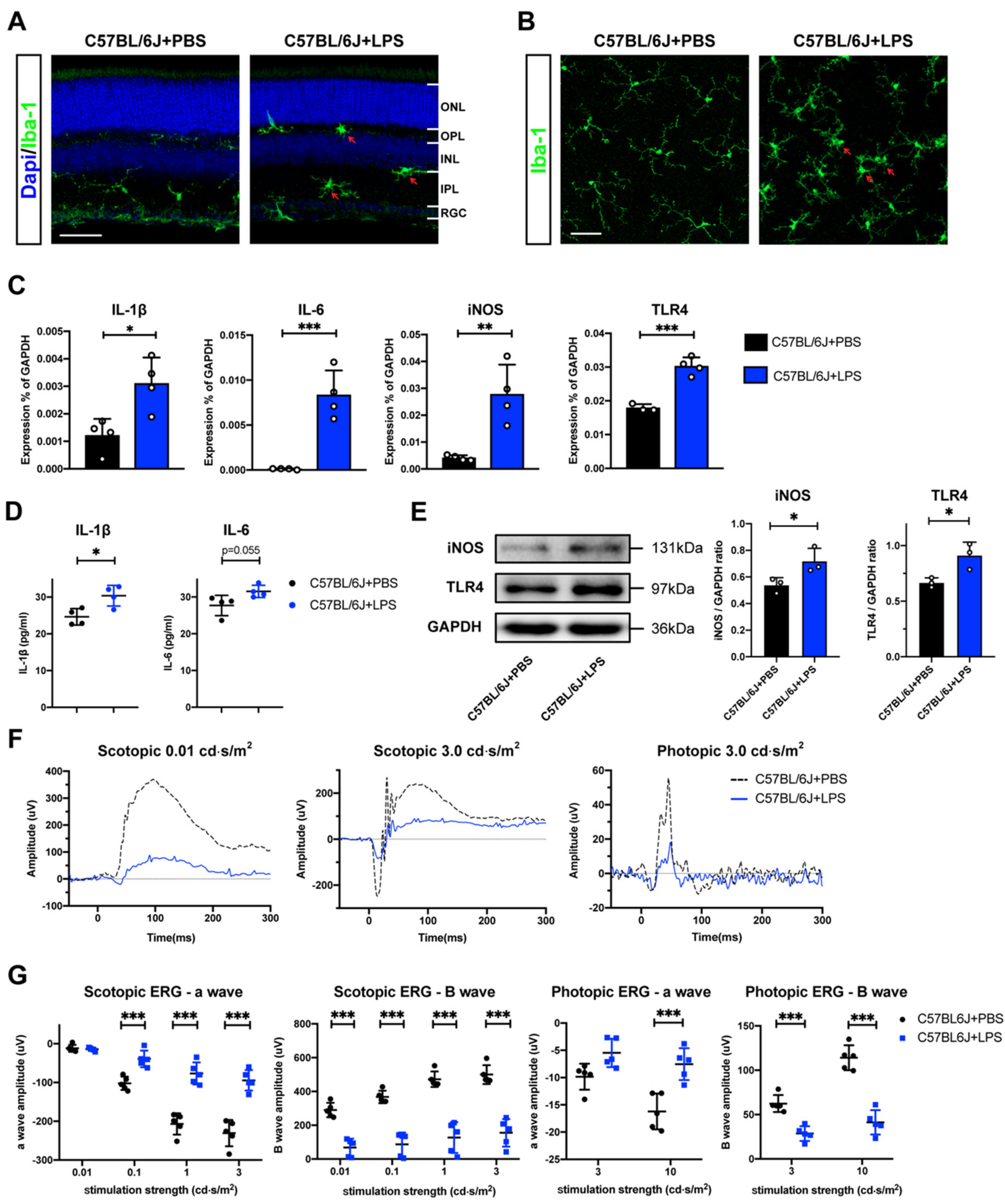
2.1. Retinal Evaluation of the EIU Mouse Model
2.2. Ion Library Generation Using a Pooled Retinal Proteome
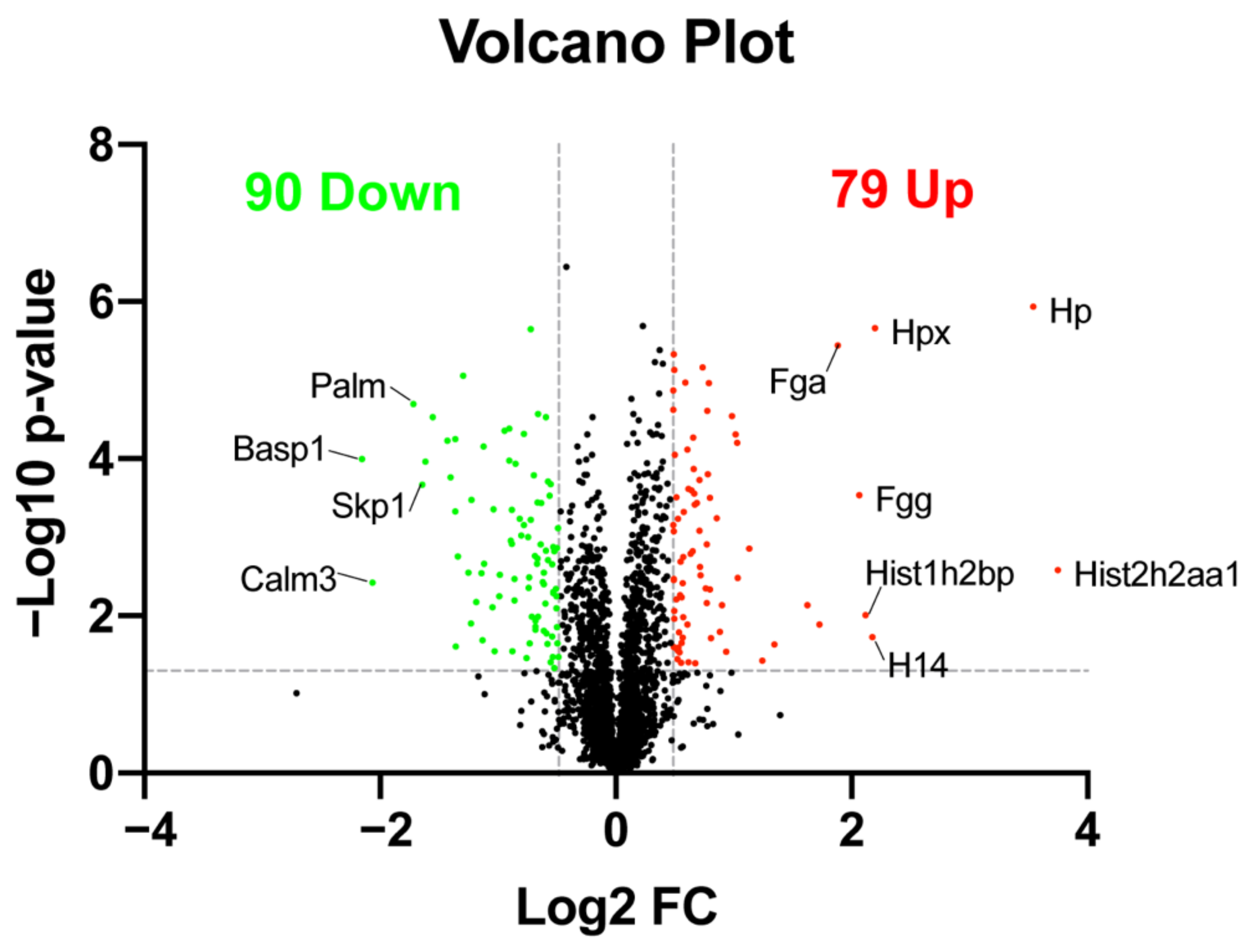
2.3. Quantitative Analysis of Differentially Expressed Proteins
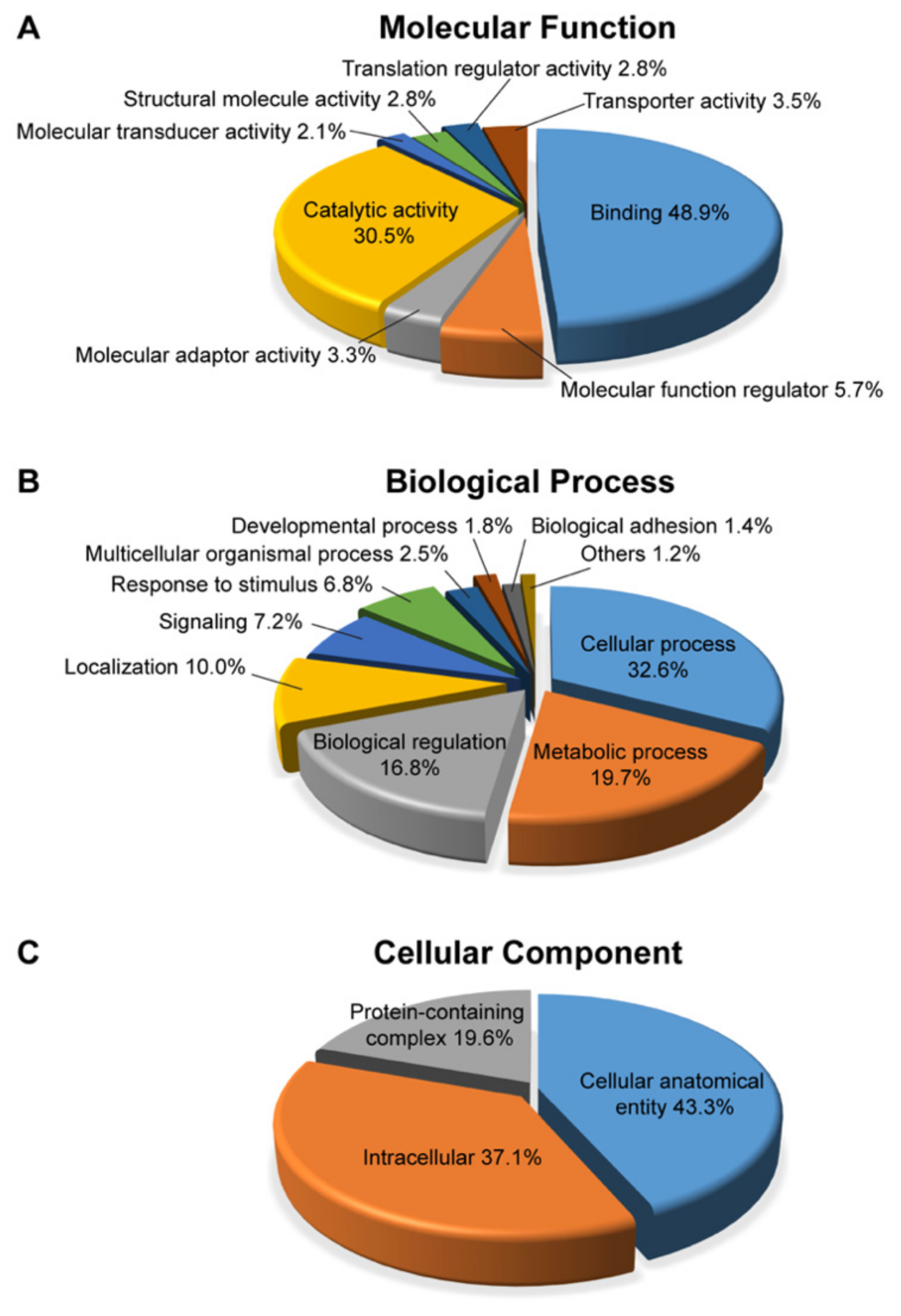
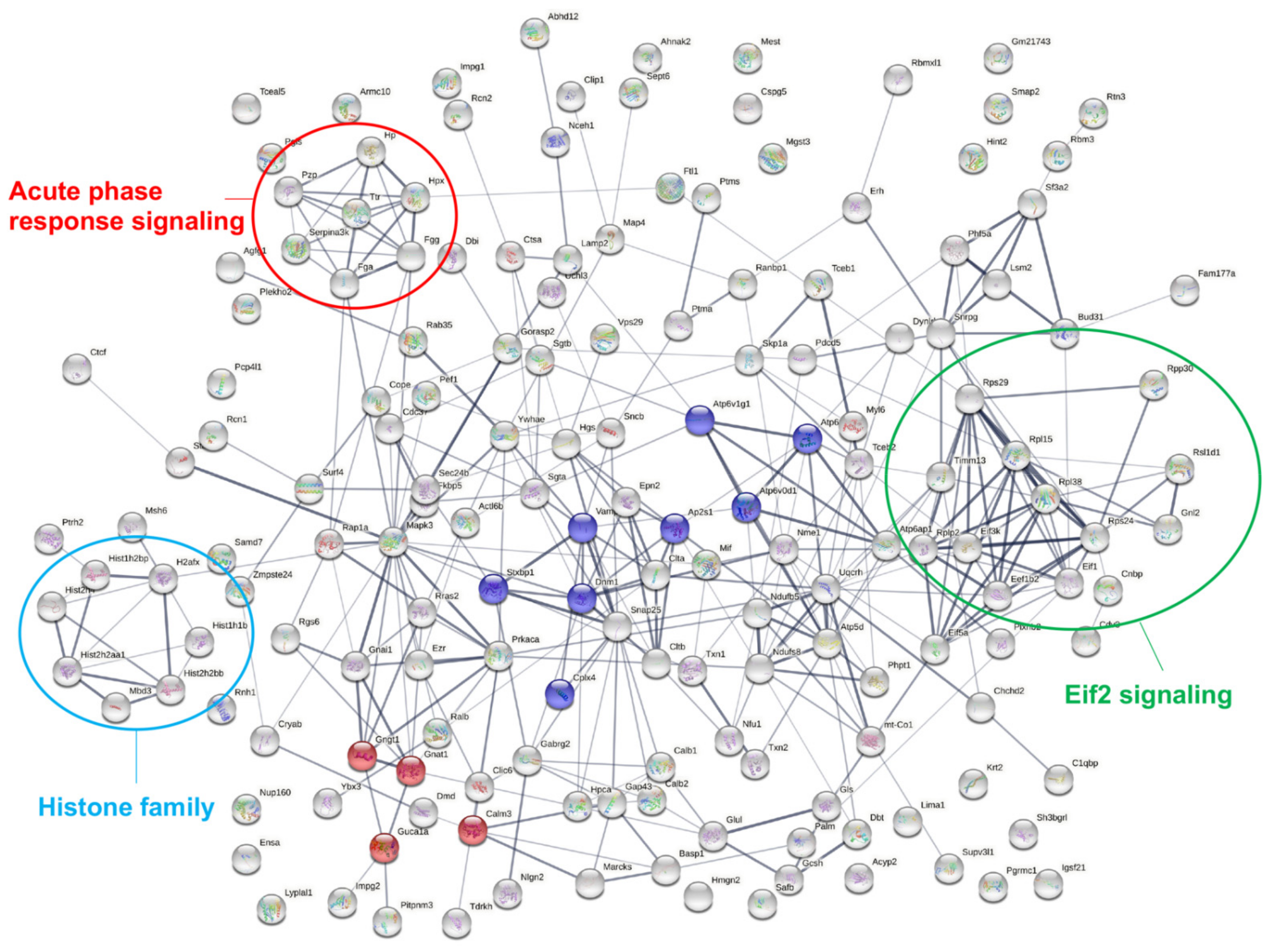
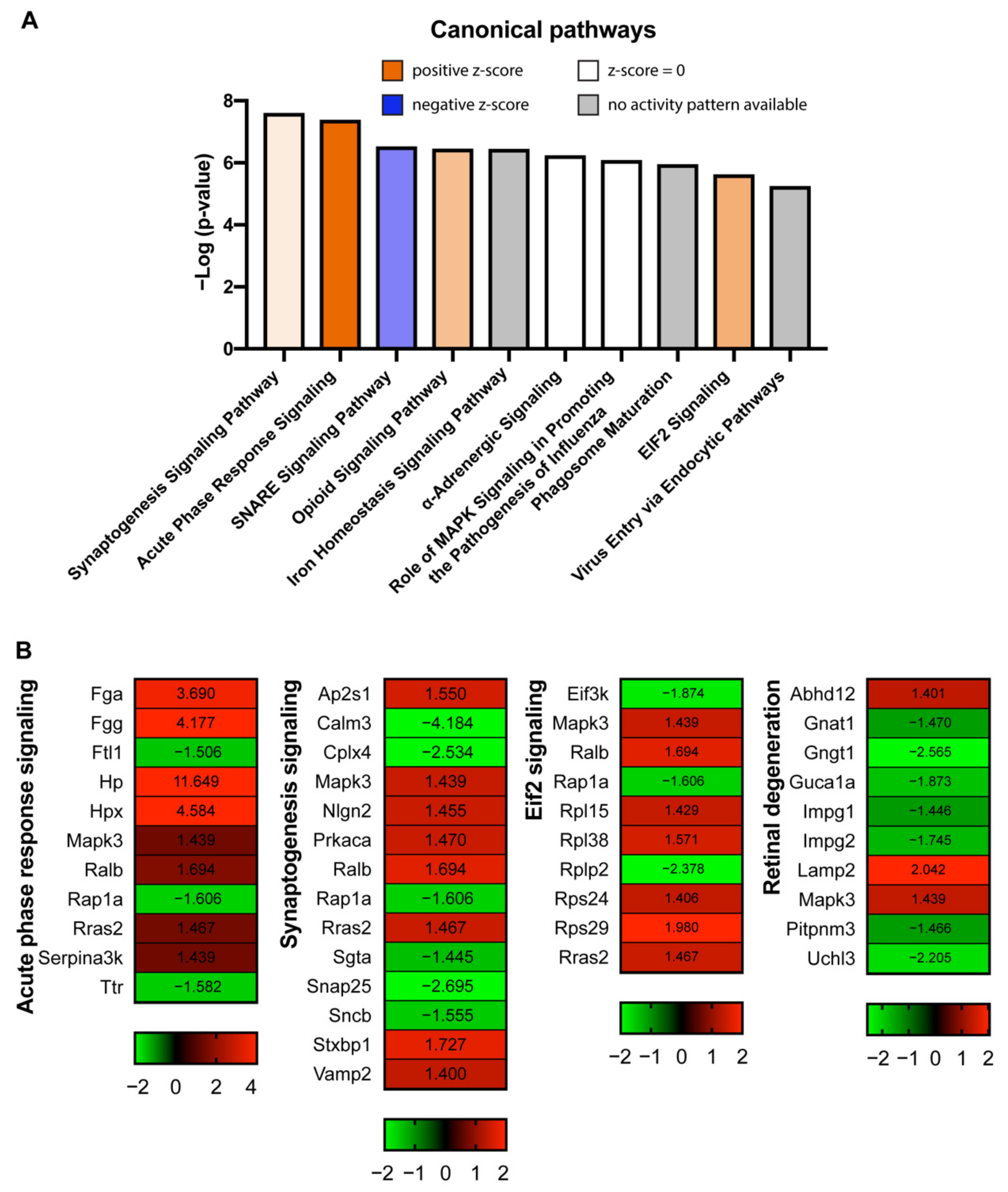
2.4. Pathway Analysis of Differentially Expressed Proteins
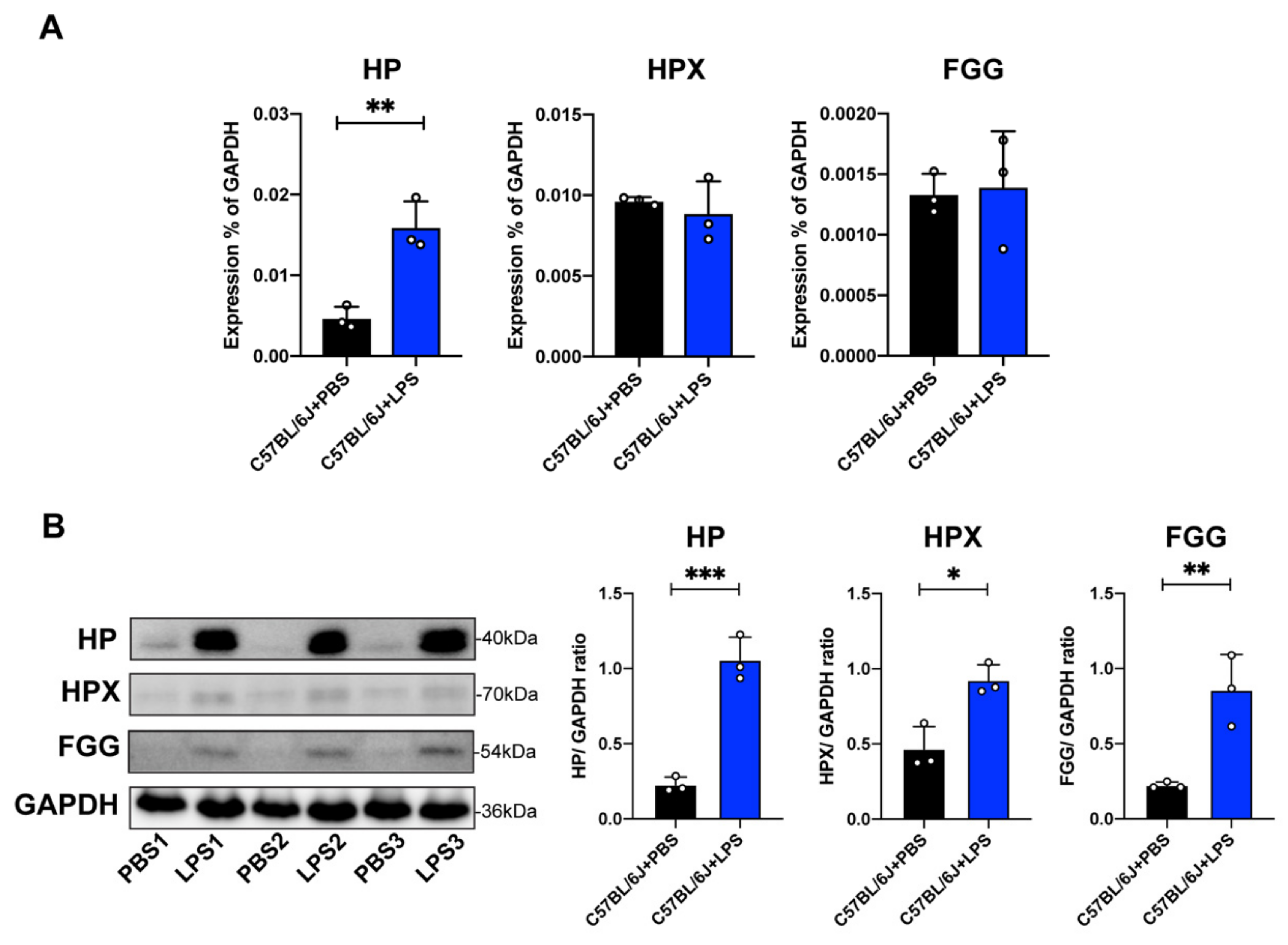
2.5. Validation of Proteins in Acute Phase Response Signaling
3. Discussion
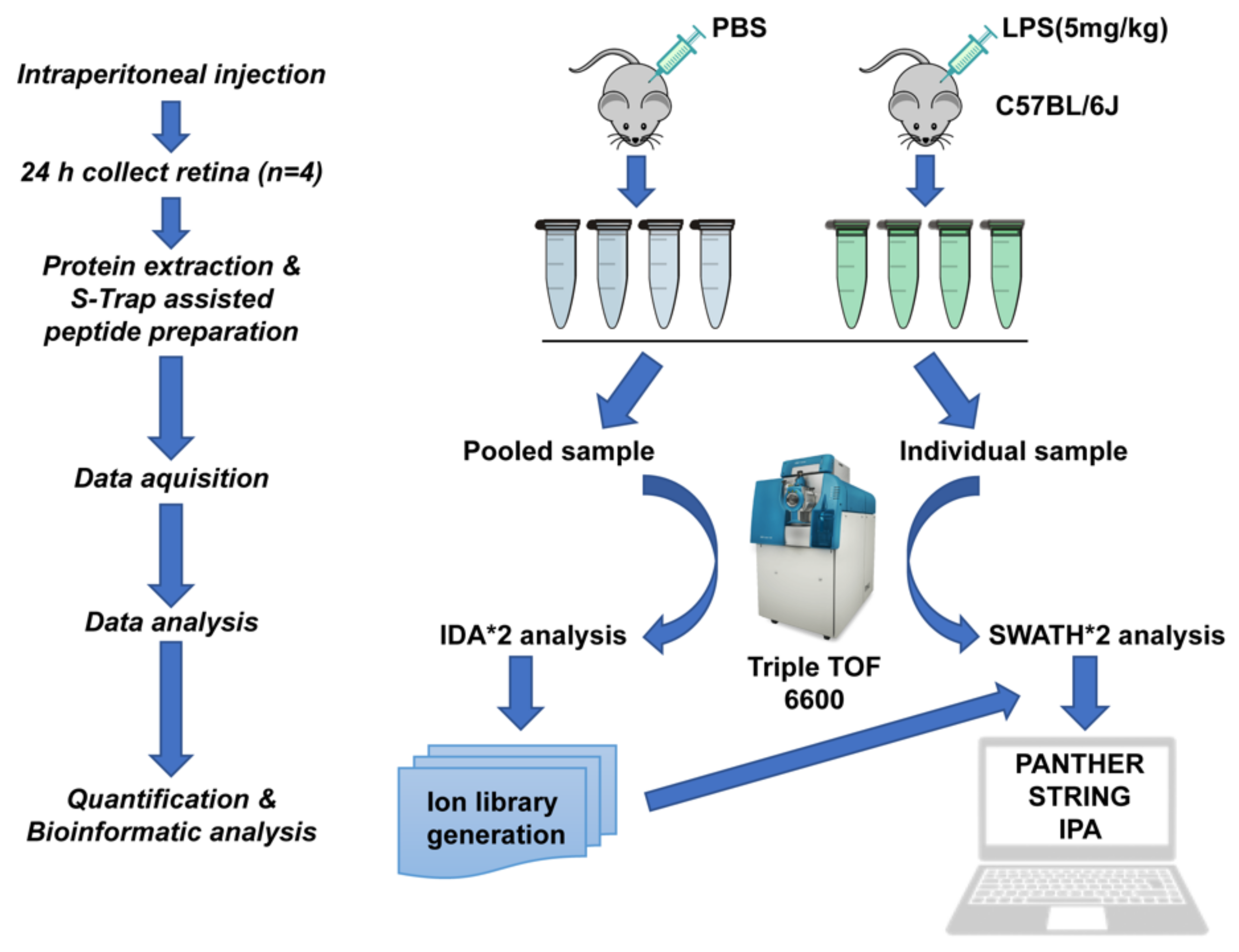
4. Material and Methods
4.1. Animals and LPS Administration
4.2. Electroretinogram (ERG) Analysis
4.3. Retina Tissue Extraction
4.4. Immunostaining
4.5. qPCR
4.6. Protein Extraction from Retina Tissues
4.7. S-Trap-Based Peptide Preparation
4.8. Protein Identification and SWATH-MS
4.9. Bioinformatic Analysis
4.10. Western Blot
4.11. ELISA
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Suttorp-Schulten, M.S.; Rothova, A. The possible impact of uveitis in blindness: A literature survey. Br. J. Ophthalmol. 1996, 80, 844–848. [Google Scholar] [CrossRef] [Green Version]
- Miserocchi, E.; Fogliato, G.; Modorati, G.; Bandello, F. Review on the worldwide epidemiology of uveitis. Eur. J. Ophthalmol. 2013, 23, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Agarwal, A.; Mahendradas, P.; Lee, C.S.; Gupta, V.; Pavesio, C.E.; Agrawal, R. Viral posterior uveitis. Surv. Ophthalmol. 2017, 62, 404–445. [Google Scholar] [CrossRef] [PubMed]
- Hart, C.T.; Zhu, E.Y.; Crock, C.; Rogers, S.L.; Lim, L.L. Epidemiology of uveitis in urban Australia. Clin. Exp. Ophthalmol. 2019, 47, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Shirahama, S.; Kaburaki, T.; Nakahara, H.; Tanaka, R.; Takamoto, M.; Fujino, Y.; Kawashima, H.; Aihara, M. Epidemiology of uveitis (2013–2015) and changes in the patterns of uveitis (2004–2015) in the central Tokyo area: A retrospective study. BMC Ophthalmol. 2018, 18, 189. [Google Scholar] [CrossRef] [Green Version]
