
Bortezomib Eliminates Persistent Chlamydia trachomatis Infection through Rapid and Specific Host Cell Apoptosis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
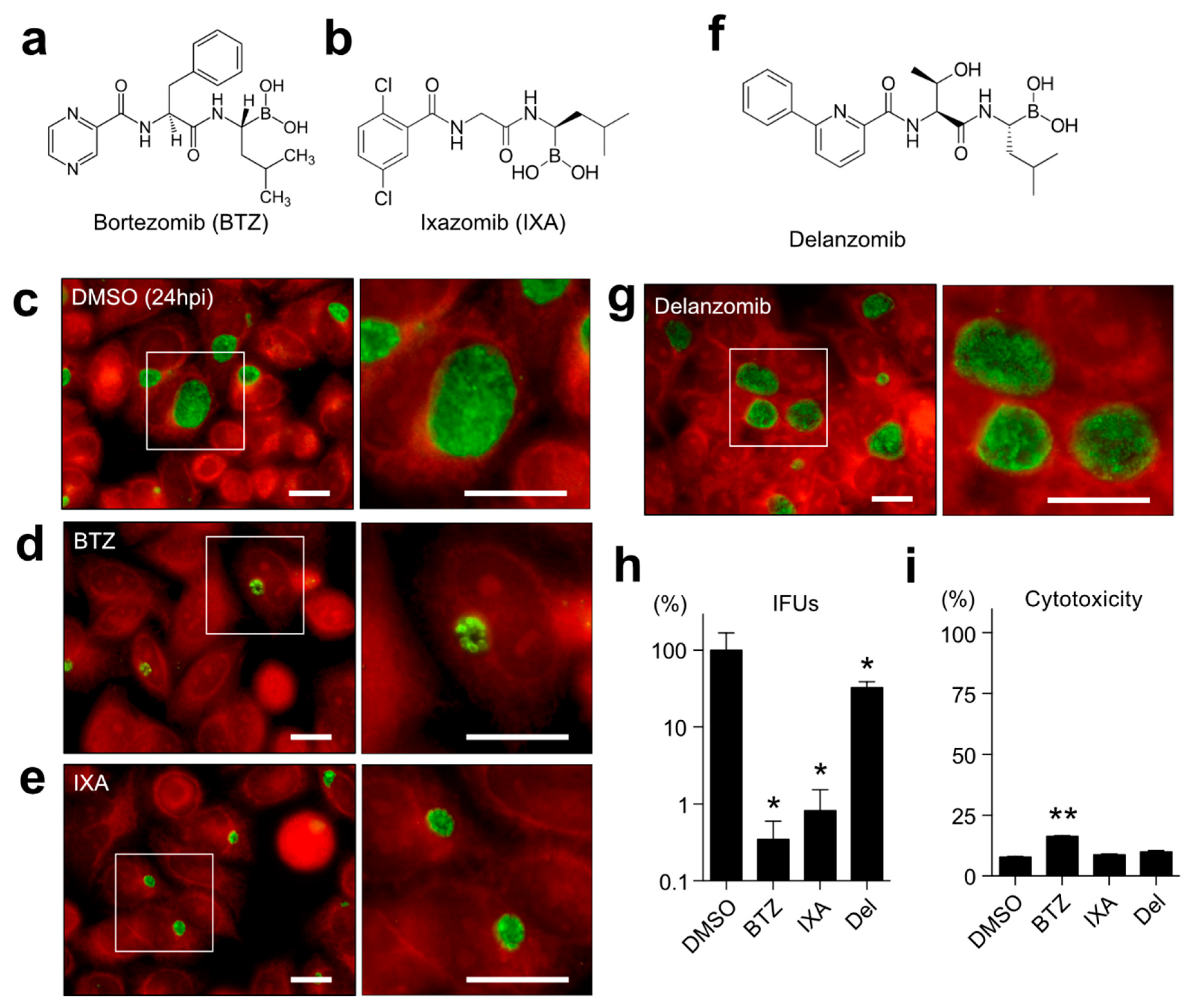
2.1. Both BTZ and IXA but Not Delanzomib Strictly Suppress the Chlamydial Growth
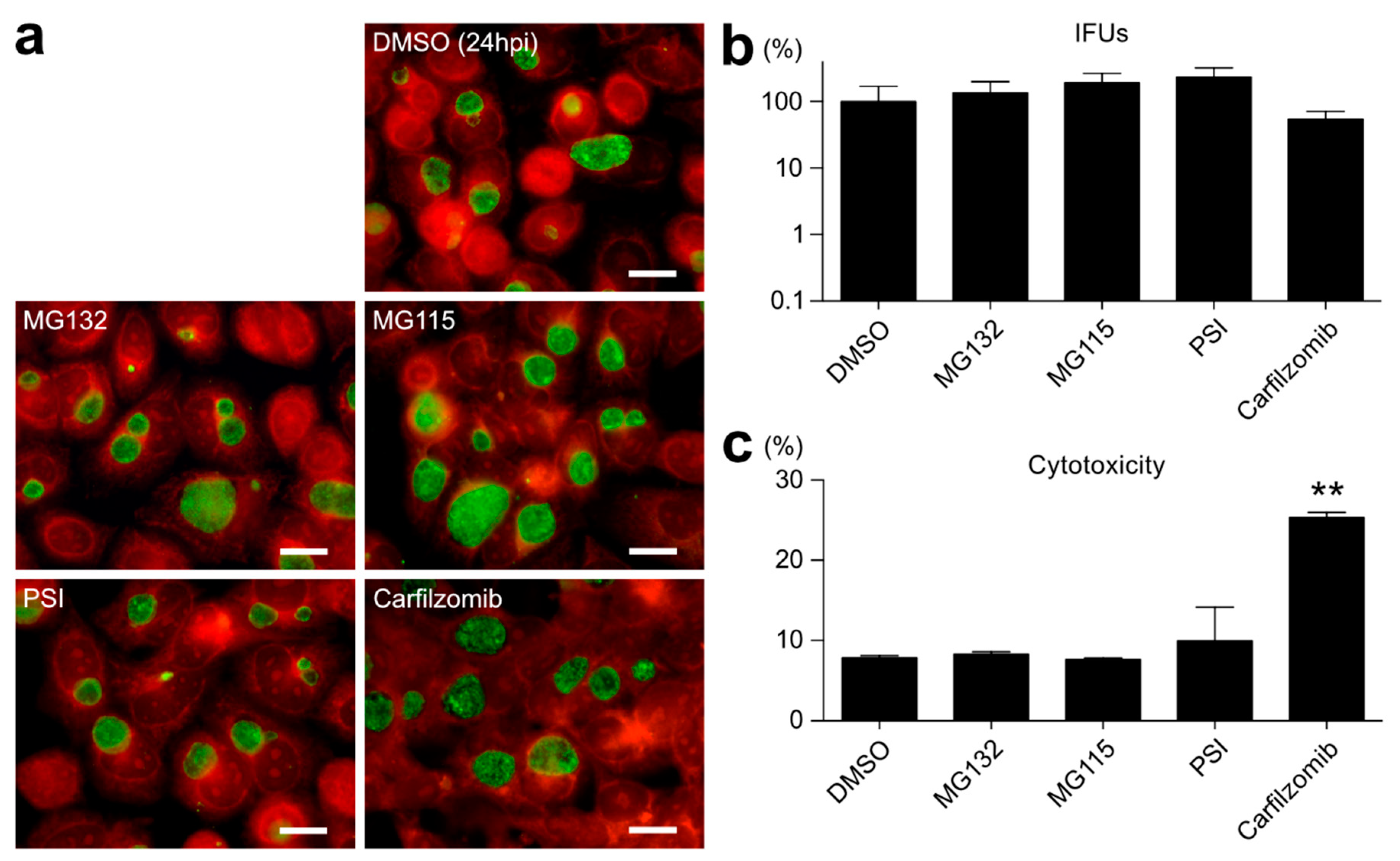
2.2. Proteasome Inhibitors Other than BTZ or IXA Do Not Inhibit Chlamydial Growth
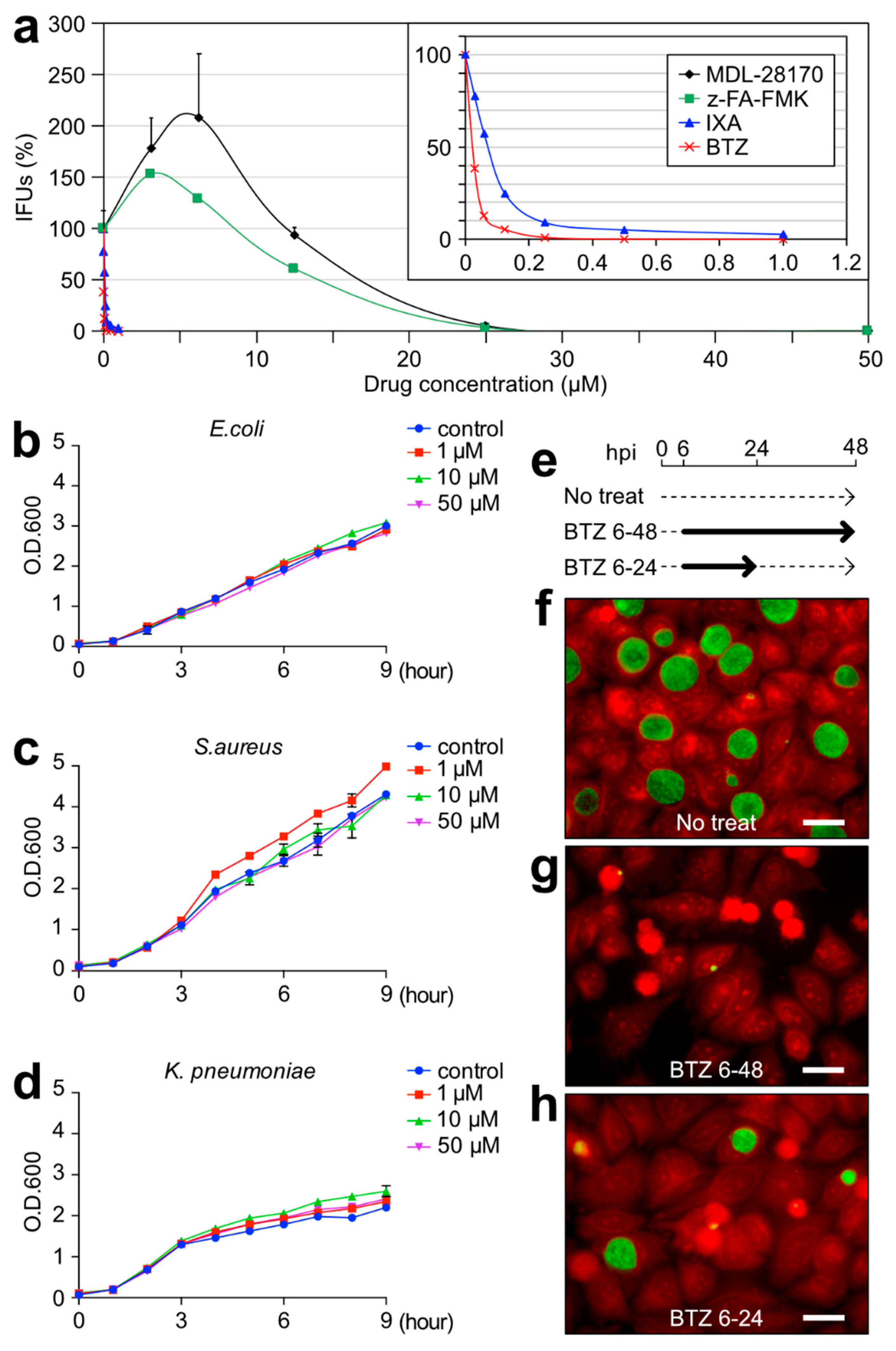
2.3. BTZ Has Antichlamydial Activity Comparable to Existing Antimicrobial Drugs
2.4. BTZ Arrests Chlamydial Replication but Does Not Induce the Persistent Form
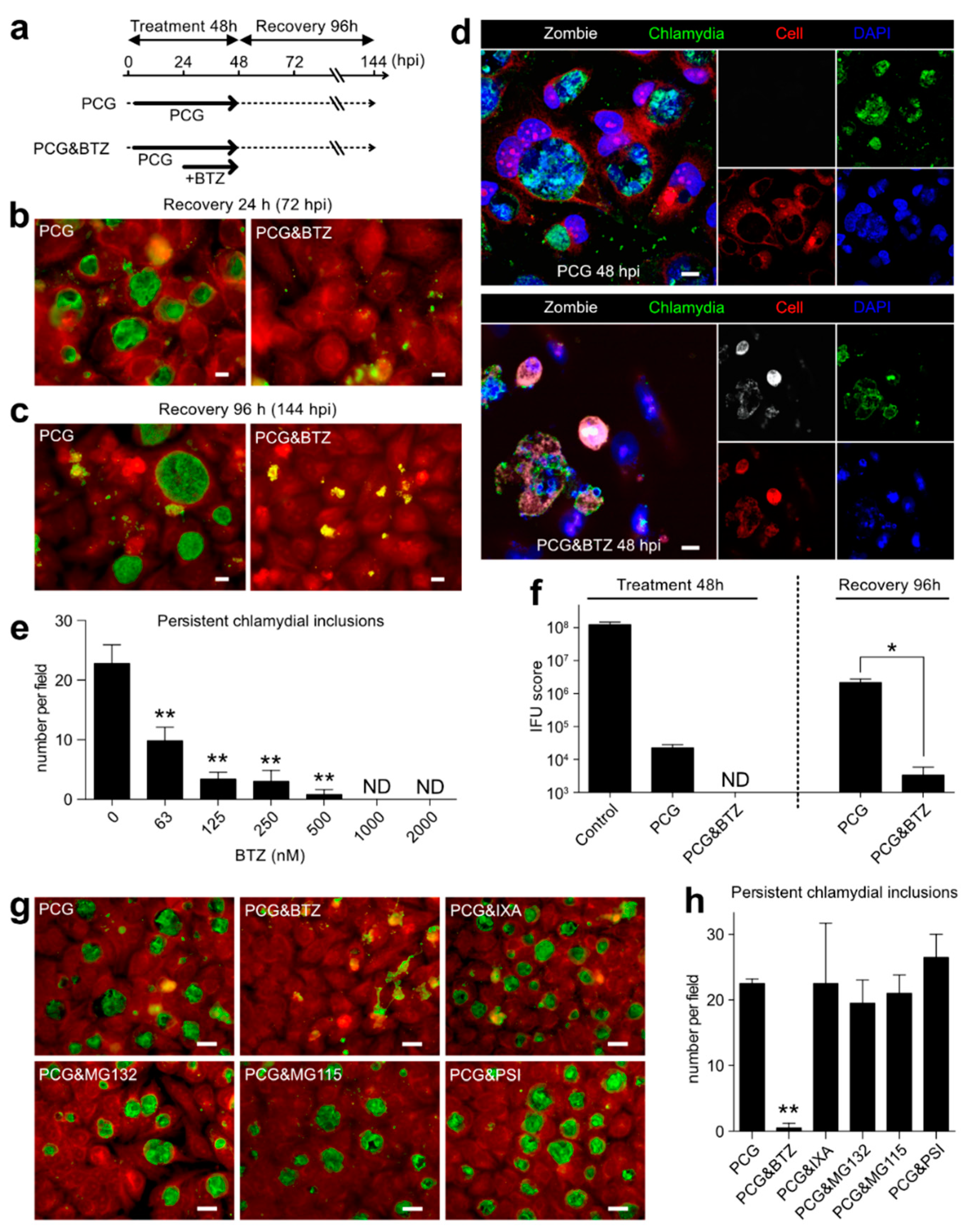
2.5. BTZ Specifically Killed Persistently Chlamydia-Infected Host Cells
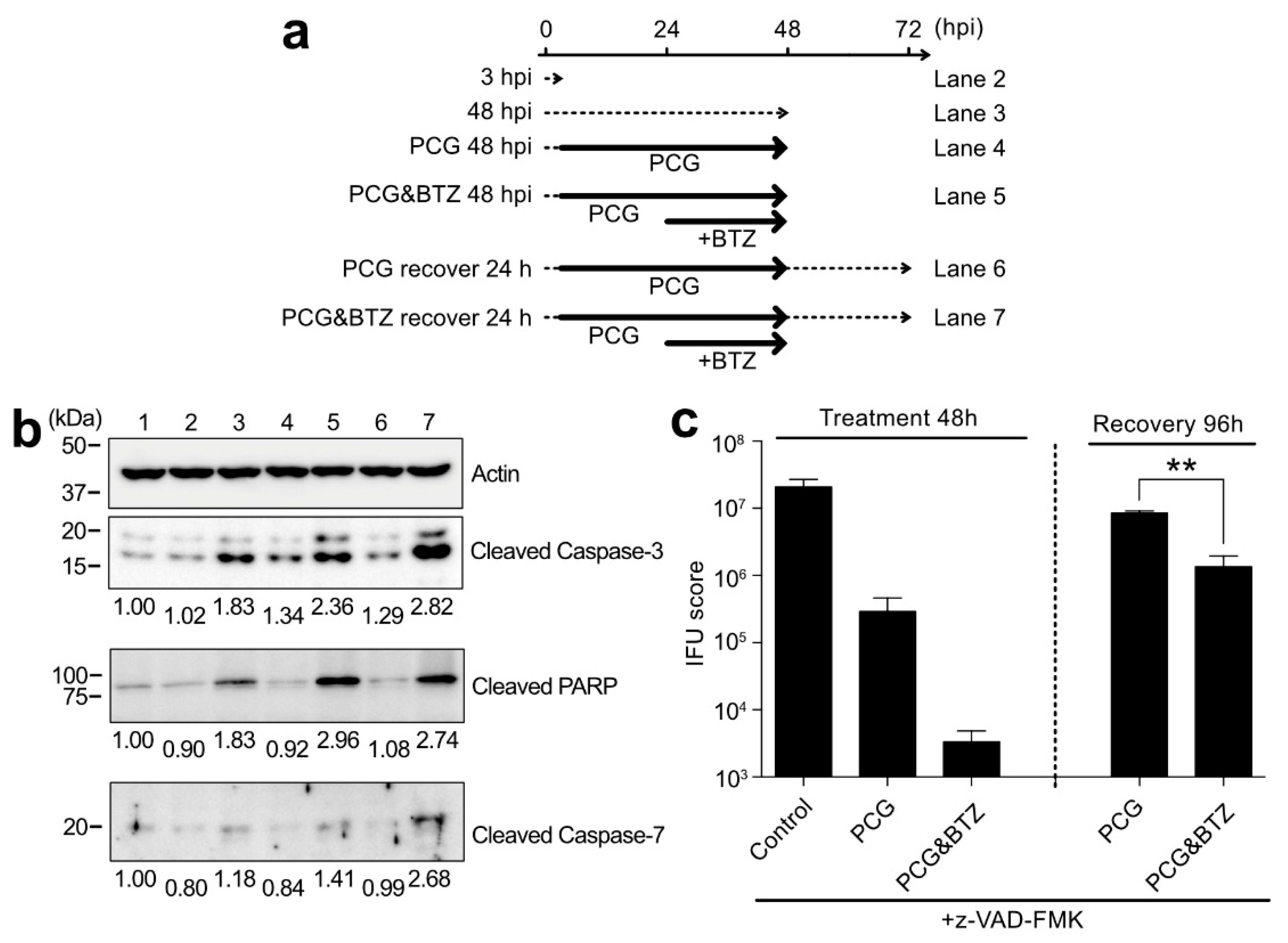
2.6. BTZ Induces Persistently Chlamydia-Infected Cell Apoptosis through Caspase-3 Activation and Following Pathway
