
Heat Shock Alters the Proteomic Profile of Equine Mesenchymal Stem Cells

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
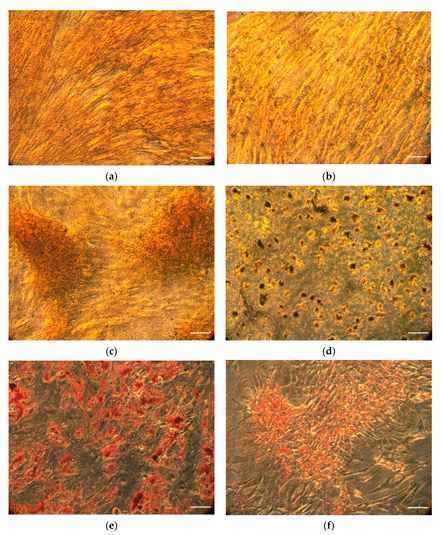
2.1. Isolation and Differentiation of Stem Cells
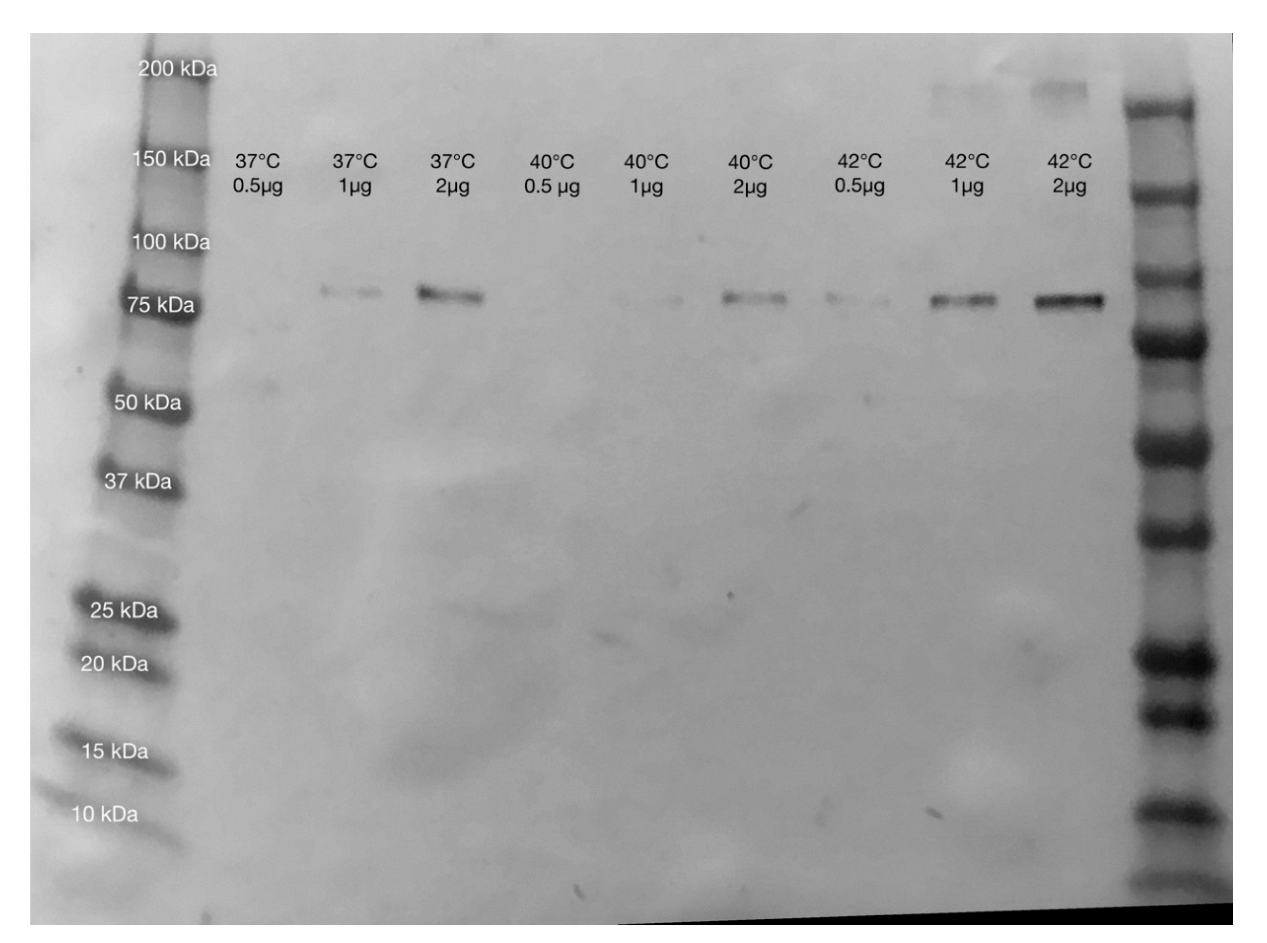
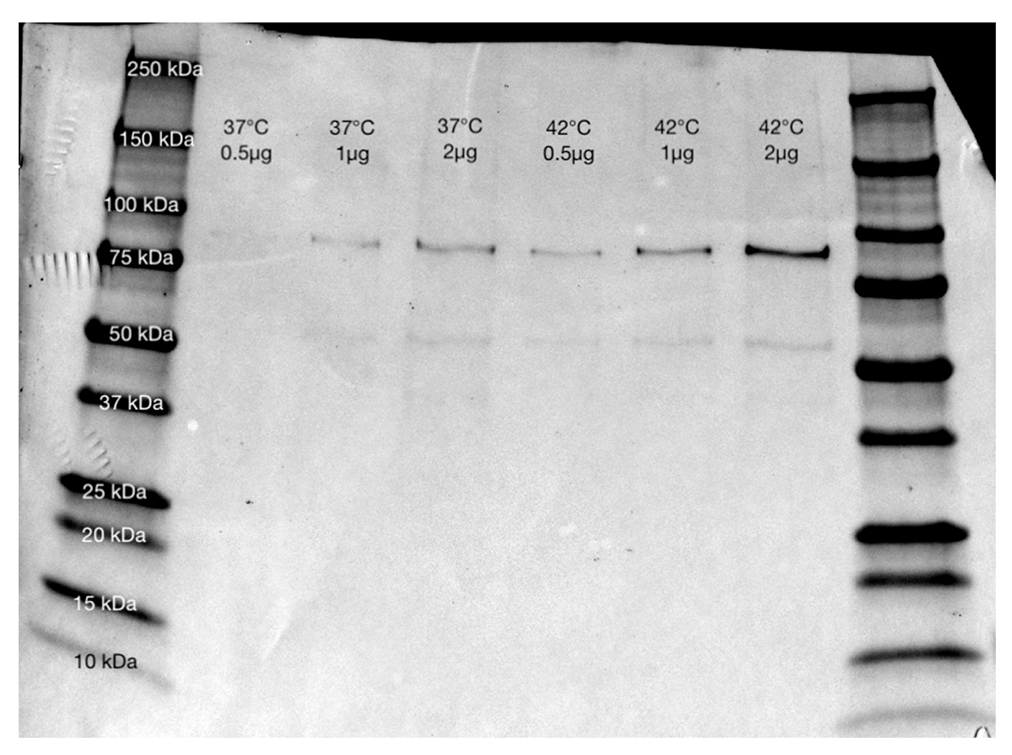
2.2. Heat Stress Conditions
2.3. Proteomic Analysis using Mass Spectrometry
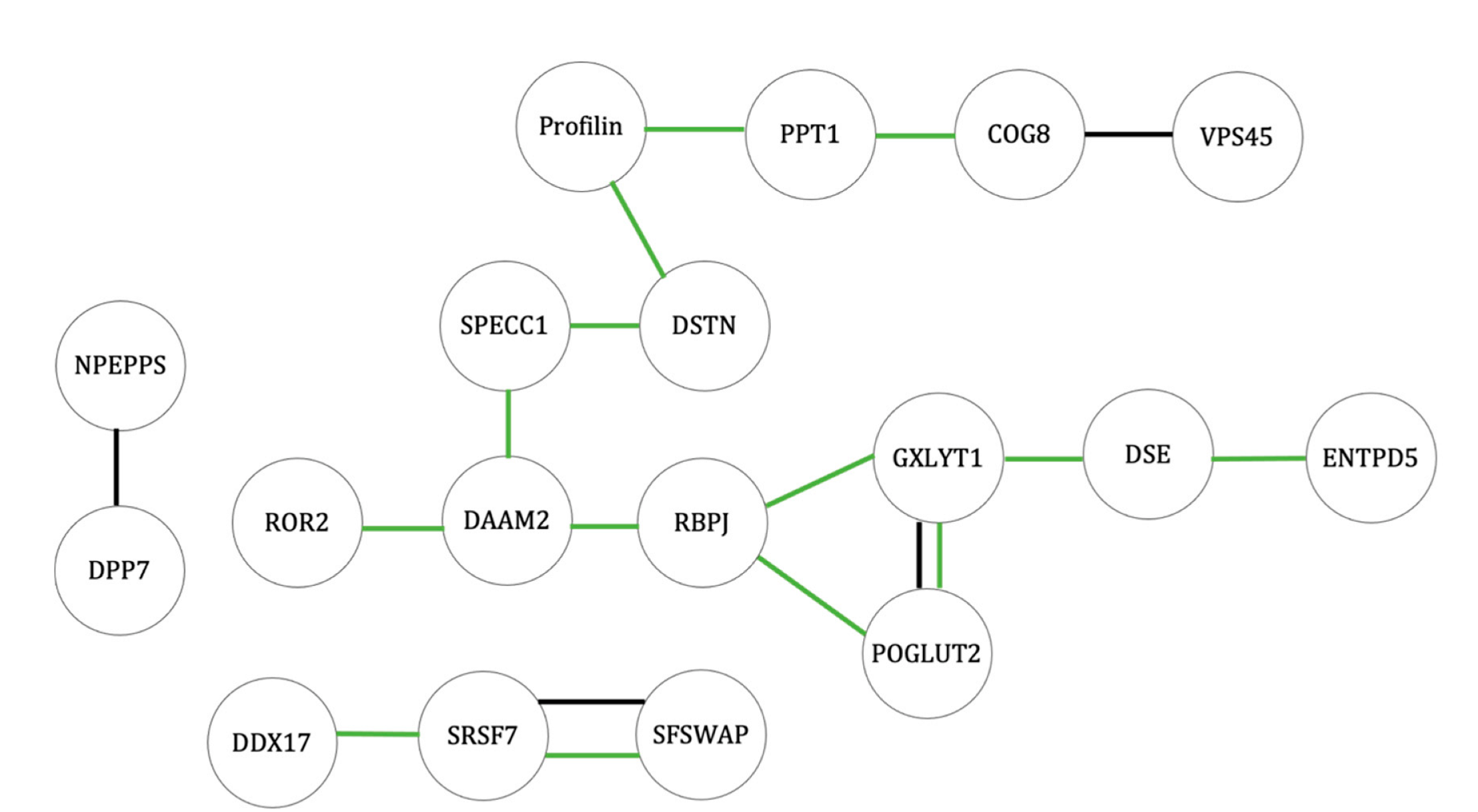
2.3.1. Altered Protein Levels of Osteoblasts at 42 °C Compared with Control Cell Population at 37 °C
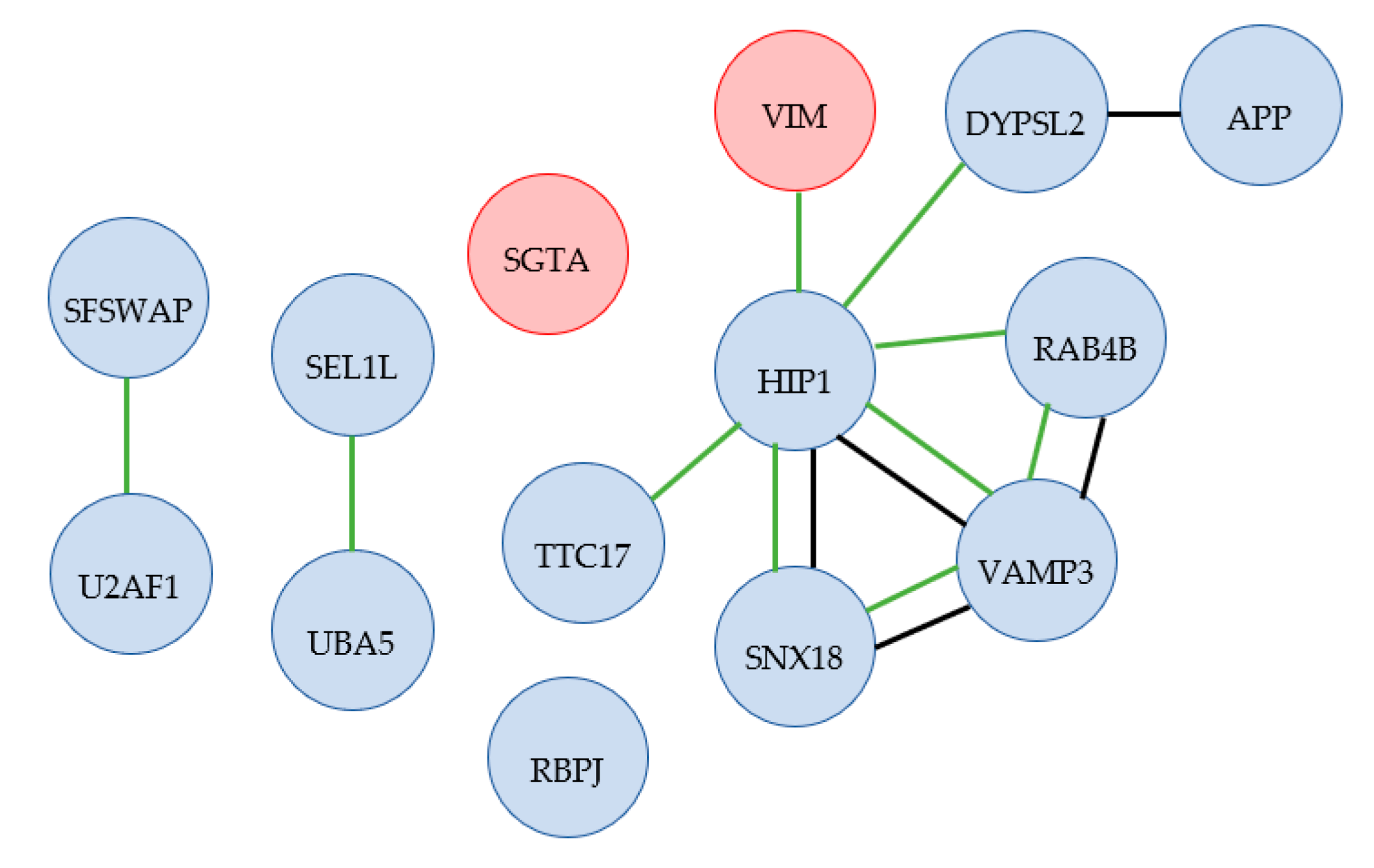
2.3.2. Altered Protein Levels of Adipocytes at 42 °C Compared with Control Cell Population at 37 °C
3. Discussion
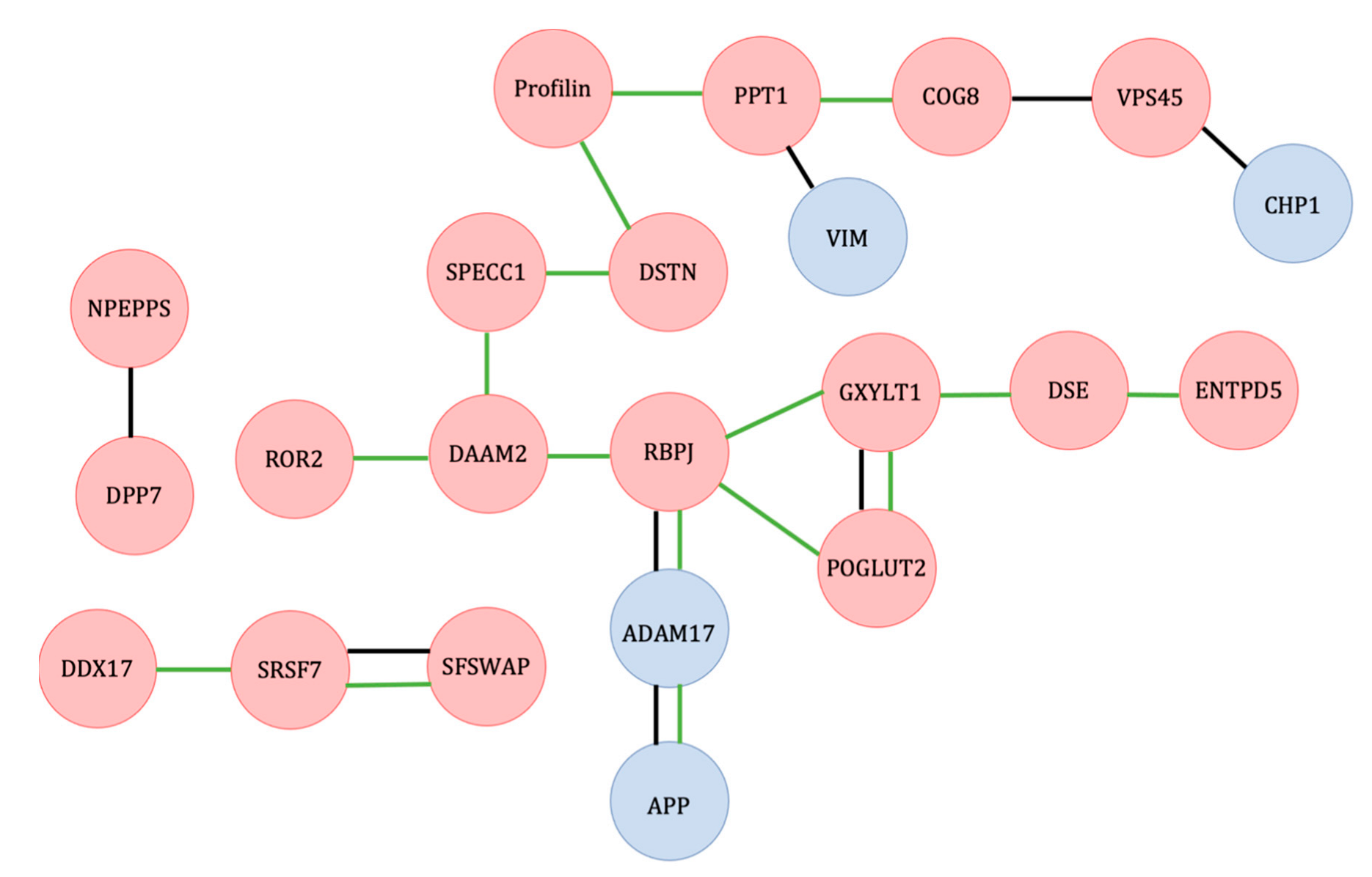
3.1. Wnt and Notch Signaling Balance and Their Effects on MSC Differentiation
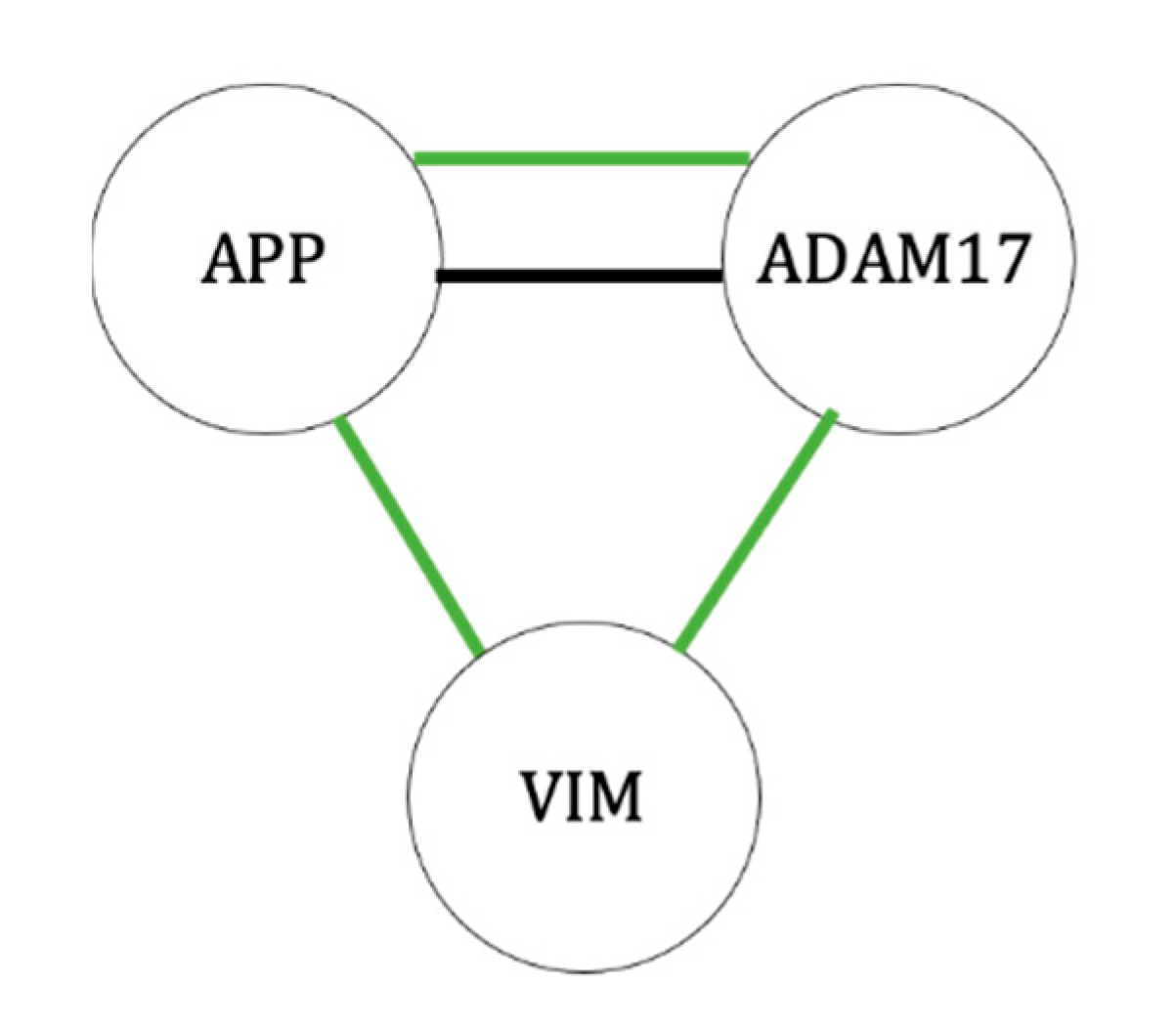
3.2. Cytoskeletal Proteins Necessary for Differentiated Cell Morphology and Cell Signaling
3.3. mRNA Transcription and Splicing Regulation of Differentiation and Stem Cell Proliferation
3.4. Endocytosis-Related Proteins in Adipogenic Differentiation
3.5. Endoplasmic Reticulum-Associated Degradation Affecting Adipocyte Differentiation
3.6. Potential New Protein Targets
3.7. Overall Effects
3.8. Impact of Heat Stress on Cell Development
4. Materials and Methods
4.1. Isolation and Differentiation of Mesenchymal Stem Cells
4.2. Heat Stress Conditions
4.3. Western Blot
4.4. Proteomic Analysis Using Liquid Chromatography/Tandem Mass Spectrometry (LC-MS/MS)
4.4.1. A Chloroform/Methanol Precipitation Protocol
4.4.2. Reduction and Digestion
4.4.3. Tandem Mass Tag (TMT) Labeling
4.4.4. Reverse-Phase Liquid Chromatography
4.4.5. Proteomics Analysis with LC-MS/MS
4.5. Proteomic Data Analysis
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mazdiyasni, O.; Sadegh, M.; Chiang, F.; AghaKouchak, A. Heat Wave Intensity Duration Frequency Curve: A Multivariate Approach for Hazard and Attribution Analysis. Sci. Rep. 2019, 9, 14117. [Google Scholar] [CrossRef] [PubMed]
- Atkins, A.R.; Wyndham, C.H. A Study of Temperature Regulation in the Human Body with the Aid of an Analogue Computer. Pflug. Arch. 1969, 307, 104–119. [Google Scholar] [CrossRef]
- Piil, J.F.; Christiansen, L.; Morris, N.B.; Mikkelsen, C.J.; Ioannou, L.G.; Flouris, A.D.; Lundbye-Jensen, J.; Nybo, L. Direct Exposure of the Head to Solar Heat Radiation Impairs Motor-Cognitive Performance. Sci. Rep. 2020, 10, 7812. [Google Scholar] [CrossRef] [PubMed]
- Epstein, Y.; Yanovich, R. Heatstroke. N. Engl. J. Med. 2019, 380, 2449–2459. [Google Scholar] [CrossRef]
- Wells, J.C.K. Thermal Environment and Human Birth Weight. J. Theor. Biol. 2002, 214, 413–425. [Google Scholar] [CrossRef] [Green Version]
- Kuehn, L.; McCormick, S. Heat Exposure and Maternal Health in the Face of Climate Change. Int. J. Environ. Res. Public Health 2017, 14, 853. [Google Scholar] [CrossRef] [Green Version]
- De Boo, H.A.; Harding, J.E. The Developmental Origins of Adult Disease (Barker) Hypothesis. Aust. N. Z. J. Obstet. Gynaecol. 2006, 46, 4–14. [Google Scholar] [CrossRef]
