
Longitudinal Proteomic Analysis of Plasma across Healthy Pregnancies Reveals Indicators of Gestational Age

Abstract
:1. Introduction
2. Results
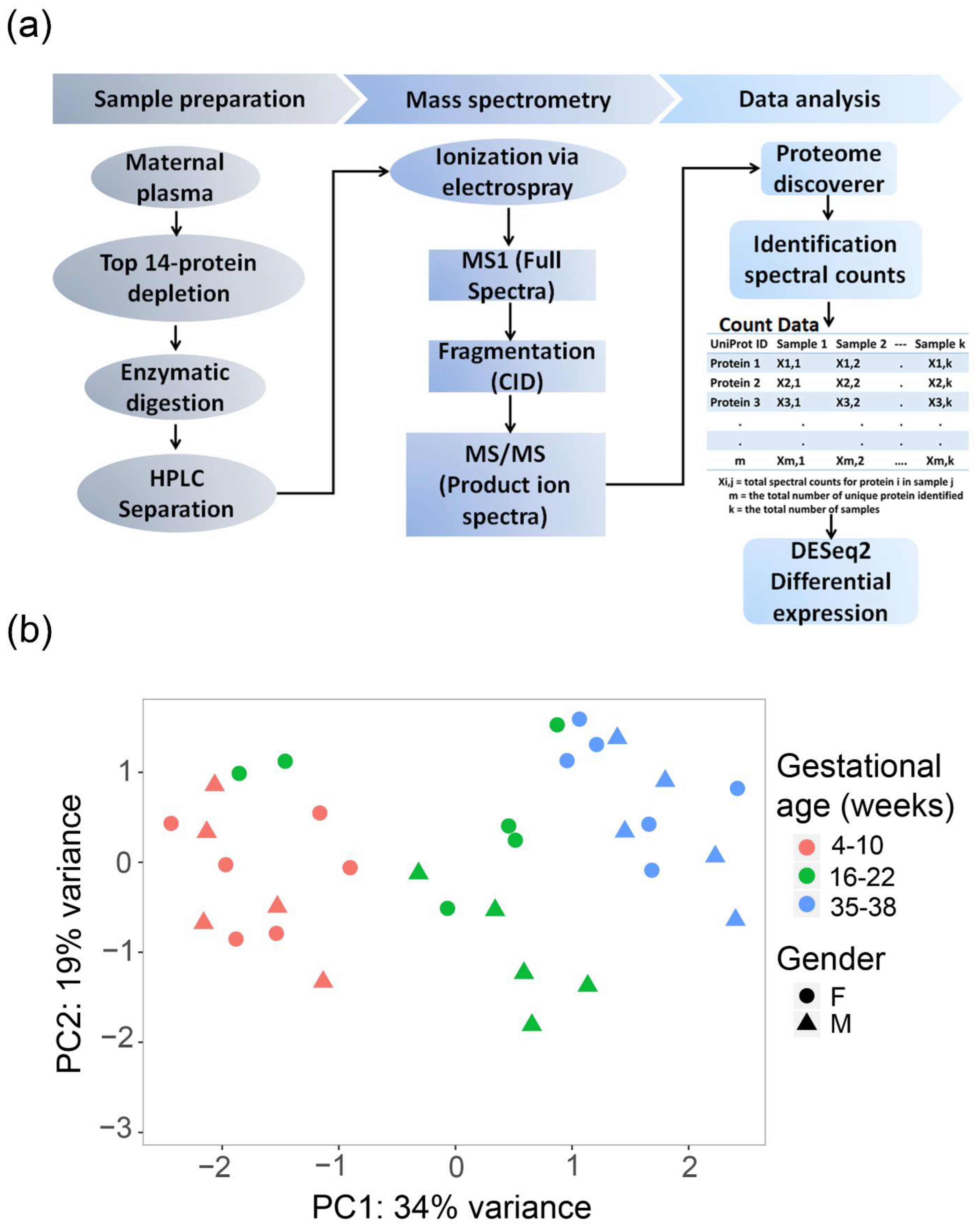
2.1. Principal Component Analysis (PCA)
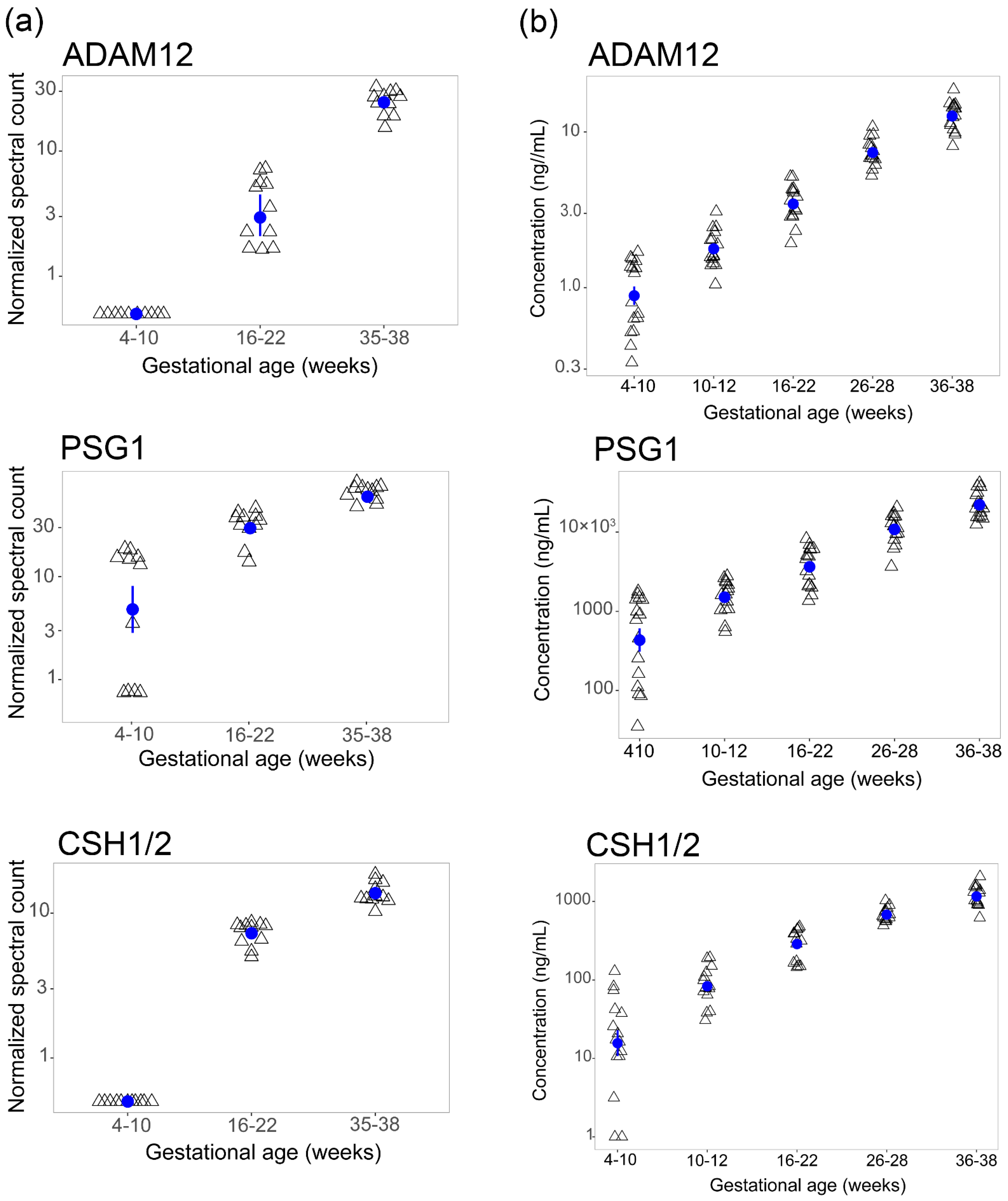
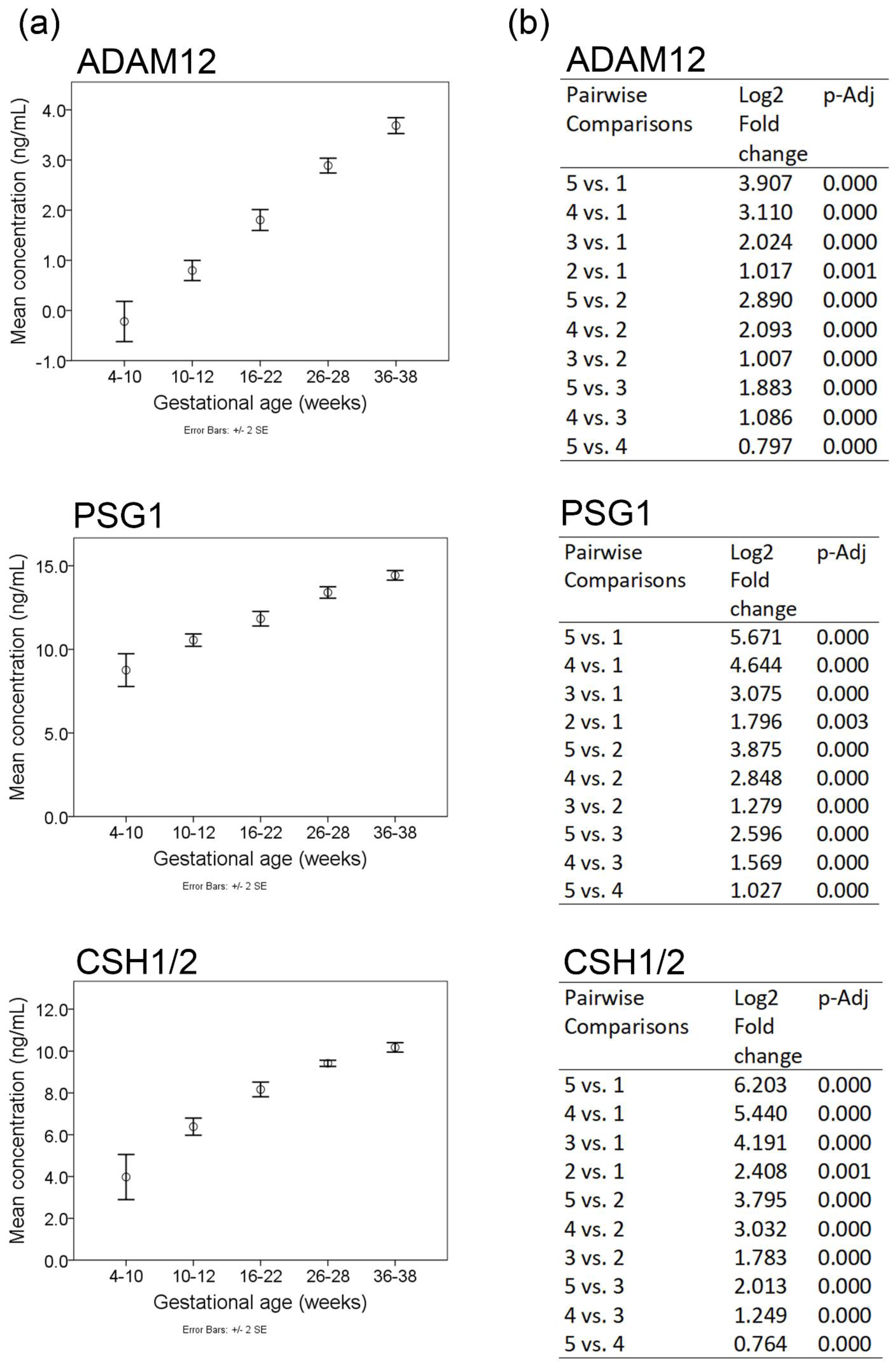
2.2. Differentially Expressed Proteins
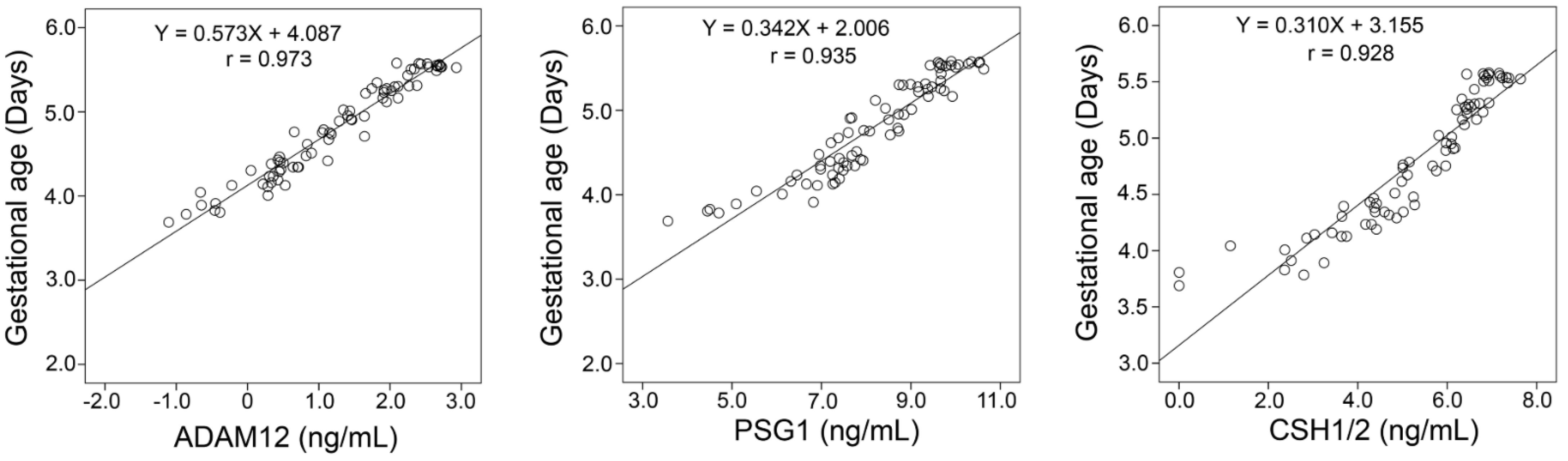
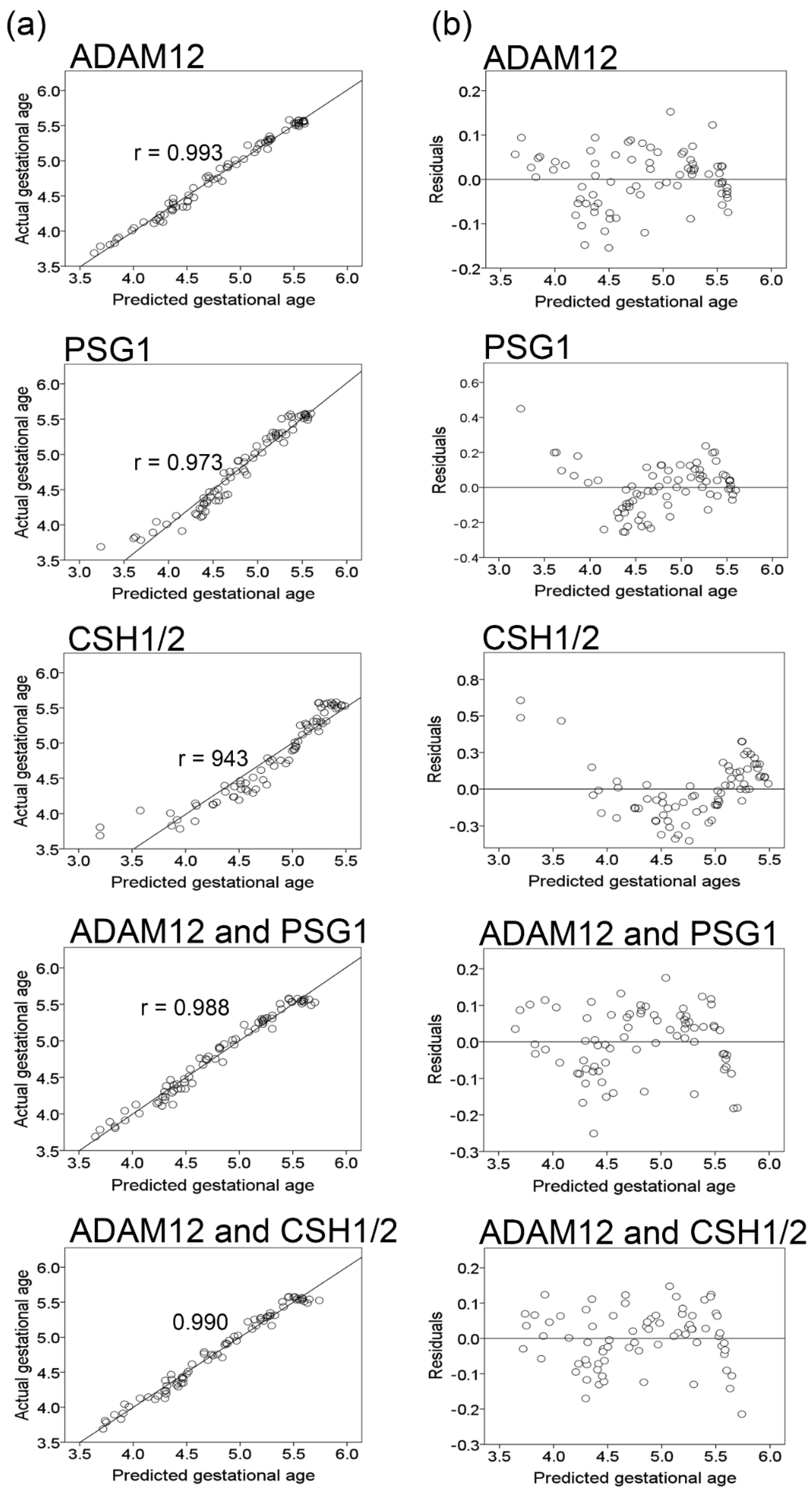
2.3. Orthogonal Validation
3. Discussion
4. Materials and Methods
4.1. Patient Selection and Clinical Samples
4.2. Experimental Design and Statistical Rationale
4.3. Label-Free Shotgun LC-MS/MS
4.4. Protein Identification and Data Search Parameters
4.5. Quantitation and Statistical Analysis
4.6. High-Dimensional Data Visualization
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. ELISA Data Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Salomon, L.J.; Alfirevic, Z.; Bilardo, C.M.; Chalouhi, G.E.; Ghi, T.; Kagan, K.O.; Lau, T.K.; Papageorghiou, A.T.; Raine-Fenning, N.J.; Stirnemann, J.; et al. ISUOG practice guidelines: Performance of first-trimester fetal ultrasound scan. Ultrasound Obstet. Gynecol. 2013, 41, 102–113. [Google Scholar] [CrossRef]
- Verburg, B.O.; Steegers, E.A.; De Ridder, M.; Snijders, R.J.; Smith, E.; Hofman, A.; Moll, H.A.; Jaddoe, V.W.; Witteman, J.C. New charts for ultrasound dating of pregnancy and assessment of fetal growth: Longitudinal data from a population-based cohort study. Ultrasound Obstet. Gynecol. 2008, 31, 388–396. [Google Scholar] [CrossRef]
- Vintzileos, A.M.; Ananth, C.V.; Smulian, J.C. Using ultrasound in the clinical management of placental implantation abnormalities. Am. J. Obstet. Gynecol. 2015, 213, S70–S77. [Google Scholar] [CrossRef]
- Tsang, J.C.H.; Vong, J.S.L.; Ji, L.; Poon, L.C.Y.; Jiang, P.; Lui, K.O.; Ni, Y.B.; To, K.F.; Cheng, Y.K.Y.; Chiu, R.W.K.; et al. Integrative single-cell and cell-free plasma RNA transcriptomics elucidates placental cellular dynamics. Proc. Natl. Acad. Sci. USA 2017, 114, E7786–E7795. [Google Scholar] [CrossRef] [Green Version]