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Bacterial Culture, Infection, and Chemical Treatment
4.3. Fluorescent Immunostaining and Microscopy
4.4. Inclusion Forming Unit Assay
4.5. Cytotoxicity Assay
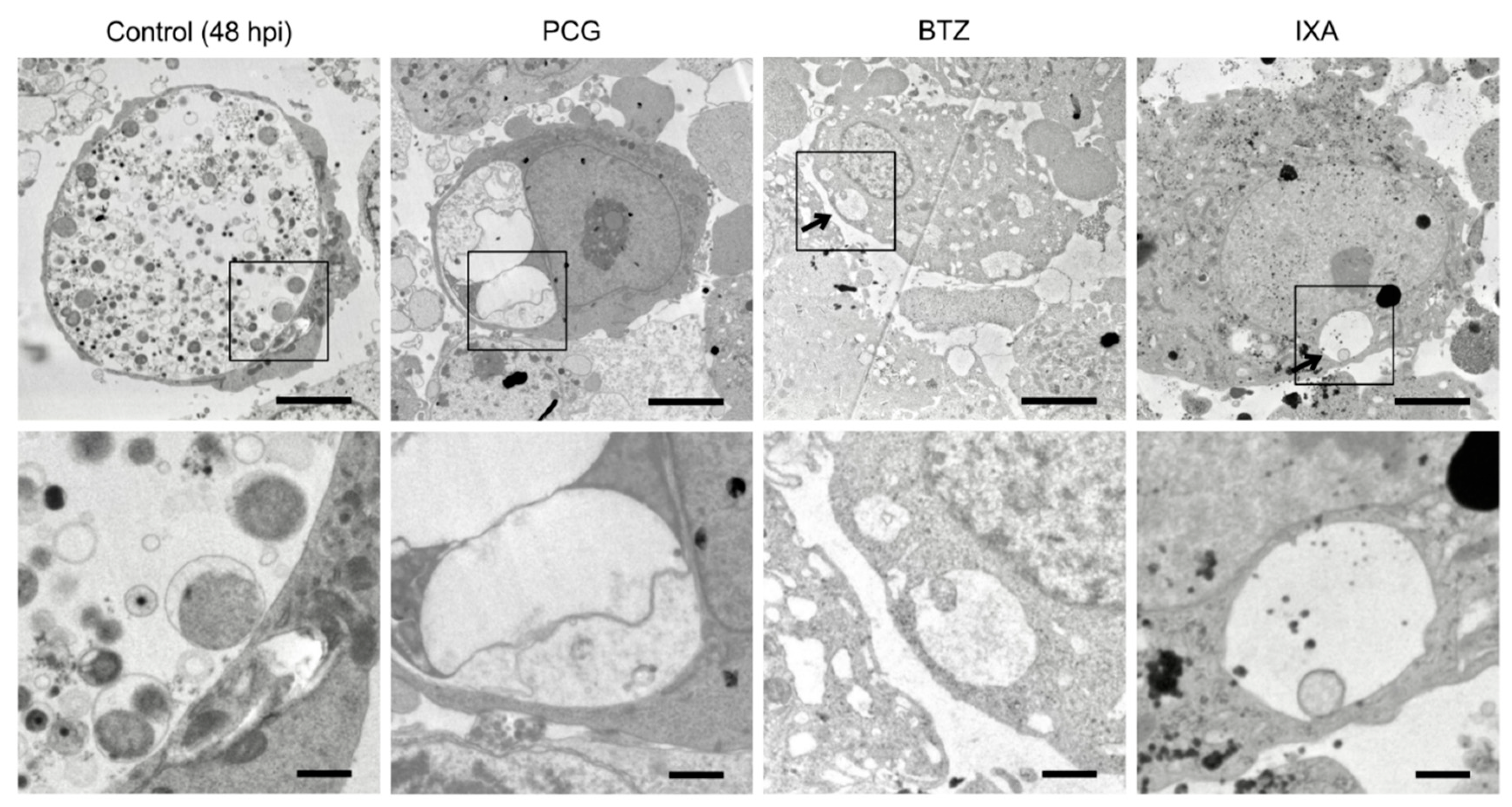
4.6. Electron Microscopy
4.7. Western Blotting
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bastidas, R.J.; Elwell, C.A.; Engel, J.N.; Valdivia, R.H. Chlamydial Intracellular Survival Strategies. Cold Spring Harb. Perspect. Med. 2013, 3, a010256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elwell, C.; Mirrashidi, K.; Engel, J. Chlamydia cell biology and pathogenesis. Nat. Rev. Microbiol. 2016, 14, 385–400. [Google Scholar] [CrossRef] [PubMed]
- Miller, K.E. Diagnosis and treatment of Chlamydia trachomatis infection. Am. Fam. Physician 2006, 73, 1411–1416. [Google Scholar] [PubMed]
