
Catalase Inhibition by Aminoalkanol Derivatives with Potential Anti-Cancer Activity—In Vitro and In Silico Studies Using Capillary Electrophoresis Method


Abstract
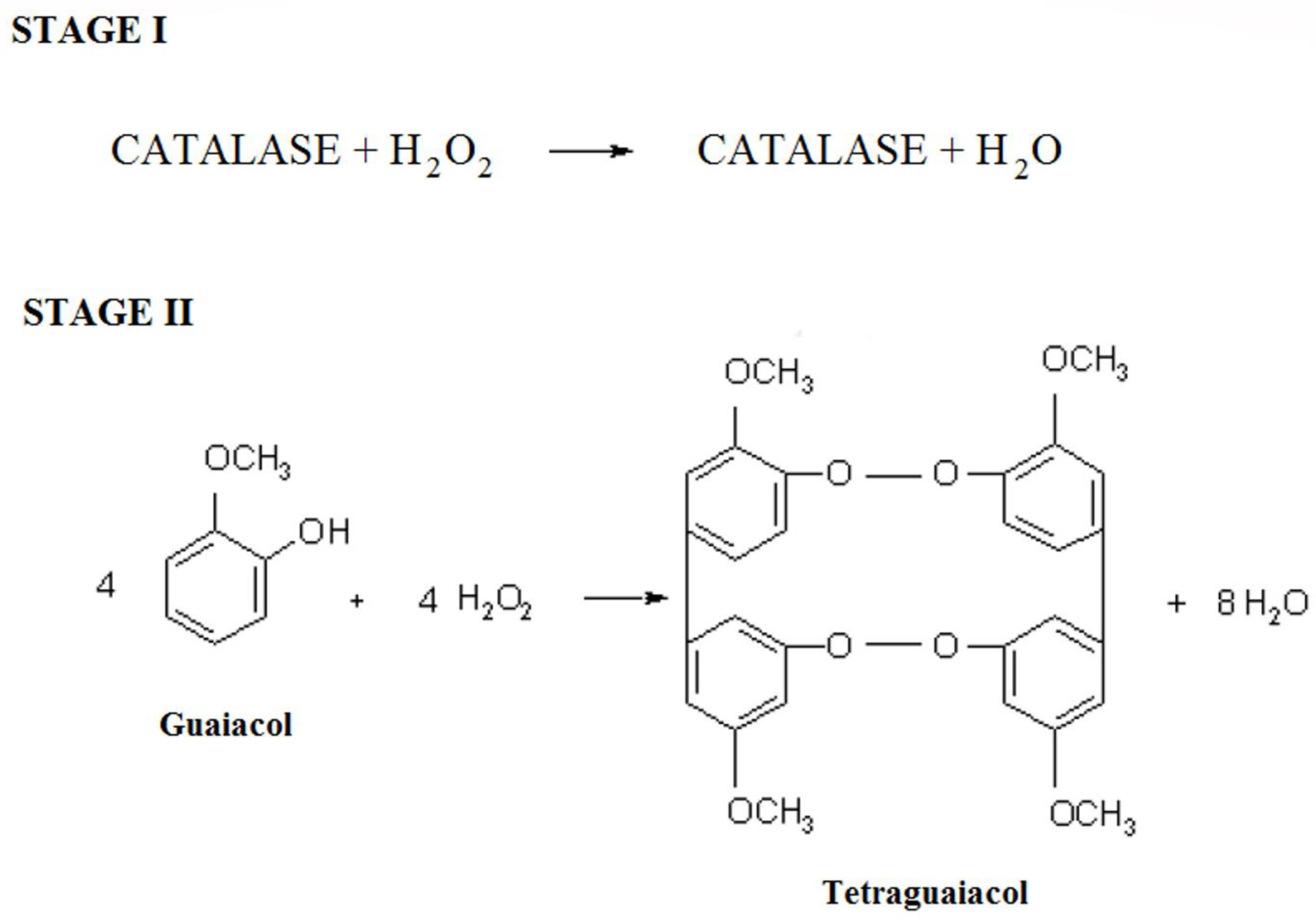
:1. Introduction
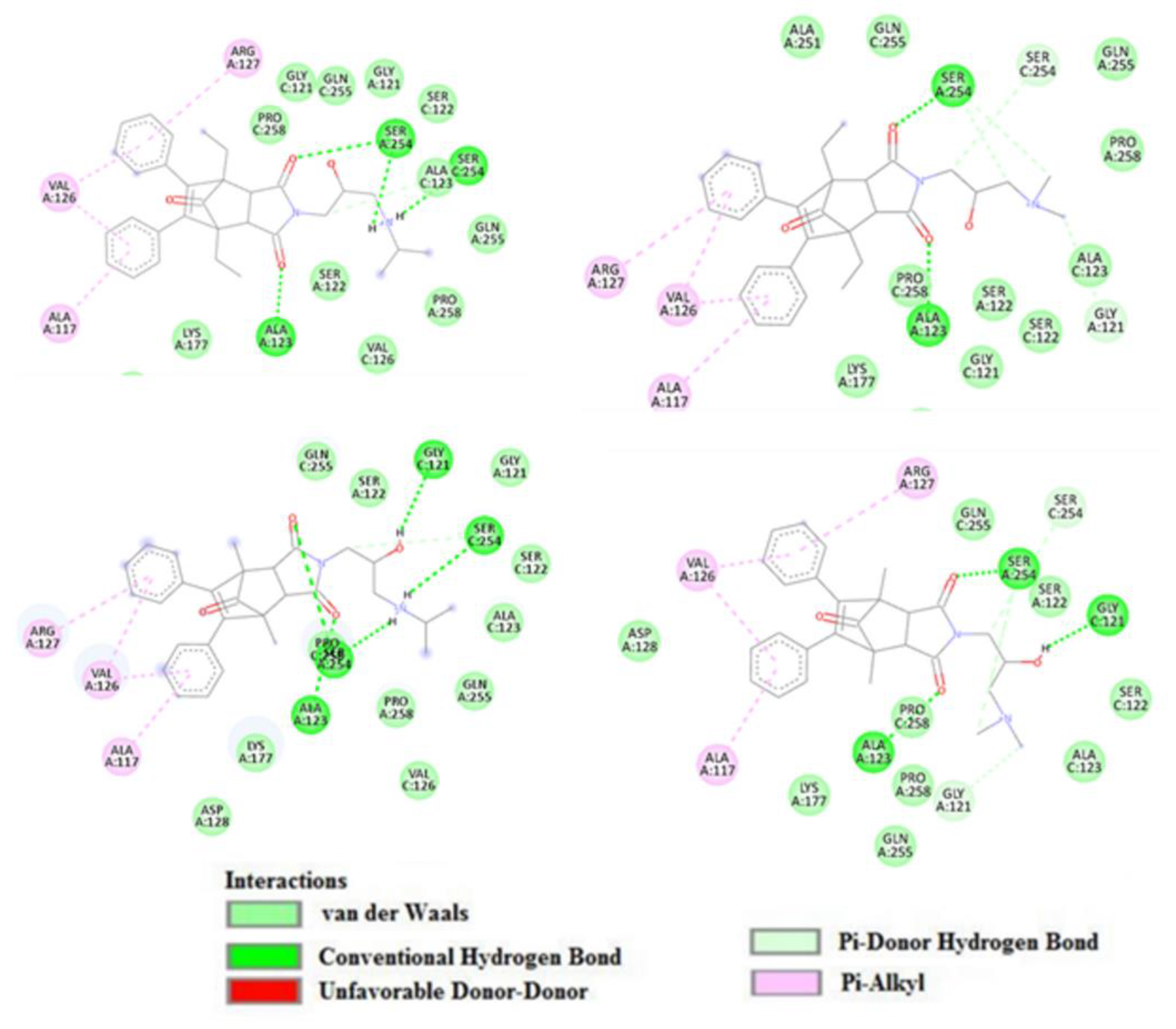
2. Results
2.1. Experimental
2.2. Analysis
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Instrumentation
4.3. Capillary Electrophoresis (CE) Conditions
4.4. Preparation of Stock and Working Standards
4.5. Sample Preparation
4.6. Docking Studies
5. Conclusions
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Von Ossowski, I.; Hausner, G.; Loewen, P.C. Molecular evolutionary analysis based on the amino acid sequence of catalase. J. Mol. Evol. 1993, 37, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Deisseroth, A.; Dounce, A.L. Catalase: Physical and chemical properties, mechanism of catalysis, and physiological role. Physiol. Rev. 1970, 50, 319–375. [Google Scholar] [CrossRef] [PubMed]
- Ivancich, A.; Jouve, H.M.; Sartor, B.; Gaillard, J. EPR investigation of compound I in Proteus mirabilis and bovine liver catalases: Formation of porphyrin and tyrosyl radical intermediates. Biochemistry 1997, 36, 9356–9364. [Google Scholar] [CrossRef] [PubMed]
- Lardinois, O.M. Reactions of bovine liver catalase with superoxide radicals and hydrogen peroxide. Free Radic Res. 1995, 22, 251–274. [Google Scholar] [CrossRef]
- Sumner, J.B.; Dounce, A.L. Crystalline catalase. J. Biochem. 1937, 121, 417–424. [Google Scholar]
- Herbert, D.; Pinsent, J. Crystalline bacterial catalase. J. Biochem. 1948, 43, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Putnam, C.D.; Arvai, A.S.; Bourne, Y.; Tainer, J.A. Active and inhibited human catalase structures: Ligand and NADPH binding and catalytic mechanism. J. Mol. Biol. 2000, 296, 295–309. [Google Scholar] [CrossRef]
- Gaetani, G.F.; Ferraris, A.M.; Roflo, M.; Mangerini, R.; Arena, S.; Kirkman, H.N. Predominant role of catalase in the disposal of hydrogen peroxide within human erythrocytes. Blood 1996, 87, 1595–1599. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.; Riedel, H.D.; Stremmel, W. Direct evidence for catalase as the predominant H2O2 removing enzyme in human erythrocytes. Blood 1997, 90, 4973–4978. [Google Scholar] [CrossRef]