- Pavlicev, M.; Wagner, G.P.; Chavan, A.R.; Owens, K.; Maziarz, J.; Dunn-Fletcher, C.; Kallapur, S.G.; Muglia, L.; Jones, H. Single-cell transcriptomics of the human placenta: Inferring the cell communication network of the maternal-fetal interface. Genome Res. 2017, 27, 349–361. [Google Scholar] [CrossRef]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.E.; Stephenson, E.; Polanski, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [Green Version]
- Romero, R.; Erez, O.; Maymon, E.; Chaemsaithong, P.; Xu, Z.; Pacora, P.; Chaiworapongsa, T.; Done, B.; Hassan, S.S.; Tarca, A.L. The maternal plasma proteome changes as a function of gestational age in normal pregnancy: A longitudinal study. Am. J. Obstet. Gynecol. 2017, 217, 67.e1–67.e21. [Google Scholar] [CrossRef]
- Aghaeepour, N.; Lehallier, B.; Baca, Q.; Ganio, E.A.; Wong, R.J.; Ghaemi, M.S.; Culos, A.; El-Sayed, Y.Y.; Blumenfeld, Y.J.; Druzin, M.L.; et al. A proteomic clock of human pregnancy. Am. J. Obstet. Gynecol. 2018, 218, 347.e1–347.e14. [Google Scholar] [CrossRef] [Green Version]
- Del Priore, G.; Chatterton, R., Jr.; Lee, C.; Silver, R.; Berg, L.; Lee, M.J. Comparison of mononuclear cell proteins and plasma proteins before and during parturition by two dimensional electrophoresis. J. Perinat. Med. 1991, 19, 373–377. [Google Scholar] [CrossRef]
- Belo, L.; Santos-Silva, A.; Rocha, S.; Caslake, M.; Cooney, J.; Pereira-Leite, L.; Quintanilha, A.; Rebelo, I. Fluctuations in C-reactive protein concentration and neutrophil activation during normal human pregnancy. Eur. J. Obstet. Gynecol. Reprod. Biol. 2005, 123, 46–51. [Google Scholar] [CrossRef]
- Joseph, J.C.; Baker, C.; Sprang, M.L.; Bermes, E.W. Changes in plasma proteins during pregnancy. Ann. Clin. Lab. Sci. 1978, 8, 130–141. [Google Scholar]