- Lee, I.; Zickuhr, L.; Hassman, L. Update on ophthalmic manifestations of systemic lupus erythematosus: Pathogenesis and precision medicine. Curr. Opin. Ophthalmol. 2021, 32, 583–589. [Google Scholar] [CrossRef]
- Joye, A.; Suhler, E. Vogt-Koyanagi-Harada disease. Curr. Opin. Ophthalmol. 2021, 32, 574–582. [Google Scholar] [CrossRef]
- Rothova, A.; Buitenhuis, H.J.; Meenken, C.; Brinkman, C.J.; Linssen, A.; Alberts, C.; Luyendijk, L.; Kijlstra, A. Uveitis and systemic disease. Br. J. Ophthalmol. 1992, 76, 137–141. [Google Scholar] [CrossRef] [Green Version]
- Yadav, U.C.; Ramana, K.V. Endotoxin-induced uveitis in rodents. Methods Mol. Biol. 2013, 1031, 155–162. [Google Scholar] [CrossRef]
- Rosenbaum, J.T.; McDevitt, H.O.; Guss, R.B.; Egbert, P.R. Endotoxin-induced uveitis in rats as a model for human disease. Nature 1980, 286, 611–613. [Google Scholar] [CrossRef]
- Yates, J.R.; Ruse, C.I.; Nakorchevsky, A. Proteomics by mass spectrometry: Approaches, advances, and applications. Annu. Rev. Biomed. Eng. 2009, 11, 49–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, J.; Sze, Y.H.; Tse, D.Y.; To, C.H.; McFadden, S.A.; Lam, C.S.; Li, K.K.; Lam, T.C. SWATH Based Quantitative Proteomics Reveals Significant Lipid Metabolism in Early Myopic Guinea Pig Retina. Int. J. Mol. Sci. 2021, 22, 4721. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Bian, J.F.; Lu, D.Q.; To, C.H.; Lam, C.S.; Li, K.K.; Yu, F.J.; Gong, B.T.; Wang, Q.; Ji, X.W.; et al. Alteration of EIF2 Signaling, Glycolysis, and Dopamine Secretion in Form-Deprived Myopia in Response to 1% Atropine Treatment: Evidence From Interactive iTRAQ-MS and SWATH-MS Proteomics Using a Guinea Pig Model. Front. Pharmacol. 2022, 13, 814814. [Google Scholar] [CrossRef]
- Bansal, R.; Gupta, A. Protein Biomarkers in Uveitis. Front. Immunol. 2020, 11, 610428. [Google Scholar] [CrossRef]