- Majumder, D.; Das, A.; Saha, C. Catalase inhibition an anti cancer property of flavonoids: A kinetic and structural evaluation. Int. J. Biol. Macromol. 2017, 104, 929–935. [Google Scholar] [CrossRef]
- Ueda, M.; Kinoshita, H.; Yoshida, T.; Kamasawa, N.; Osumi, M.; Tanaka, A. Effect of catalase specific inhibitor 3-amino-1,2,4-triazole on yeast peroxisomal catalase in vivo. FEMS Microbiol Lett. 2003, 219, 93–98. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Liu, M.; Ai, Q.; Lin, L.; Wu, K.; Deng, X. Involvement of catalase in the protective benefits of metformin in mice with oxidative liver injury. Chem. Biol. Interact. 2014, 216, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Akbayirli, P.; Akyilmaz, E. Activation-Based Catalase Enzyme Electrode and its Usage for Glucose Determination. Anal. Lett. 2007, 40, 3360–3372. [Google Scholar] [CrossRef]
- Murali, A.; Patel, S. The Effect of Different Heavy Metal Acetate Solutions on the Inhibition of Catalase Enzyme. J. S. C. Acad. Sci. 2017, 15, 68–74. [Google Scholar]
- Atli, G.; Alptekin, O.; Tukel, S.; Canli, M. Response of catalase activity to Ag+, Cd2+, Cr6+,Cu2+ and Zn2+ in five tissues of freshwater fish Oreochromis niloticus. Comparative Biochemistry and Physiology. Toxicol. Pharmacol. 2006, 143, 218–224. [Google Scholar]
- Lee, H.I.; Yoon, J.H.; Nam, J.S.; Kim, Y.M.; Ro, Y.T. Cloning, expression and characterization of the catalase-peroxidase (KatG) gene from a fast-growing Mycobacterium sp. strain JC1DSM 3803. J. Biochem. 2010, 147, 511–522. [Google Scholar] [CrossRef]
- Kengen, S.W.; Bikker, F.J.; Hagen, W.R.; de Vos, W.M.; van der Oost, J. Characterization of a catalase-peroxidase from the hyperthermophilic archaeon Archaeoglobus fulgidus. Extremophiles 2001, 5, 323–332. [Google Scholar] [CrossRef]
- Darr, D.; Fridovich, I. Inhibition of catalase by 3,3’-diaminobenzidine. Biochem. J. 1985, 226, 781–787. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.; Hu, Z.L.; Zhang, L.; Zhao, Y.S.; Meng, Q.H.; Guan, Q.B. Association of catalase genotype with oxidative stress in the predication of colorectal cancer: Modification by epidemiological factors. Biomed. Environ. Sci. 2012, 25, 156–162. [Google Scholar]