- Tarca, A.L.; Romero, R.; Benshalom-Tirosh, N.; Than, N.G.; Gudicha, D.W.; Done, B.; Pacora, P.; Chaiworapongsa, T.; Panaitescu, B.; Tirosh, D.; et al. The prediction of early preeclampsia: Results from a longitudinal proteomics study. PLoS ONE 2019, 14, e0217273. [Google Scholar] [CrossRef]
- Hedman, A.M.; Lundholm, C.; Andolf, E.; Pershagen, G.; Fall, T.; Almqvist, C. Longitudinal plasma inflammatory proteome profiling during pregnancy in the Born into Life study. Sci. Rep. 2020, 10, 17819. [Google Scholar] [CrossRef]
- Max, K.E.A.; Bertram, K.; Akat, K.M.; Bogardus, K.A.; Li, J.; Morozov, P.; Ben-Dov, I.Z.; Li, X.; Weiss, Z.R.; Azizian, A.; et al. Human plasma and serum extracellular small RNA reference profiles and their clinical utility. Proc. Natl. Acad. Sci. USA 2018, 115, E5334–E5343. [Google Scholar] [CrossRef] [Green Version]
- Anderson, N.L.; Anderson, N.G. The human plasma proteome: History, character, and diagnostic prospects. Mol. Cell. Proteom. 2002, 1, 845–867. [Google Scholar] [CrossRef] [Green Version]
- Keshishian, H.; Burgess, M.W.; Gillette, M.A.; Mertins, P.; Clauser, K.R.; Mani, D.R.; Kuhn, E.W.; Farrell, L.A.; Gerszten, R.E.; Carr, S.A. Multiplexed, Quantitative Workflow for Sensitive Biomarker Discovery in Plasma Yields Novel Candidates for Early Myocardial Injury. Mol. Cell. Proteom. 2015, 14, 2375–2393. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.; Kim, J.; Strittmatter, E.F.; Jacobs, J.M.; Camp, D.G., 2nd; Fang, R.; Tolie, N.; Moore, R.J.; Smith, R.D. Characterization of the human blood plasma proteome. Proteomics 2005, 5, 4034–4045. [Google Scholar] [CrossRef]
- Zhang, Y.; Fonslow, B.R.; Shan, B.; Baek, M.C.; Yates, J.R., 3rd. Protein analysis by shotgun/bottom-up proteomics. Chem. Rev. 2013, 113, 2343–2394. [Google Scholar] [CrossRef] [Green Version]
- Joshi, A.; Mayr, M. In Aptamers They Trust: The Caveats of the SOMAscan Biomarker Discovery Platform from SomaLogic. Circulation 2018, 138, 2482–2485. [Google Scholar] [CrossRef]