- Prohl, A.; Lohr, M.; Ostermann, C.; Liebler-Tenorio, E.; Berndt, A.; Schroedl, W.; Rothe, M.; Schubert, E.; Sachse, K.; Reinhold, P. Enrofloxacin and Macrolides Alone or in Combination with Rifampicin as Antimicrobial Treatment in a Bovine Model of Acute Chlamydia psittaci Infection. PLoS ONE 2015, 10, e0119736. [Google Scholar] [CrossRef] [Green Version]
- Hocking, J.S.; Vodstrcil, L.A.; Huston, W.M.; Timms, P.; Chen, M.Y.; Worthington, K.; McIver, R.; Tabrizi, S.N.; Australian Chlamydia Treatment, S. A cohort study of Chlamydia trachomatis treatment failure in women: A study protocol. BMC Infect. Dis. 2013, 13, 379. [Google Scholar] [CrossRef] [Green Version]
- Hocking, J.S.; Kong, F.Y.S.; Timms, P.; Huston, W.M.; Tabrizi, S.N. Treatment of rectal chlamydia infection may be more complicated than we originally thought. J. Antimicrob. Chemother. 2015, 70, 961–964. [Google Scholar] [CrossRef] [Green Version]
- Witkin, S.S.; Minis, E.; Athanasiou, A.; Leizer, J.; Linhares, I.M. Chlamydia trachomatis: The Persistent Pathogen. Clin. Vaccine Immunol. 2017, 24, e00203-17. [Google Scholar] [CrossRef] [Green Version]
- Panzetta, M.E.; Valdivia, R.H.; Saka, H.A. Chlamydia Persistence: A Survival Strategy to Evade Antimicrobial Effects in-vitro and in-vivo. Front. Microbiol. 2018, 9, 3101. [Google Scholar] [CrossRef]
- Wyrick, P.B. Chlamydia trachomatis Persistence In Vitro: An Overview. J. Infect. Dis. 2010, 201, S88–S95. [Google Scholar] [CrossRef] [Green Version]
- Schuchardt, L.; Rupp, J. Chlamydia trachomatis as the Cause of Infectious Infertility: Acute, Repetitive or Persistent Long-Term Infection? Biol. Chlamydia 2018, 412, 159–182. [Google Scholar] [CrossRef]