- Monari, M.; Foschi, J.; Calabrese, C.; Liguori, G.; Di Febo, G.; Rizzello, F. Implications of antioxidant enzymes in human gastric neoplasms. Int. J. Mol. Med. 2009, 24, 693–700. [Google Scholar] [CrossRef] [Green Version]
- Iborra, M.; Moret, I.; Rausell, F.; Bastida, G.; Aguas, M.; Cerrillo, E. Role of oxidative stress and antioxidant enzymes in Crohn’s disease. Biochem. Soc. Trans. 2011, 39, 1102–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Góth, L. Two cases of acatalasemia in Hungary. Clin. Chim Acta 1992, 207, 155–158. [Google Scholar] [CrossRef]
- Al-Abrash, A.S.; Al-Quobaili, F.A.; Al-Akhras, G.N. Catalase evaluation in different human diseases associated with oxidative stress. Saudi J. Med. 2000, 21, 826–830. [Google Scholar]
- Habib, L.K.; Lee, M.T.C.; Yang, J. Inhibitors of catalase-amyloid interactions protect cells from β-amyloid induced oxidative stress and toxicity. J. Biol. Chem. 2010, 285, 38933–38943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, M.; Suzuki, H.; Suzuki, M.; Kai, A.; Miura, S.; Ishii, H. Catalase and superoxide dismutase secreted from Helicobacter pylori. Helicobacter 1997, 2, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Taylor, W.I.; Achanzar, D. Catalase test as an aid to the identification of Enterobacteriaceae. Appl. Microbiol. 1972, 24, 58–61. [Google Scholar] [CrossRef] [PubMed]
- Karakus, Y.Y. Typical Catalases: Function and Structure; Intech Open: London, UK, 2020; Chapter 7; pp. 1–16. [Google Scholar] [CrossRef] [Green Version]
- Nicholls, P.; Fita, I.; Loewen, P.C. Enzymology and structure of catalases. Adv. Inorg. Chem. 2001, 51, 51–106. [Google Scholar]
- Kuran, B.; Krawiecka, M.; Kossakowski, J.; Cieślak, M.; Kaźmierczak-Barańska, J.; Królewska, K.; Nawrot, B. Synthesis and Biological Activity of Novel Series of Heterocyclic Compounds Containing Succinimide Moiety. Patent Application P-400000 (PL), 13 July 2013. [Google Scholar]
- Grodner, B.; Napiórkowska, M. Characterization and inhibition studies of tissue nonspecific alkaline phosphatase by aminoalkanol derivatives of 1,7-dimethyl-8,9-diphenyl-4-azatricyclo[5.2.1.02,6]dec-8-ene-3,5,10-trione, new competitive and non-competitive inhibitors, by capillary electrophoresis. J. Pharm. Biomed. Anal. 2017, 143, 285–290. [Google Scholar]