- Yohannes, E.; Kazanjian, A.A.; Lindsay, M.E.; Fujii, D.T.; Ieronimakis, N.; Chow, G.E.; Beesley, R.D.; Heitmann, R.J.; Burney, R.O. The human tubal lavage proteome reveals biological processes that may govern the pathology of hydrosalpinx. Sci. Rep. 2019, 9, 8980. [Google Scholar] [CrossRef] [Green Version]
- Candia, J.; Cheung, F.; Kotliarov, Y.; Fantoni, G.; Sellers, B.; Griesman, T.; Huang, J.; Stuccio, S.; Zingone, A.; Ryan, B.M.; et al. Assessment of Variability in the SOMAscan Assay. Sci. Rep. 2017, 7, 14248. [Google Scholar] [CrossRef] [Green Version]
- Benson, M.D.; Ngo, D.; Ganz, P.; Gerszten, R.E. Emerging Affinity Reagents for High Throughput Proteomics: Trust, but Verify. Circulation 2019, 140, 1610–1612. [Google Scholar] [CrossRef]
- Bracamontes, C.G.; Lopez-Valdez, R.; Subramani, R.; Arumugam, A.; Nandy, S.; Rajamanickam, V.; Ravichandran, V.; Lakshmanaswamy, R. The serum protein profile of early parity which induces protection against breast cancer. Oncotarget 2016, 7, 82538–82553. [Google Scholar] [CrossRef] [Green Version]
- Wallace, J.M.; Bhattacharya, S.; Horgan, G.W. Gestational age, gender and parity specific centile charts for placental weight for singleton deliveries in Aberdeen, UK. Placenta 2013, 34, 269–274. [Google Scholar] [CrossRef]
- Laigaard, J.; Sorensen, T.; Placing, S.; Holck, P.; Frohlich, C.; Wojdemann, K.R.; Sundberg, K.; Shalmi, A.C.; Tabor, A.; Norgaard-Pedersen, B.; et al. Reduction of the disintegrin and metalloprotease ADAM12 in preeclampsia. Obstet. Gynecol. 2005, 106, 144–149. [Google Scholar] [CrossRef]
- Laigaard, J.; Cuckle, H.; Wewer, U.M.; Christiansen, M. Maternal serum ADAM12 levels in Down and Edwards’ syndrome pregnancies at 9–12 weeks’ gestation. Prenat. Diagn. 2006, 26, 689–691. [Google Scholar] [CrossRef]
- Cowans, N.J.; Spencer, K. First-trimester ADAM12 and PAPP-A as markers for intrauterine fetal growth restriction through their roles in the insulin-like growth factor system. Prenat. Diagn. 2007, 27, 264–271. [Google Scholar] [CrossRef]