- Somani, J.; Bhullar, V.B.; Workowski, K.A.; Farshy, C.E.; Black, C.M. Multiple drug-resistant Chlamydia trachomatis associated with clinical treatment failure. J. Infect. Dis. 2000, 181, 1421–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itoh, R.; Soejima, T.; Hiromatsu, K. Anti-chlamydial activities of cell-permeable hydrophobic dipeptide-containing derivatives. J. Infect. Chemother. 2019, 25, 987–994. [Google Scholar] [CrossRef] [PubMed]
- Mangiacavalli, S.; Pochintesta, L.; Pascutto, C.; Cocito, F.; Pompa, A.; Cazzola, M.; Corso, A. Good clinical activity and favorable toxicity profile of once weekly bortezomib, fotemustine, and dexamethasone (B-MuD) for the treatment of relapsed multiple myeloma. Am. J. Hematol. 2013, 88, 102–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, M.; Zhang, J.J.; Tang, R.; Wang, Y.H.; Zhan, X.K.; Fan, S.B.; Huang, Z.X.; Zhong, Y.P.; Li, X. Ixazomib-based maintenance therapy after bortezomib-based induction in patients with multiple myeloma not undergoing transplantation: A real-world study. Cancer Med. 2022, 11, 2173–2183. [Google Scholar] [CrossRef]
- Oka, S.; Ono, K. Successful ixazomib treatment for relapsed and refractory acute myeloid leukemia transformed from myelodysplastic syndrome. Clin. Case Rep. 2021, 9, e04287. [Google Scholar] [CrossRef]
- Li, J.; Zhuo, J.Y.; Zhou, W.; Hong, J.W.; Chen, R.G.; Xie, H.Y.; Zhou, L.; Zheng, S.S.; Jiang, D.H. Endoplasmic reticulum stress triggers delanzomib-induced apoptosis in HCC cells through the PERK/eIF2 alpha/ATF4/CHOP pathway. Am. J. Transl. Res. 2020, 12, 2875. [Google Scholar]