- Cao-Lei, L.; Dancause, K.N.; Elgbeili, G.; Massart, R.; Szyf, M.; Liu, A.; Laplante, D.P.; King, S. DNA Methylation Mediates the Impact of Exposure to Prenatal Maternal Stress on BMI and Central Adiposity in Children at Age 13½ Years: Project Ice Storm. Epigenetics 2015, 10, 749–761. [Google Scholar] [CrossRef]
- Laplante, D.P.; Hart, K.J.; O’Hara, M.W.; Brunet, A.; King, S. Prenatal Maternal Stress Is Associated with Toddler Cognitive Functioning: The Iowa Flood Study. Early Hum. Dev. 2018, 116, 84–92. [Google Scholar] [CrossRef]
- Paquin, V.; Elgbeili, G.; Laplante, D.P.; Kildea, S.; King, S. Positive Cognitive Appraisal “Buffers” the Long-Term Effect of Peritraumatic Distress on Maternal Anxiety: The Queensland Flood Study. J. Affect. Disord. 2021, 278, 5–12. [Google Scholar] [CrossRef]
- Uccelli, A.; Moretta, L.; Pistoia, V. Mesenchymal Stem Cells in Health and Disease. Nat. Rev. Immunol. 2008, 8, 726–736. [Google Scholar] [CrossRef]
- Radtke, C.L.; Nino-Fong, R.; Esparza Gonzalez, B.P.; Stryhn, H.; McDuffee, L.A. Characterization and Osteogenic Potential of Equine Muscle Tissue- and Periosteal Tissue-Derived Mesenchymal Stem Cells in Comparison with Bone Marrow- and Adipose Tissue-Derived Mesenchymal Stem Cells. Am. J. Vet. Res. 2013, 74, 790–800. [Google Scholar] [CrossRef]
- Kanehisa, M.; Sato, Y.; Kawashima, M.; Furumichi, M.; Tanabe, M. KEGG as a Reference Resource for Gene and Protein Annotation. Nucleic Acids Res. 2016, 44, D457–D462. [Google Scholar] [CrossRef] [Green Version]
- UniProt Consortium, T. UniProt: The Universal Protein Knowledgebase. Nucleic Acids Res. 2018, 46, 2699. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Gable, A.L.; Lyon, D.; Junge, A.; Wyder, S.; Huerta-Cepas, J.; Simonovic, M.; Doncheva, N.T.; Morris, J.H.; Bork, P.; et al. STRING V11: Protein-Protein Association Networks with Increased Coverage, Supporting Functional Discovery in Genome-Wide Experimental Datasets. Nucleic Acids Res. 2019, 47, D607–D613. [Google Scholar] [CrossRef] [Green Version]
- Shimoni, C.; Goldstein, M.; Ribarski-Chorev, I.; Schauten, I.; Nir, D.; Strauss, C.; Schlesinger, S. Heat Shock Alters Mesenchymal Stem Cell Identity and Induces Premature Senescence. Front. Cell Dev. Biol. 2020, 8, 956. [Google Scholar] [CrossRef]
- de Magalhães, J.P.; Passos, J.F. Stress, Cell Senescence and Organismal Ageing. Mech. Ageing Dev. 2018, 170, 2–9. [Google Scholar] [CrossRef]
- Takam Kamga, P.; Bazzoni, R.; Dal Collo, G.; Cassaro, A.; Tanasi, I.; Russignan, A.; Tecchio, C.; Krampera, M. The Role of Notch and Wnt Signaling in MSC Communication in Normal and Leukemic Bone Marrow Niche. Front. Cell Dev. Biol. 2021, 8, 599276. [Google Scholar] [CrossRef]
- Zanotti, S.; Smerdel-Ramoya, A.; Stadmeyer, L.; Durant, D.; Radtke, F.; Canalis, E. Notch Inhibits Osteoblast Differentiation and Causes Osteopenia. Endocrinology 2008, 149, 3890–3899. [Google Scholar] [CrossRef] [Green Version]
- Regan, J.; Long, F. Notch Signaling and Bone Remodeling. Curr. Osteoporos. Rep. 2013, 11, 126–129. [Google Scholar] [CrossRef] [Green Version]
- Siebel, C.; Lendahl, U. Notch Signaling in Development, Tissue Homeostasis, and Disease. Physiol. Rev. 2017, 97, 1235–1294. [Google Scholar] [CrossRef] [Green Version]
- Tao, J.; Chen, S.; Yang, T.; Dawson, B.; Munivez, E.; Bertin, T.; Lee, B. Osteosclerosis Owing to Notch Gain of Function Is Solely Rbpj-dependent. J. Bone Min. Res. 2010, 25, 2175–2183. [Google Scholar] [CrossRef]