- Sahraravand, M.; Jarvela, I.Y.; Laitinen, P.; Tekay, A.H.; Ryynanen, M. The secretion of PAPP-A, ADAM12, and PP13 correlates with the size of the placenta for the first month of pregnancy. Placenta 2011, 32, 999–1003. [Google Scholar] [CrossRef]
- Wortelboer, E.J.; Koster, M.P.; Kuc, S.; Eijkemans, M.J.; Bilardo, C.M.; Schielen, P.C.; Visser, G.H. Longitudinal trends in fetoplacental biochemical markers, uterine artery pulsatility index and maternal blood pressure during the first trimester of pregnancy. Ultrasound Obstet. Gynecol. 2011, 38, 383–388. [Google Scholar] [CrossRef]
- Aghababaei, M.; Hogg, K.; Perdu, S.; Robinson, W.P.; Beristain, A.G. ADAM12-directed ectodomain shedding of E-cadherin potentiates trophoblast fusion. Cell Death Differ. 2015, 22, 1970–1984. [Google Scholar] [CrossRef] [Green Version]
- Grudzinskas, J.G.; Gordon, Y.B.; Menabawey, M.; Lee, J.N.; Wadsworth, J.; Chard, T. Identification of high-risk pregnancy by the routine measurement of pregnancy-specific beta 1-glycoprotein. Am. J. Obstet. Gynecol. 1983, 147, 10–12. [Google Scholar] [CrossRef]
- Pihl, K.; Larsen, T.; Laursen, I.; Krebs, L.; Christiansen, M. First trimester maternal serum pregnancy-specific beta-1-glycoprotein (SP1) as a marker of adverse pregnancy outcome. Prenat. Diagn. 2009, 29, 1256–1261. [Google Scholar] [CrossRef]
- Salem, H.T.; Lee, J.N.; Seppala, M.; Vaara, L.; Aula, P.; Al-Ani, A.T.; Chard, T. Measurement of placental protein 5, placental lactogen and pregnancy-specific beta 1 glycoprotein in mid-trimester as a predictor of outcome of pregnancy. Br. J. Obstet. Gynaecol. 1981, 88, 371–374. [Google Scholar] [CrossRef]
- Towler, C.M.; Horne, C.H.; Jandial, V.; Campbell, D.M.; MacGillivray, I. Plasma levels of pregnancy-specific beta 1-glycoprotein in complicated pregnancies. Br. J. Obstet. Gynaecol. 1977, 84, 258–263. [Google Scholar] [CrossRef]