- Grodner, B.; Napiórkowska, M.; Pisklak, D.M. Kinetic Studies of Newly Patented Aminoalkanol Derivatives with Potential Anticancer Activity as Competitive Inhibitors of Prostate Acid Phosphatase. Int. J. Mol. Sci. 2021, 22, 11761. [Google Scholar] [CrossRef]
- Grodner, B.; Napiórkowska, M.; Pisklak, D.M. In Vitro and In Silico Kinetic Studies of Patented 1,7-diEthyl and 1,7-diMethyl Aminoalkanol Derivatives as New Inhibitors of Acetylcholinesterase. Int. J. Mol. Sci. 2022, 23, 270. [Google Scholar] [CrossRef]
- Grodner, B.; Napiórkowska, M. Capillary electrophoresis for the investigation of two novel aminoalkanol derivatives of 1,7-diethyl-8,9-diphenyl-4-azatricyclo[5.2.1.02,6] dec-8-ene-3,5,10-trione as potential anticancer drugs in water solution and serum. J. Sep. Sci. 2020, 43, 648–656. [Google Scholar] [CrossRef] [PubMed]
- Grodner, B.; Napiórkowska, M. Dual 2-Hydroxypropyl-β-Cyclodextrin and 5,10,15,20-Tetrakis (4-Hydroxyphenyl) Porphyrin System as a Novel Chiral-Achiral Selector Complex for Enantioseparation of Aminoalkanol Derivatives with Anticancer Activity in Capillary Electrophoresis. Molecules 2021, 26, 993. [Google Scholar] [CrossRef] [PubMed]
- Grodner, B.; Łukaszkiewicz, J.; Krawiecka, M. Capillary electrophoresis separation of aminoalkanol derivatives of 1,7-dimethyl-8,9-diphenyl-4-azatricyclo[5.2.1.02,6]dec-8-ene-3,5,10-trione as potential anticancer drugs. J. Sep. Sci. 2014, 37, 3564–3570. [Google Scholar] [CrossRef] [PubMed]
- Grodner, B.; Łukaszkiewicz, J.; Napiórkowska, M. Determination of the stereoisomers in aqueous medium and serum and validation studies of racemic aminoalkanol derivatives of 1,7-dimethyl-8,9-diphenyl-4-azatricyclo[5.2.1.02,6]dec-8-ene-3,5,10-trione, potential new anticancer drugs, by capillary electrophoresis. J. Sep. Sci. 2016, 39, 3246–3253. [Google Scholar]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comp. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Linearity Range of Product (Tetraguaiacol) (mM) | R2 | RSD (%) | LOD (mM) | LOQ (mM) | Regression Equation | Standard | Deviation |