- Wang, S.; Kawashima, N.; Sakamoto, K.; Katsube, K.; Umezawa, A.; Suda, H. Osteogenic Differentiation of Mouse Mesenchymal Progenitor Cell, Kusa-A1 Is Promoted by Mammalian Transcriptional Repressor Rbpj. Biochem. Biophys. Res. Commun. 2010, 400, 39–45. [Google Scholar] [CrossRef]
- Thomas, S.; Jaganathan, B.G. Signaling Network Regulating Osteogenesis in Mesenchymal Stem Cells. J. Cell Commun. Signal. 2021, 16, 46–61. [Google Scholar] [CrossRef]
- Takeuchi, H.; Schneider, M.; Williamson, D.B.; Ito, A.; Takeuchi, M.; Handford, P.A.; Haltiwanger, R.S. Two Novel Protein O-Glucosyltransferases That Modify Sites Distinct from POGLUT1 and Affect Notch Trafficking and Signaling. Proc. Natl. Acad. Sci. USA 2018, 115, E8395–E8402. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, H.; Haltiwanger, R.S. Significance of Glycosylation in Notch Signaling. Biochem. Biophys. Res. Commun. 2014, 453, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.V.; Sethi, M.K.; Leonardi, J.; Rana, N.A.; Buettner, F.F.R.; Haltiwanger, R.S.; Bakker, H.; Jafar-Nejad, H. Negative Regulation of Notch Signaling by Xylose. PLoS Genet. 2013, 9, e1003547. [Google Scholar] [CrossRef] [Green Version]
- Müller, T.; Mizumoto, S.; Suresh, I.; Komatsu, Y.; Vodopiutz, J.; Dundar, M.; Straub, V.; Lingenhel, A.; Melmer, A.; Lechner, S.; et al. Loss of Dermatan Sulfate Epimerase (DSE) Function Results in Musculocontractural Ehlers-Danlos Syndrome. Hum. Mol. Genet. 2013, 22, 3761–3772. [Google Scholar] [CrossRef] [Green Version]
- Paganini, C.; Costantini, R.; Superti-Furga, A.; Rossi, A. Bone and Connective Tissue Disorders Caused by Defects in Glycosaminoglycan Biosynthesis: A Panoramic View. FEBS J. 2019, 286, 3008–3032. [Google Scholar] [CrossRef] [Green Version]
- Read, R.; Hansen, G.; Kramer, J.; Finch, R.; Li, L.; Vogel, P. Ectonucleoside Triphosphate Diphosphohydrolase Type 5 (Entpd5)-Deficient Mice Develop Progressive Hepatopathy, Hepatocellular Tumors, and Spermatogenic Arrest. Vet. Pathol. 2009, 46, 491–504. [Google Scholar] [CrossRef]
- Huitema, L.F.A.; Apschner, A.; Logister, I.; Spoorendonk, K.M.; Bussmann, J.; Hammond, C.L.; Schulte-Merker, S. Entpd5 Is Essential for Skeletal Mineralization and Regulates Phosphate Homeostasis in Zebrafish. Proc. Natl. Acad. Sci. USA 2012, 109, 21372–21377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, K.C.; Hill, D.; Otero, M.; Plumb, D.A.; Froemel, D.; Dragomir, C.L.; Maretzky, T.; Boskey, A.; Crawford, H.C.; Selleri, L.; et al. ADAM17 Controls Endochondral Ossification by Regulating Terminal Differentiation of Chondrocytes. Mol. Cell. Biol. 2013, 33, 3077–3090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murthy, A.; Shao, Y.W.; Narala, S.R.; Molyneux, S.D.; Zúñiga-Pflücker, J.C.; Khokha, R. Notch Activation by the Metalloproteinase ADAM17 Regulates Myeloproliferation and Atopic Barrier Immunity by Suppressing Epithelial Cytokine Synthesis. Immunity 2012, 36, 105–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araya, H.F.; Sepulveda, H.; Lizama, C.O.; Vega, O.A.; Jerez, S.; Briceño, P.F.; Thaler, R.; Riester, S.M.; Antonelli, M.; Salazar-Onfray, F.; et al. Expression of the Ectodomain-Releasing Protease ADAM17 Is Directly Regulated by the Osteosarcoma and Bone-Related Transcription Factor RUNX2. J. Cell. Biochem. 2018, 119, 8204–8219. [Google Scholar] [CrossRef]