- Wurz, H.; Geiger, W.; Kunzig, H.J.; Jabs-Lehmann, A.; Bohn, H.; Luben, G. Radioimmunoassay of SP1 (pregnancy-specific beta1-glycoprotein) in maternal blood and in amniotic fluid normal and pathologic pregnancies. J. Perinat. Med. 1981, 9, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Cui, Y.; He, L.; Yang, C.Y.; Ye, Q. iTRAQ and PRM-based quantitative proteomics in early recurrent spontaneous abortion: Biomarkers discovery. Clin. Proteom. 2019, 16, 36. [Google Scholar] [CrossRef]
- Arnold, L.L.; Doherty, T.M.; Flor, A.W.; Simon, J.A.; Chou, J.Y.; Chan, W.Y.; Mansfield, B.C. Pregnancy-specific glycoprotein gene expression in recurrent aborters: A potential correlation to interleukin-10 expression. Am. J. Reprod. Immunol. 1999, 41, 174–182. [Google Scholar] [CrossRef]
- Bersinger, N.A.; Odegard, R.A. Second- and third-trimester serum levels of placental proteins in preeclampsia and small-for-gestational age pregnancies. Acta Obstet. Gynecol. Scand. 2004, 83, 37–45. [Google Scholar] [CrossRef]
- Silver, R.M.; Heyborne, K.D.; Leslie, K.K. Pregnancy specific beta 1 glycoprotein (SP-1) in maternal serum and amniotic fluid; pre-eclampsia, small for gestational age fetus and fetal distress. Placenta 1993, 14, 583–589. [Google Scholar] [CrossRef]
- Spellacy, W.N.; Carlson, K.L.; Birk, S.A. Dynamics of human placental lactogen. Am. J. Obstet. Gynecol. 1966, 96, 1164–1173. [Google Scholar] [CrossRef]
- Spellacy, W.N.; Carlson, K.L.; Birk, S.A. Human placental lactogen levels as a variable of placental weight and infant weight. Am. J. Obstet. Gynecol. 1966, 95, 118–119. [Google Scholar] [CrossRef]
- Saxena, B.N.; Emerson, K., Jr.; Selenkow, H.A. Serum placental lactogen (HPL) levels as an index of placental function. N. Engl. J. Med. 1969, 281, 225–231. [Google Scholar] [CrossRef]
- Saxena, B.N.; Refetoff, S.; Emerson, K., Jr.; Selenkow, H.A. A rapid radioimmunoassay for human placental lactogen. Application to normal and pathologic pregnancies. Am. J. Obstet. Gynecol. 1968, 101, 874–885. [Google Scholar] [CrossRef]
- Peeters, L.L.; Lemons, J.A.; Niswender, G.D.; Battaglia, F.C. Serum levels of human placental lactogen and human chorionic gonadotropin in early pregnancy: A maturational index of the placenta. Am. J. Obstet. Gynecol. 1976, 126, 707–711. [Google Scholar] [CrossRef]