|---|---|---|---|---|---|---|---|
| Slope | Intercept | ||||||
| 0.05–20.00 | 0.9999 | 2.38 | 0.01 | 0.04 | y = 10.351 x − 1.232 | ±0.67 | ±0.058 |
| Linearity Range of Substrate 2 (G uaiacol) (mM) | R2 | RSD (%) | LOD (mM) | LOQ (mM) | Regression Equation | Standard | Deviation |
| Slope | Intercept | ||||||
| 0.05–20.00 | 0.9999 | 3.14 | 0.02 | 0.07 | y = 9.979 x + 1.695 | ±0.83 | ±0.067 |
| Concentration Inhibitors (I), (II), (III) and (IV) (mM) | Linearity Range of Substrate (Hydrogen Peroxide) (mM) | R2 | RSD (%) | Regression Equation | Standard Deviation | |
|---|---|---|---|---|---|---|
| Slope | Intercept | |||||
| 0 | 0.08–10.00 | 0.9989 | 2.74 | y = 0.4308 x + 0.7933 | ±0.0047 | ±0.0036 |
| (I) 20.00 | 0.08–10.00 | 0.9957 | 3.27 | y = 0.5950 x + 0.8182 | ±0.0072 | ±0.0047 |
| (II) 20.00 | 0.08–10.00 | 0.9979 | 3.01 | y = 0.5109 x + 0.7981 | ±0.0053 | ±0.0042 |
| (III) 20.00 | 0.08–10.00 | 0.9949 | 3.96 | y = 0.6414 x + 0.8375 | ±0.0076 | ±0.0053 |
| (IV) 20.00 | 0.08–10.00 | 0.9978 | 2.93 | y = 0.5605 x + 0.7918 | ±0.0052 | ±0.0039 |
| Control System (Without Inhibitor) | System with Inhibitor (I) | System with Inhibitor (II) | System with Inhibitor (III) | System with Inhibitor (IV) | |
|---|---|---|---|---|---|
| Km | 0.54 | 0.73 | 0.64 | 0.77 | 0.71 |
| Vmax | 1.26 | 1.25 | 1.24 | 1.25 | 1.26 |
| Type of inhibition | Competitive | Competitive | Competitive | Competitive |
| Compound (I) | ||||
|---|---|---|---|---|
| Concentration (mM) | Straight Line Equation | R2 | Slope | |
| (I) 0.00 | y = 0.4308 x + 0.7933 | 0.9997 ± 0.0008 | 23.31 ± 0.04 | |
| (I) 2.50 | y = 0.4604 x + 0.8103 | 0.9988 ± 0.0011 | 24.72 ± 0.05 | |
| (I) 5.00 | y = 0.4754 x + 0.8293 | 0.9995 ± 0.0009 | 25.43 ± 0.05 | |
| (I) 10.00 | y = 0.5167 x + 0.8259 | 0.9994 ± 0.0009 | 27.33 ± 0.06 | |
| (I) 15.00 | y = 0.54575 x + 0.8388 | 0.9996 ± 0.0009 | 28.62 ± 0.06 | |
| (I) 20.00 | y = 0.5950 x + 0.8192 | 0.9998 ± 0.0008 | 30.75 ± 0.04 | |
| Concentration (mM) | Km (mM) | Vmax (mM/min) | Ki (mM) | IC50 (mM) |
| (I) 0.00 | 0.54 ± 0.03 | 1.26 ± 0.02 | ||
| (I) 2.50 | 0.56 ± 0.04 | 1.25 ± 0.04 | 11.73 ± 0.05 | |
| (I) 5.00 | 0.57 ± 0.04 | 1.24 ± 0.07 | 9.76 ± 0.05 | |