- Coronel, R.; Bernabeu-Zornoza, A.; Palmer, C.; Muñiz-Moreno, M.; Zambrano, A.; Cano, E.; Liste, I. Role of Amyloid Precursor Protein (APP) and Its Derivatives in the Biology and Cell Fate Specification of Neural Stem Cells. Mol. Neurobiol. 2018, 55, 7107–7117. [Google Scholar] [CrossRef]
- Pan, J.-X.; Tang, F.; Xiong, F.; Xiong, L.; Zeng, P.; Wang, B.; Zhao, K.; Guo, H.; Shun, C.; Xia, W.-F.; et al. APP Promotes Osteoblast Survival and Bone Formation by Regulating Mitochondrial Function and Preventing Oxidative Stress. Cell Death Dis. 2018, 9, 1077. [Google Scholar] [CrossRef] [Green Version]
- Kwak, Y.-D.; Marutle, A.; Dantuma, E.; Merchant, S.; Bushnev, S.; Sugaya, K. Involvement of Notch Signaling Pathway in Amyloid Precursor Protein Induced Glial Differentiation. Eur. J. Pharm. 2011, 650, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Ramos, I.; Stamatakis, K.; Oeste, C.L.; Pérez-Sala, D. Vimentin as a Multifaceted Player and Potential Therapeutic Target in Viral Infections. Int. J. Mol. Sci. 2020, 21, 4675. [Google Scholar] [CrossRef]
- van Engeland, N.C.A.; Suarez Rodriguez, F.; Rivero-Müller, A.; Ristori, T.; Duran, C.L.; Stassen, O.M.J.A.; Antfolk, D.; Driessen, R.C.H.; Ruohonen, S.; Ruohonen, S.T.; et al. Vimentin Regulates Notch Signaling Strength and Arterial Remodeling in Response to Hemodynamic Stress. Sci. Rep. 2019, 9, 12415. [Google Scholar] [CrossRef]
- Lian, N.; Wang, W.; Li, L.; Elefteriou, F.; Yang, X. Vimentin Inhibits ATF4-Mediated Osteocalcin Transcription and Osteoblast Differentiation. J. Biol. Chem. 2009, 284, 30518–30525. [Google Scholar] [CrossRef] [Green Version]
- Pattabiraman, S.; Azad, G.K.; Amen, T.; Brielle, S.; Park, J.E.; Sze, S.K.; Meshorer, E.; Kaganovich, D. Vimentin Protects Differentiating Stem Cells from Stress. Sci. Rep. 2020, 10, 19525. [Google Scholar] [CrossRef]
- Shan, T.; Liu, J.; Wu, W.; Xu, Z.; Wang, Y. Roles of Notch Signaling in Adipocyte Progenitor Cells and Mature Adipocytes. J. Cell Physiol. 2017, 232, 1258–1261. [Google Scholar] [CrossRef]
- Ellisen, L.W.; Bird, J.; West, D.C.; Soreng, A.L.; Reynolds, T.C.; Smith, S.D.; Sklar, J. TAN-1, the Human Homolog of the Drosophila Notch Gene, Is Broken by Chromosomal Translocations in T Lymphoblastic Neoplasms. Cell 1991, 66, 649–661. [Google Scholar] [CrossRef]
- Heid, H.; Rickelt, S.; Zimbelmann, R.; Winter, S.; Schumacher, H.; Dörflinger, Y.; Kuhn, C.; Franke, W.W. On the Formation of Lipid Droplets in Human Adipocytes: The Organization of the Perilipin-Vimentin Cortex. PLoS ONE 2014, 9, e90386. [Google Scholar] [CrossRef] [Green Version]
- Uehara, S.; Udagawa, N.; Mukai, H.; Ishihara, A.; Maeda, K.; Yamashita, T.; Murakami, K.; Nishita, M.; Nakamura, T.; Kato, S.; et al. Protein Kinase N3 Promotes Bone Resorption by Osteoclasts in Response to Wnt5a-Ror2 Signaling. Sci. Signal. 2017, 10, eaan0023. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.K.; Deneen, B. Daam2 Is Required for Dorsal Patterning via Modulation of Canonical Wnt Signaling in the Developing Spinal Cord. Dev. Cell 2012, 22, 183–196. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, D.A.; Mullins, R.D. Cell Mechanics and the Cytoskeleton. Nature 2010, 463, 485–492. [Google Scholar] [CrossRef] [Green Version]