- Moore, T.; Dveksler, G.S. Pregnancy-specific glycoproteins: Complex gene families regulating maternal-fetal interactions. Int. J. Dev. Biol. 2014, 58, 273–280. [Google Scholar] [CrossRef] [Green Version]
- Aleksic, D.; Blaschke, L.; Missbach, S.; Hanske, J.; Weiss, W.; Handler, J.; Zimmermann, W.; Cabrera-Sharp, V.; Read, J.E.; de Mestre, A.M.; et al. Convergent evolution of pregnancy-specific glycoproteins in human and horse. Reproduction 2016, 152, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Blois, S.M.; Sulkowski, G.; Tirado-Gonzalez, I.; Warren, J.; Freitag, N.; Klapp, B.F.; Rifkin, D.; Fuss, I.; Strober, W.; Dveksler, G.S. Pregnancy-specific glycoprotein 1 (PSG1) activates TGF-beta and prevents dextran sodium sulfate (DSS)-induced colitis in mice. Mucosal Immunol. 2014, 7, 348–358. [Google Scholar] [CrossRef] [Green Version]
- Streydio, C.; Vassart, G. Expression of human pregnancy specific beta 1 glycoprotein (PSG) genes during placental development. Biochem. Biophys. Res. Commun. 1990, 166, 1265–1273. [Google Scholar] [CrossRef]
- Grobman, W.A.; Rice, M.M.; Reddy, U.M.; Tita, A.T.N.; Silver, R.M.; Mallett, G.; Hill, K.; Thom, E.A.; El-Sayed, Y.Y.; Perez-Delboy, A.; et al. Labor Induction versus Expectant Management in Low-Risk Nulliparous Women. N. Engl. J. Med. 2018, 379, 513–523. [Google Scholar] [CrossRef]
- Hersh, A.R.; Skeith, A.E.; Sargent, J.A.; Caughey, A.B. Induction of labor at 39 weeks of gestation versus expectant management for low-risk nulliparous women: A cost-effectiveness analysis. Am. J. Obstet. Gynecol. 2019, 220, 590.e1–590.e10. [Google Scholar] [CrossRef]
- Committee on Obstetric Practice American Institute of Ultrasound in Medicine Society for Maternal–Fetal Medicine. Committee Opinion No 700: Methods for Estimating the Due Date. Obstet. Gynecol. 2017, 129, e150–e154. [Google Scholar] [CrossRef]