- Bahk, S.C.; Lee, S.H.; Jang, J.U.; Choi, C.U.; Lee, B.S.; Chae, S.C.; Song, H.J.; Park, Z.Y.; Yang, Y.S.; Chung, H.T. Identification of crystallin family proteins in vitreous body in rat endotoxin-induced uveitis: Involvement of crystallin truncation in uveitis pathogenesis. Proteomics 2006, 6, 3436–3444. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Yang, L.; Luo, J.; Guo, L.; Wang, Z.; Yang, X.; Jin, W.; Fang, Y.; Ye, J.; Shan, B.; et al. SWATH enables precise label-free quantification on proteome scale. Proteomics 2015, 15, 1215–1223. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.C.; Yeh, W.C.; Ohashi, P.S. LPS/TLR4 signal transduction pathway. Cytokine 2008, 42, 145–151. [Google Scholar] [CrossRef]
- Okunuki, Y.; Mukai, R.; Nakao, T.; Tabor, S.J.; Butovsky, O.; Dana, R.; Ksander, B.R.; Connor, K.M. Retinal microglia initiate neuroinflammation in ocular autoimmunity. Proc. Natl. Acad. Sci. USA 2019, 116, 9989–9998. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liang, Y.; Lin, B. Accumulation of systematic TPM1 mediates inflammation and neuronal remodeling by phosphorylating PKA and regulating the FABP5/NF-kappaB signaling pathway in the retina of aged mice. Aging Cell 2022, e13566. [Google Scholar] [CrossRef]
- Hoeksema, M.; van Eijk, M.; Haagsman, H.P.; Hartshorn, K.L. Histones as mediators of host defense, inflammation and thrombosis. Future Microbiol. 2016, 11, 441–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.S.; Cho, J.H.; Park, H.W.; Yoon, H.; Kim, M.S.; Kim, S.C. Endotoxin-neutralizing antimicrobial proteins of the human placenta. J. Immunol. 2002, 168, 2356–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augusto, L.A.; Decottignies, P.; Synguelakis, M.; Nicaise, M.; Le Marechal, P.; Chaby, R. Histones: A novel class of lipopolysaccharide-binding molecules. Biochemistry 2003, 42, 3929–3938. [Google Scholar] [CrossRef] [PubMed]
- Abrams, S.T.; Zhang, N.; Dart, C.; Wang, S.S.; Thachil, J.; Guan, Y.; Wang, G.; Toh, C.H. Human CRP defends against the toxicity of circulating histones. J. Immunol. 2013, 191, 2495–2502. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Lu, H.; Dutt, K.; Smith, A.; Hunt, D.M.; Hunt, R.C. Expression of the protective proteins hemopexin and haptoglobin by cells of the neural retina. Exp. Eye Res. 1998, 67, 83–93. [Google Scholar] [CrossRef]