- Sankaran, J.; Uzer, G.; van Wijnen, A.J.; Rubin, J. Gene Regulation through Dynamic Actin Control of Nuclear Structure. Exp. Biol. Med. 2019, 244, 1345–1353. [Google Scholar] [CrossRef] [Green Version]
- Poukkula, M.; Kremneva, E.; Serlachius, M.; Lappalainen, P. Actin-Depolymerizing Factor Homology Domain: A Conserved Fold Performing Diverse Roles in Cytoskeletal Dynamics. Cytoskeleton 2011, 68, 471–490. [Google Scholar] [CrossRef]
- Xue, B.; Robinson, R.C. Guardians of the Actin Monomer. Eur. J. Cell Biol. 2013, 92, 316–332. [Google Scholar] [CrossRef]
- Mooren, O.L.; Galletta, B.J.; Cooper, J.A. Roles for Actin Assembly in Endocytosis. Annu. Rev. Biochem. 2012, 81, 661–686. [Google Scholar] [CrossRef] [PubMed]
- Wilbur, J.D.; Chen, C.-Y.; Manalo, V.; Hwang, P.K.; Fletterick, R.J.; Brodsky, F.M. Actin Binding by Hip1 (Huntingtin-Interacting Protein 1) and Hip1R (Hip1-Related Protein) Is Regulated by Clathrin Light Chain. J. Biol. Chem. 2008, 283, 32870–32879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saadi, I.; Alkuraya, F.S.; Gisselbrecht, S.S.; Goessling, W.; Cavallesco, R.; Turbe-Doan, A.; Petrin, A.L.; Harris, J.; Siddiqui, U.; Grix, A.W.; et al. Deficiency of the Cytoskeletal Protein SPECC1L Leads to Oblique Facial Clefting. Am. J. Hum. Genet. 2011, 89, 44–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, W.; Guo, X.; Thein, S.; Xu, F.; Sugii, S.; Baas, P.W.; Radda, G.K.; Han, W. Regulation of Adipogenesis by Cytoskeleton Remodelling Is Facilitated by Acetyltransferase MEC-17-Dependent Acetylation of α-Tubulin. Biochem. J. 2013, 449, 605–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bontems, F.; Fish, R.J.; Borlat, I.; Lembo, F.; Chocu, S.; Chalmel, F.; Borg, J.-P.; Pineau, C.; Neerman-Arbez, M.; Bairoch, A.; et al. C2orf62 and TTC17 Are Involved in Actin Organization and Ciliogenesis in Zebrafish and Human. PLoS ONE 2014, 9, e86476. [Google Scholar] [CrossRef] [Green Version]
- Xiong, L.-L.; Qiu, D.-L.; Xiu, G.-H.; Al-Hawwas, M.; Jiang, Y.; Wang, Y.-C.; Hu, Y.; Chen, L.; Xia, Q.-J.; Wang, T.-H. DPYSL2 Is a Novel Regulator for Neural Stem Cell Differentiation in Rats: Revealed by Panax Notoginseng Saponin Administration. Stem Cell Res. Ther. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Rmaileh, A.A.; Solaimuthu, B.; Yosef, M.B.; Khatib, A.; Lichtenstein, M.; Tanna, M.; Hayashi, A.; Pillar, N.; Shaul, Y.D. Dihydropyrimidinase-like 2 (DPYSL2) Regulates Breast Cancer Migration via a JAK/STAT3/Vimentin Axis. bioRxiv 2021. [Google Scholar] [CrossRef]
- Mettlen, M.; Chen, P.-H.; Srinivasan, S.; Danuser, G.; Schmid, S.L. Regulation of Clathrin-Mediated Endocytosis. Annu. Rev. Biochem. 2018, 87, 871–896. [Google Scholar] [CrossRef]