| (I) 10.00 | 0.62 ± 0.05 | 1.25 ± 0.05 | 3.57 ± 0.04 | 11.40 ± 0.05 |
| (I) 15.00 | 0.65 ± 0.04 | 1.24 ± 0.07 | 2.74 ± 0.04 | |
| (I) 20.00 | 0.73 ± 0.03 | 1.25 ± 0.05 | 1.61 ± 0.03 | |
| Compound (II) | ||||
| Concentration (mM) | Straight Line Equation | R2 | Slope | |
| (II) 0.00 | y = 0.4308 x + 0.7933 | 0.9997 ± 0.0008 | 23.31 ± 0.04 | |
| (II) 2.50 | y = 0.4390 x + 0.7993 | 0.9992 ± 0.0015 | 23.70 ± 0.05 | |
| (II) 5.00 | y = 0.4503 x + 0.8151 | 0.9996 ± 0.0009 | 24.24 ± 0.05 | |
| (II) 10.00 | y = 0.4727 x + 0.7936 | 0.9996 ± 0.0010 | 25.30 ± 0.06 | |
| (II) 15.00 | y = 0.4874 x + 0.7985 | 0.9995 ± 0.0011 | 26.00 ± 0.04 | |
| (II) 20.00 | y = 0.5109 x + 0.7982 | 0.9998 ± 0.0008 | 27.06 ± 0.05 | |
| Concentration (mM) | Km (mM) | Vmax (mM/min) | Ki (mM) | IC50 (mM) |
| (II) 0.00 | 0.54 ± 0.03 | 1.26 ± 0.02 | ||
| (II) 2.50 | 0.55 ± 0.05 | 1.24 ± 0.06 | 47.70 ± 0.08 | |
| (II) 5.00 | 0.56 ± 0.05 | 1.24 ± 0.06 | 18.13 ± 0.06 | |
| (II) 10.00 | 0.59 ± 0.04 | 1.26 ± 0.03 | 5.61 ± 0.05 | 12.80 ± 0.07 |
| (II) 15.00 | 0.61 ± 0.03 | 1.25 ± 0.04 | 4.38 ± 0.05 | |
| (II) 20.00 | 0.64 ± 0.03 | 1.24 ± 0.04 | 3.04 ± 0.03 | |
| Compound (III) | ||||
| Concentration (mM) | Straight Line Equation | R2 | Slope | |
| (III) 0.00 | y = 0.4308 x + 0.7933 | 0.9997 ± 0.0008 | 23.31 ± 0.04 | |
| (III) 2.50 | y = 0.4682 x + 0.8124 | 0.9996 ± 0.0013 | 25.09 ± 0.04 | |
| (III) 5.00 | y = 0.4843 x + 0.8395 | 0.9998 ± 0.0006 | 25.84 ± 0.05 | |
| (III) 10.00 | y = 0.5452 x + 0.8396 | 0.9997 ± 0.0009 | 28.60 ± 0.04 | |
| (III) 15.00 | y = 0.5976 x + 0.8449 | 0.9997 ± 0.0010 | 29.58 ± 0.03 | |
| (III) 20.00 | y = 0.6414 x + 0.8375 | 0.9998 ± 0.0007 | 32.68 ± 0.02 | |
| Concentration (mM) | Km (mM) | Vmax (mM/min) | Ki (mM) | IC50 (mM) |
| (III) 0.00 | 0.54 ± 0.03 | 1.26 ± 0.02 | ||
| (III) 2.50 | 0.57 ± 0.04 | 1.24 ± 0.07 | 8.86 ± 0.05 | |
| (III) 5.00 | 0.58 ± 0.03 | 1.25 ± 0.08 | 8.71 ± 0.05 | |
| (III) 10.00 | 0.65 ± 0.04 | 1.24 ± 0.07 | 2.77 ± 0.04 | 10.30 ± 0.03 |
| (III) 15.00 | 0.71 ± 0.02 | 1.24 ± 0.09 | 1.80 ± 0.05 | |
| (III) 20.00 | 0.77 ± 0.02 | 1.25 ± 0.02 | 1.32 ± 0.02 | |
| Compound (IV) | ||||
| Concentration (mM) | Straight Line Equation | R2 | Slope | |