- Hernandez, C.; Garcia-Ramirez, M.; Simo, R. Overexpression of hemopexin in the diabetic eye: A new pathogenic candidate for diabetic macular edema. Diabetes Care 2013, 36, 2815–2821. [Google Scholar] [CrossRef] [Green Version]
- Chen-Roetling, J.; Li, Y.; Cao, Y.; Yan, Z.; Lu, X.; Regan, R.F. Effect of hemopexin treatment on outcome after intracerebral hemorrhage in mice. Brain Res. 2021, 1765, 147507. [Google Scholar] [CrossRef]
- Ma, B.; Day, J.P.; Phillips, H.; Slootsky, B.; Tolosano, E.; Dore, S. Deletion of the hemopexin or heme oxygenase-2 gene aggravates brain injury following stroma-free hemoglobin-induced intracerebral hemorrhage. J. Neuroinflammation 2016, 13, 26. [Google Scholar] [CrossRef] [Green Version]
- Leclerc, J.L.; Li, C.; Jean, S.; Lampert, A.S.; Amador, C.L.; Diller, M.A.; Tolosano, E.; Dore, S. Temporal and age-dependent effects of haptoglobin deletion on intracerebral hemorrhage-induced brain damage and neurobehavioral outcomes. Exp. Neurol. 2019, 317, 22–33. [Google Scholar] [CrossRef]
- Schachtrup, C.; Lu, P.; Jones, L.L.; Lee, J.K.; Lu, J.; Sachs, B.D.; Zheng, B.; Akassoglou, K. Fibrinogen inhibits neurite outgrowth via beta 3 integrin-mediated phosphorylation of the EGF receptor. Proc. Natl. Acad. Sci. USA 2007, 104, 11814–11819. [Google Scholar] [CrossRef] [Green Version]
- Merlini, M.; Rafalski, V.A.; Rios Coronado, P.E.; Gill, T.M.; Ellisman, M.; Muthukumar, G.; Subramanian, K.S.; Ryu, J.K.; Syme, C.A.; Davalos, D.; et al. Fibrinogen Induces Microglia-Mediated Spine Elimination and Cognitive Impairment in an Alzheimer’s Disease Model. Neuron 2019, 101, 1099–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bian, Z.; Yamashita, T.; Shi, X.; Feng, T.; Yu, H.; Hu, X.; Hu, X.; Bian, Y.; Sun, H.; Tadokoro, K.; et al. Accelerated accumulation of fibrinogen peptide chains with Abeta deposition in Alzheimer’s disease (AD) mice and human AD brains. Brain Res. 2021, 1767, 147569. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.; Seilhamer, J. A comparison of selected mRNA and protein abundances in human liver. Electrophoresis 1997, 18, 533–537. [Google Scholar] [CrossRef]