- Rao, D.S.; Chang, J.C.; Kumar, P.D.; Mizukami, I.; Smithson, G.M.; Bradley, S.V.; Parlow, A.F.; Ross, T.S. Huntingtin Interacting Protein 1 Is a Clathrin Coat Binding Protein Required for Differentiation of Late Spermatogenic Progenitors. Mol. Cell. Biol. 2001, 21, 7796–7806. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Liu, F. Regulation of Alternative Splicing of Tau Exon 10. Neurosci. Bull. 2014, 30, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Neves-Carvalho, A.; Duarte-Silva, S.; Silva, J.; Almeida, B.; Heetveld, S.; Sotiropoulos, I.; Heutink, P.; Li, K.W.; Maciel, P. Regulation of Neuronal MRNA Splicing and Tau Isoform Ratio by ATXN3 through Deubiquitylation of Splicing Factors. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Pîrşcoveanu, D.F.V.; Pirici, I.; Tudorică, V.; Bălşeanu, T.A.; Albu, V.C.; Bondari, S.; Bumbea, A.M.; Pîrşcoveanu, M. Tau Protein in Neurodegenerative Diseases—A Review. Rom. J. Morphol. Embryol. 2017, 58, 1141–1150. [Google Scholar]
- Liu, F.; Gong, C.-X. Tau Exon 10 Alternative Splicing and Tauopathies. Mol. Neurodegener. 2008, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Sarkissian, M.; Winne, A.; Lafyatis, R. The Mammalian Homolog of Suppressor-of-White-Apricot Regulates Alternative MRNA Splicing of CD45 Exon 4 and Fibronectin IIICS. J. Biol. Chem. 1996, 271, 31106–31114. [Google Scholar] [CrossRef] [Green Version]
- Giraud, G.; Terrone, S.; Bourgeois, C.F. Functions of DEAD Box RNA Helicases DDX5 and DDX17 in Chromatin Organization and Transcriptional Regulation. BMB Rep. 2018, 51, 613–622. [Google Scholar] [CrossRef] [Green Version]
- Dardenne, E.; Polay Espinoza, M.; Fattet, L.; Germann, S.; Lambert, M.-P.; Neil, H.; Zonta, E.; Mortada, H.; Gratadou, L.; Deygas, M.; et al. RNA Helicases DDX5 and DDX17 Dynamically Orchestrate Transcription, MiRNA, and Splicing Programs in Cell Differentiation. Cell. Rep. 2014, 7, 1900–1913. [Google Scholar] [CrossRef] [Green Version]
- Laaref, A.M.; Manchon, L.; Bareche, Y.; Lapasset, L.; Tazi, J. The Core Spliceosomal Factor U2AF1 Controls Cell-Fate Determination via the Modulation of Transcriptional Networks. RNA Biol. 2020, 17, 857–871. [Google Scholar] [CrossRef]
- Doherty, G.J.; McMahon, H.T. Mechanisms of Endocytosis. Available online: http://0-www-annualreviews-org.brum.beds.ac.uk/doi/abs/10.1146/annurev.biochem.78.081307.110540 (accessed on 5 July 2021).
- Kaddai, V.; Gonzalez, T.; Keslair, F.; Grémeaux, T.; Bonnafous, S.; Gugenheim, J.; Tran, A.; Gual, P.; Le Marchand-Brustel, Y.; Cormont, M. Rab4b Is a Small GTPase Involved in the Control of the Glucose Transporter GLUT4 Localization in Adipocyte. PLoS ONE 2009, 4, 5257. [Google Scholar] [CrossRef] [Green Version]
- Sadler, J.B.A.; Bryant, N.J.; Gould, G.W. Characterization of VAMP Isoforms in 3T3-L1 Adipocytes: Implications for GLUT4 Trafficking. Mol. Biol. Cell 2015, 26, 530–536. [Google Scholar] [CrossRef]
- Nakazawa, S.; Gotoh, N.; Matsumoto, H.; Murayama, C.; Suzuki, T.; Yamamoto, T. Expression of Sorting Nexin 18 (SNX18) Is Dynamically Regulated in Developing Spinal Motor Neurons. J. Histochem. Cytochem. 2011, 59, 202–213. [Google Scholar] [CrossRef] [Green Version]
- Park, J.; Kim, Y.; Lee, S.; Park, J.J.; Park, Z.Y.; Sun, W.; Kim, H.; Chang, S. SNX18 Shares a Redundant Role with SNX9 and Modulates Endocytic Trafficking at the Plasma Membrane. J. Cell Sci. 2010, 123, 1742–1750. [Google Scholar] [CrossRef] [Green Version]