- Foster, D.G.; Gould, H.; Biggs, M.A. Timing of pregnancy discovery among women seeking abortion. Contraception 2021, 104, 642–647. [Google Scholar] [CrossRef]
- Flood-Nichols, S.K.; Tinnemore, D.; Wingerd, M.A.; Abu-Alya, A.I.; Napolitano, P.G.; Stallings, J.D.; Ippolito, D.L. Longitudinal analysis of maternal plasma apolipoproteins in pregnancy: A targeted proteomics approach. Mol. Cell. Proteom. 2013, 12, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of the Study Population | Mean (IQR) or % (n) * |
|---|---|
| Maternal Age (years) | 26.13 ± 1 (22–29) |
| Body mass index (BMI), kg/m2 | 25.21 ± 0.89 (24–30) |
| Parity | 0 |
| Race | |
| White (Caucasian) | 82.6% (19) |
| African American | 8.7% (2) |
| Asian | 4.3% (1) |
| Other | 4.3% (1) |
| Type of delivery | |
| Vaginal | 69.6% (16) |
| Cesarean | 30% (7) |
| Birthweight, Kg | 3.57 ± 0.14 (3.26–3.76) |
| Gestational age (weeks) | 39.72 ± 0.24 (39.14–40.75) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yohannes, E.; Ippolito, D.L.; Damicis, J.R.; Dornisch, E.M.; Leonard, K.M.; Napolitano, P.G.; Ieronimakis, N. Longitudinal Proteomic Analysis of Plasma across Healthy Pregnancies Reveals Indicators of Gestational Age. Int. J. Mol. Sci. 2022, 23, 7076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137076
Yohannes E, Ippolito DL, Damicis JR, Dornisch EM, Leonard KM, Napolitano PG, Ieronimakis N. Longitudinal Proteomic Analysis of Plasma across Healthy Pregnancies Reveals Indicators of Gestational Age. International Journal of Molecular Sciences. 2022; 23(13):7076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137076
Chicago/Turabian StyleYohannes, Elizabeth, Danielle L. Ippolito, Jennifer R. Damicis, Elisabeth M. Dornisch, Katherine M. Leonard, Peter G. Napolitano, and Nicholas Ieronimakis. 2022. "Longitudinal Proteomic Analysis of Plasma across Healthy Pregnancies Reveals Indicators of Gestational Age" International Journal of Molecular Sciences 23, no. 13: 7076. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137076