- Tian, N. Visual experience and maturation of retinal synaptic pathways. Vision Res. 2004, 44, 3307–3316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petralia, R.S.; Mattson, M.P.; Yao, P.J. Communication breakdown: The impact of ageing on synapse structure. Ageing Res. Rev. 2014, 14, 31–42. [Google Scholar] [CrossRef] [Green Version]
- Rao, J.S.; Kellom, M.; Kim, H.W.; Rapoport, S.I.; Reese, E.A. Neuroinflammation and synaptic loss. Neurochem. Res. 2012, 37, 903–910. [Google Scholar] [CrossRef]
- Witcher, K.G.; Bray, C.E.; Chunchai, T.; Zhao, F.; O’Neil, S.M.; Gordillo, A.J.; Campbell, W.A.; McKim, D.B.; Liu, X.; Dziabis, J.E.; et al. Traumatic Brain Injury Causes Chronic Cortical Inflammation and Neuronal Dysfunction Mediated by Microglia. J. Neurosci. 2021, 41, 1597–1616. [Google Scholar] [CrossRef]
- Shrestha, N.; Bahnan, W.; Wiley, D.J.; Barber, G.; Fields, K.A.; Schesser, K. Eukaryotic initiation factor 2 (eIF2) signaling regulates proinflammatory cytokine expression and bacterial invasion. J. Biol. Chem. 2012, 287, 28738–28744. [Google Scholar] [CrossRef] [Green Version]
- Khan, H.N.; Perlee, D.; Schoenmaker, L.; van der Meer, A.J.; Franitza, M.; Toliat, M.R.; Nurnberg, P.; Zwinderman, A.H.; van der Poll, T.; Scicluna, B.P. Leukocyte transcriptional signatures dependent on LPS dosage in human endotoxemia. J. Leukoc. Biol. 2019, 106, 1153–1160. [Google Scholar] [CrossRef] [Green Version]
- Chang, R.C.; Wong, A.K.; Ng, H.K.; Hugon, J. Phosphorylation of eukaryotic initiation factor-2alpha (eIF2alpha) is associated with neuronal degeneration in Alzheimer’s disease. Neuroreport 2002, 13, 2429–2432. [Google Scholar] [CrossRef]