- Ma, M.P.C.; Chircop, M. SNX9, SNX18 and SNX33 Are Required for Progression through and Completion of Mitosis. J. Cell Sci. 2012, 125, 4372–4382. [Google Scholar] [CrossRef] [Green Version]
- Kliza, K.; Husnjak, K. Resolving the Complexity of Ubiquitin Networks. Front. Mol. Biosci. 2020, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Gavin, J.M.; Hoar, K.; Xu, Q.; Ma, J.; Lin, Y.; Chen, J.; Chen, W.; Bruzzese, F.J.; Harrison, S.; Mallender, W.D.; et al. Mechanistic Study of Uba5 Enzyme and the Ufm1 Conjugation Pathway. J. Biol. Chem. 2014, 289, 22648–22658. [Google Scholar] [CrossRef] [Green Version]
- Tatsumi, K.; Yamamoto-Mukai, H.; Shimizu, R.; Waguri, S.; Sou, Y.-S.; Sakamoto, A.; Taya, C.; Shitara, H.; Hara, T.; Chung, C.H.; et al. The Ufm1-Activating Enzyme Uba5 Is Indispensable for Erythroid Differentiation in Mice. Nat. Commun. 2011, 2, 181. [Google Scholar] [CrossRef]
- Olzmann, J.A.; Kopito, R.R.; Christianson, J.C. The Mammalian Endoplasmic Reticulum-Associated Degradation System. Cold Spring Harb. Perspect. Biol. 2013, 5, a013185. [Google Scholar] [CrossRef] [Green Version]
- Sha, H.; Sun, S.; Francisco, A.; Ehrhardt, N.; Xue, Z.; Liu, L.; Lawrence, P.; Mattijssen, F.; Guber, R.; Panhwar, M.S.; et al. The ER-Associated Degradation Adaptor Protein Sel1L Regulates LPL Secretion and Lipid Metabolism. Cell Metab. 2014, 20, 458–470. [Google Scholar] [CrossRef] [Green Version]
- Williams, K.J. Molecular Processes That Handle—And Mishandle—Dietary Lipids. J. Clin. Investig. 2008, 118, 3247–3259. [Google Scholar] [CrossRef] [Green Version]
- Philp, L.K.; Butler, M.S.; Hickey, T.E.; Butler, L.M.; Tilley, W.D.; Day, T.K. SGTA: A New Player in the Molecular Co-Chaperone Game. Horm. Cancer 2013, 4, 343–357. [Google Scholar] [CrossRef]
- Francisco, A.B.; Singh, R.; Li, S.; Vani, A.K.; Yang, L.; Munroe, R.J.; Diaferia, G.; Cardano, M.; Biunno, I.; Qi, L.; et al. Deficiency of Suppressor Enhancer Lin12 1 Like (SEL1L) in Mice Leads to Systemic Endoplasmic Reticulum Stress and Embryonic Lethality. J. Biol. Chem. 2010, 285, 13694–13703. [Google Scholar] [CrossRef] [Green Version]
- Laplante, D.P.; Zelazo, P.R.; Brunei, A.; King, S. Functional Play at 2 Years of Age: Effects of Prenatal Maternal Stress. Infancy 2007, 12, 69–93. [Google Scholar] [CrossRef] [PubMed]
- Cao-Lei, L.; Elgbeili, G.; Massart, R.; Laplante, D.P.; Szyf, M.; King, S. Pregnant Women’s Cognitive Appraisal of a Natural Disaster Affects DNA Methylation in Their Children 13 Years Later: Project Ice Storm. Transl. Psychiatry 2015, 5, e515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidal, M.A.; Kilroy, G.E.; Lopez, M.J.; Johnson, J.R.; Moore, R.M.; Gimble, J.M. Characterization of Equine Adipose Tissue-Derived Stromal Cells: Adipogenic and Osteogenic Capacity and Comparison with Bone Marrow-Derived Mesenchymal Stromal Cells. Vet. Surg. 2007, 36, 613–622. [Google Scholar] [CrossRef] [PubMed]
- Bhargava, U.; Bar-Lev, M.; Bellows, C.G.; Aubin, J.E. Ultrastructural Analysis of Bone Nodules Formed in vitro by Isolated Fetal Rat Calvaria Cells. Bone 1988, 9, 155–163. [Google Scholar] [CrossRef]
- Wessel, D.; Flügge, U.I. A Method for the Quantitative Recovery of Protein in Dilute Solution in the Presence of Detergents and Lipids. Anal. Biochem. 1984, 138, 141–143. [Google Scholar] [CrossRef]