| (IV) 0.00 | y = 0.4308 x + 0.7933 | 0.9997 ± 0.0008 | 23.31 ± 0.04 | |
| (IV) 2.50 | y = 0.4522 x + 0.8021 | 0.9995 ± 0.0010 | 24.33 ± 0.06 | |
| (IV) 5.00 | y = 0.4662 x + 0.8167 | 0.9995 ± 0.0009 | 25.00 ± 0.05 | |
| (IV) 10.00 | y = 0.4930 x + 0.8052 | 0.9993 ± 0.0018 | 26.24 ± 0.04 | |
| (IV) 15.00 | y = 0.5162 x + 0.8146 | 0.9996 ± 0.0009 | 27.30 ± 0.05 | |
| (IV) 20.00 | y = 0.5605 x + 0.7918 | 0.9998 ± 0.0008 | 29.27 ± 0.03 | |
| Concentration (mM) | Km (mM) | Vmax (mM/min) | Ki (mM) | IC50 (mM) |
| (IV) 0.00 | 0.54 ± 0.03 | 1.26 ± 0.02 | ||
| (IV) 2.50 | 0.56 ± 0.04 | 1.25 ± 0.03 | 14.23 ± 0.06 | |
| (IV) 5.00 | 0.57 ± 0.05 | 1.24 ± 0.06 | 10.61 ± 0.04 | |
| (IV) 10.00 | 0.61 ± 0.05 | 1.24 ± 0.06 | 4.26 ± 0.04 | 12.20 ± 0.06 |
| (IV) 15.00 | 0.63 ± 0.03 | 1.23 ± 0.05 | 3.25 ± 0.04 | |
| (IV) 20.00 | 0.71 ± 0.03 | 1.26 ± 0.02 | 1.79 ± 0.04 | |
| Compound | R1 | R2 | R3 | Ki [mM] | IC50 [mM] | Docking Energy [kcal/mol] |
|---|---|---|---|---|---|---|
| Derivative (I) |  |  |  | 1.61 | 11.40 | −8.6 |
| Derivative (II) |  |  |  | 3.04 | 12.80 | −8.4 |
| Derivative (III) |  |  |  | 1.32 | 10.30 | −8.9 |
| Derivative (IV) |  |  |  | 1.79 | 12.20 | −8.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Grodner, B.; Napiórkowska, M.; Pisklak, D.M. Catalase Inhibition by Aminoalkanol Derivatives with Potential Anti-Cancer Activity—In Vitro and In Silico Studies Using Capillary Electrophoresis Method. Int. J. Mol. Sci. 2022, 23, 7123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137123
Grodner B, Napiórkowska M, Pisklak DM. Catalase Inhibition by Aminoalkanol Derivatives with Potential Anti-Cancer Activity—In Vitro and In Silico Studies Using Capillary Electrophoresis Method. International Journal of Molecular Sciences. 2022; 23(13):7123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137123
Chicago/Turabian StyleGrodner, Błażej, Mariola Napiórkowska, and Dariusz Maciej Pisklak. 2022. "Catalase Inhibition by Aminoalkanol Derivatives with Potential Anti-Cancer Activity—In Vitro and In Silico Studies Using Capillary Electrophoresis Method" International Journal of Molecular Sciences 23, no. 13: 7123. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137123