- Kitiratschky, V.B.; Behnen, P.; Kellner, U.; Heckenlively, J.R.; Zrenner, E.; Jagle, H.; Kohl, S.; Wissinger, B.; Koch, K.W. Mutations in the GUCA1A gene involved in hereditary cone dystrophies impair calcium-mediated regulation of guanylate cyclase. Hum. Mutat. 2009, 30, E782–E796. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.H.; Dizhoor, A.M.; Makino, C.L. Membrane guanylyl cyclase complexes shape the photoresponses of retinal rods and cones. Front. Mol. Neurosci. 2014, 7, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nong, E.; Lee, W.; Merriam, J.E.; Allikmets, R.; Tsang, S.H. Disease progression in autosomal dominant cone-rod dystrophy caused by a novel mutation (D100G) in the GUCA1A gene. Doc. Ophthalmol. 2014, 128, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.; Sheng, X.; Zhuang, W.; Sun, X.; Liu, G.; Shi, X.; Huang, G.; Mei, Y.; Li, Y.; Pan, X.; et al. GUCA1A mutation causes maculopathy in a five-generation family with a wide spectrum of severity. Genet. Med. 2017, 19, 945–954. [Google Scholar] [CrossRef] [Green Version]
- Ruebsam, A.; Dulle, J.E.; Myers, A.M.; Sakrikar, D.; Green, K.M.; Khan, N.W.; Schey, K.; Fort, P.E. A specific phosphorylation regulates the protective role of alphaA-crystallin in diabetes. JCI Insight 2018, 3, e97919. [Google Scholar] [CrossRef] [Green Version]
- Nath, M.; Sluzala, Z.B.; Phadte, A.S.; Shan, Y.; Myers, A.M.; Fort, P.E. Evidence for Paracrine Protective Role of Exogenous alphaA-Crystallin in Retinal Ganglion Cells. Eneuro 2022, 9, ENEURO-0045. [Google Scholar] [CrossRef]
- Saraswathy, S.; Rao, N.A. Mitochondrial proteomics in experimental autoimmune uveitis oxidative stress. Investig. Ophthalmol. Vis. Sci. 2009, 50, 5559–5566. [Google Scholar] [CrossRef]
- Pepple, K.L.; Rotkis, L.; Wilson, L.; Sandt, A.; Van Gelder, R.N. Comparative Proteomic Analysis of Two Uveitis Models in Lewis Rats. Investig. Ophthalmol. Vis. Sci. 2015, 56, 8449–8456. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Ozawa, Y.; Kurihara, T.; Noda, K.; Imamura, Y.; Kobayashi, S.; Ishida, S.; Tsubota, K. Neuroprotective effect of an antioxidant, lutein, during retinal inflammation. Investig. Ophthalmol. Vis. Sci. 2009, 50, 1433–1439. [Google Scholar] [CrossRef] [Green Version]