- Wu, M.; McCain, J.S.P.; Rowland, E.; Middag, R.; Sandgren, M.; Allen, A.E.; Bertrand, E.M. Manganese and Iron Deficiency in Southern Ocean Phaeocystis Antarctica Populations Revealed through Taxon-Specific Protein Indicators. Nat. Commun. 2019, 10, 3582. [Google Scholar] [CrossRef] [Green Version]
- Käll, L.; Canterbury, J.D.; Weston, J.; Noble, W.S.; MacCoss, M.J. Semi-Supervised Learning for Peptide Identification from Shotgun Proteomics Datasets. Nat. Methods 2007, 4, 923–925. [Google Scholar] [CrossRef]
- Plubell, D.L.; Wilmarth, P.A.; Zhao, Y.; Fenton, A.M.; Minnier, J.; Reddy, A.P.; Klimek, J.; Yang, X.; David, L.L.; Pamir, N. Extended Multiplexing of Tandem Mass Tags (TMT) Labeling Reveals Age and High Fat Diet Specific Proteome Changes in Mouse Epididymal Adipose Tissue. Mol. Cell. Proteom. 2017, 16, 873–890. [Google Scholar] [CrossRef] [Green Version]
- Duncan, D.B. Multiple Range and Multiple F Tests. Biometrics 1955, 11, 1–42. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2021, 50, D543–D552. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample No. | Condition | Reagent |
|---|---|---|
| S1 | SM1 37 °C | TMT10-126 |
| S2 | SM2 37 °C | TMT10-127N |
| S3 | SM3 37 °C | TMT10-127C |
| S4 | OM1 37 °C | TMT10-128N |
| S5 | OM2 37 °C | TMT10-128C |
| S6 | OM3 37 °C | TMT10-129N |
| S7 | AD1 37 °C | TMT10-129C |
| S8 | AD2 37 °C | TMT10-130N |
| S9 | AD3 37 °C | TMT10-130C |
| Pool 1 | All | TMT10-131 |
| Pool 2 | All | TMT11-131C |
| Sample No. | Condition | Reagent |
|---|---|---|
| S10 | SM1 42 °C | TMT10-126 |
| S11 | SM2 42 °C | TMT10-127N |
| S12 | SM3 42 °C | TMT10-127C |
| S13 | OM1 42 °C | TMT10-128N |
| S14 | OM2 42 °C | TMT10-128C |
| S15 | OM3 42 °C | TMT10-129N |
| S16 | AD1 42 °C | TMT10-129C |
| S17 | AD2 42 °C | TMT10-130N |
| S18 | AD3 42 °C | TMT10-130C |
| Pool 1 | All | TMT10-131 |
| Pool 2 | All | TMT11-131C |
| Step | CV (Column Volumes) |
|---|---|
| Equilibrate | 0.83 |
| Sample application | 0.5 |
| Column wash | 0.83 |
| Elution | 16 |
| Column wash | 0.83 |
| Equilibrate | 1.66 |
| SM | OM | AD | |
|---|---|---|---|
n = 3 | n = 3 | n = 3 | 37 °C |
n = 3 | n = 3 | n = 3 | 42 °C |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd-El-Aziz, A.; Riveroll, A.; Esparza-Gonsalez, B.; McDuffee, L.; Cohen, A.M.; Fenech, A.L.; Montelpare, W.J. Heat Shock Alters the Proteomic Profile of Equine Mesenchymal Stem Cells. Int. J. Mol. Sci. 2022, 23, 7233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137233
Abd-El-Aziz A, Riveroll A, Esparza-Gonsalez B, McDuffee L, Cohen AM, Fenech AL, Montelpare WJ. Heat Shock Alters the Proteomic Profile of Equine Mesenchymal Stem Cells. International Journal of Molecular Sciences. 2022; 23(13):7233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137233
Chicago/Turabian StyleAbd-El-Aziz, Ahmad, Angela Riveroll, Blanca Esparza-Gonsalez, Laurie McDuffee, Alejandro M. Cohen, Adam L. Fenech, and William J. Montelpare. 2022. "Heat Shock Alters the Proteomic Profile of Equine Mesenchymal Stem Cells" International Journal of Molecular Sciences 23, no. 13: 7233. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137233