- Vaczy, A.; Kovari, P.; Kovacs, K.; Farkas, K.; Szabo, E.; Kvarik, T.; Kocsis, B.; Fulop, B.; Atlasz, T.; Reglodi, D. Protective Role of Endogenous PACAP in Inflammation-induced Retinal Degeneration. Curr. Pharm. Des. 2018, 24, 3534–3542. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Baban, B.; Rojas, M.; Tofigh, S.; Virmani, S.K.; Patel, C.; Behzadian, M.A.; Romero, M.J.; Caldwell, R.W.; Caldwell, R.B. Arginase activity mediates retinal inflammation in endotoxin-induced uveitis. Am. J. Pathol. 2009, 175, 891–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Xiong, G.; Lin, B. Cyclooxygenase-1 mediates neuroinflammation and neurotoxicity in a mouse model of retinitis pigmentosa. J. Neuroinflamm. 2020, 17, 306. [Google Scholar] [CrossRef] [PubMed]
- Sze, Y.H.; Zhao, Q.; Cheung, J.K.W.; Li, K.K.; Tse, D.Y.Y.; To, C.H.; Lam, T.C. High-pH reversed-phase fractionated neural retina proteome of normal growing C57BL/6 mouse. Sci. Data 2021, 8, 27. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the PANTHER classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Szklarczyk, D.; Gable, A.L.; Nastou, K.C.; Lyon, D.; Kirsch, R.; Pyysalo, S.; Doncheva, N.T.; Legeay, M.; Fang, T.; Bork, P.; et al. The STRING database in 2021: Customizable protein-protein networks, and functional characterization of user-uploaded gene/measurement sets. Nucleic Acids Res. 2021, 49, D605–D612. [Google Scholar] [CrossRef] [PubMed]
- Kramer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; Garcia-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE database resources in 2022: A hub for mass spectrometry-based proteomics evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| UniProt ID | Gene Name | Protein Name | Fold Change | p-Value |
|---|---|---|---|---|
| B2RWH3 | Hist2h2aa1 | Histone H2A | 13.450 | 0.003 |
| Q3UBS3 | Hp | Haptoglobin | 11.649 | 0.000 |
| Q91X72 | Hpx | Hemopexin | 4.584 | 0.000 |
| P43274 | H14 | Histone H1.4 | 4.519 | 0.019 |
| Q8CGP2 | Hist1h2bp | Histone H2B type 1-P | 4.344 | 0.010 |
| Q3UER8 | Fgg | Fibrinogen gamma chain | 4.177 | 0.000 |
| E9PV24 | Fga | Fibrinogen alpha chain | 3.690 | 0.000 |
| Q64525 | Hist2h2bb | Histone H2B type 2-B | 3.307 | 0.013 |
| Q9D0J8 | Ptms | Parathymosin | 3.082 | 0.007 |
| Q1WWK3 | H1f5 | Hist1h1b protein | 2.537 | 0.023 |
| Q6W8Q3 | Pcp4l1 | Purkinje cell protein 4-like protein 1 | −2.568 | 0.000 |
| Q3TE95 | Rcn2 | Uncharacterized protein | −2.569 | 0.000 |
| Q05186 | Rcn1 | Reticulocalbin-1 | −2.642 | 0.000 |
| P60879 | Snap25 | Synaptosomal-associated protein 25 | −2.695 | 0.000 |
| Q3U6E4 | Ptma | Uncharacterized protein | −2.933 | 0.000 |
| A0A087WPA0 | Lbhd1 | LBH domain-containing 1 | −3.062 | 0.000 |
| Q3TL58 | Skp1 | S-phase kinase-associated protein 1 | −3.117 | 0.000 |
| Q542N8 | Palm | Uncharacterized protein | −3.286 | 0.000 |
| P0DP28 | Calm3 | Calmodulin-3 | −4.183 | 0.004 |
| Q91XV3 | Basp1 | Brain acid soluble protein 1 | −4.443 | 0.000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wu, J.; Lu, D.; To, C.-H.; Lam, T.C.; Lin, B. Retinal Proteomic Analysis in a Mouse Model of Endotoxin-Induced Uveitis Using Data-Independent Acquisition-Based Mass Spectrometry. Int. J. Mol. Sci. 2022, 23, 6464. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126464
Zhang J, Wu J, Lu D, To C-H, Lam TC, Lin B. Retinal Proteomic Analysis in a Mouse Model of Endotoxin-Induced Uveitis Using Data-Independent Acquisition-Based Mass Spectrometry. International Journal of Molecular Sciences. 2022; 23(12):6464. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126464
Chicago/Turabian StyleZhang, Jing, Jiangmei Wu, Daqian Lu, Chi-Ho To, Thomas Chuen Lam, and Bin Lin. 2022. "Retinal Proteomic Analysis in a Mouse Model of Endotoxin-Induced Uveitis Using Data-Independent Acquisition-Based Mass Spectrometry" International Journal of Molecular Sciences 23, no. 12: 6464. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23126464