- Gallerani, E.; Zucchetti, M.; Brunelli, D.; Marangon, E.; Noberasco, C.; Hess, D.; Delmonte, A.; Martinelli, G.; Bohm, S.; Driessen, C.; et al. A first in human phase I study of the proteasome inhibitor CEP-18770 in patients with advanced solid tumours and multiple myeloma. Eur. J. Cancer 2013, 49, 290–296. [Google Scholar] [CrossRef]
- Richardson, P.G.; Sonneveld, P.; Schuster, M.W.; Irwin, D.; Stadtmauer, E.A.; Facon, T.; Harousseau, J.L.; Ben-Yehuda, D.; Lonial, S.; Goldschmidt, H.; et al. Bortezomib or high-dose dexamethasone for relapsed multiple myeloma. N. Engl. J. Med. 2005, 352, 2487–2498. [Google Scholar] [CrossRef] [Green Version]
- Groll, M.; Berkers, C.R.; Ploegh, H.L.; Ovaa, H. Crystal structure of the boronic acid-based proteasome inhibitor bortezomib in complex with the yeast 20S proteasome. Structure 2006, 14, 451–456. [Google Scholar] [CrossRef] [Green Version]
- Li, J.L.; Yang, N.J.; Huang, L.; Chen, D.D.; Zhao, Y.; Tang, M.M.; Fan, H.Z.; Bao, X.F. Pyocyanin Inhibits Chlamydia Infection by Disabling Infectivity of the Elementary Body and Disrupting Intracellular Growth. Antimicrob. Agents Chemother. 2018, 62, e02260-17. [Google Scholar] [CrossRef] [Green Version]
- Moreira, W.; Ngan, G.J.Y.; Low, J.L.; Poulsen, A.; Chia, B.C.S.; Ang, M.J.Y.; Yap, A.; Fulwood, J.; Lakshmanan, U.; Lim, J.; et al. Target Mechanism-Based Whole-Cell Screening Identifies Bortezomib as an Inhibitor of Caseinolytic Protease in Mycobacteria. MBio 2015, 6, e00253-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandu, D.; Nandi, D. Comparative genomics and functional roles of the ATP-dependent proteases Lon and Clp during cytosolic protein degradation. Res. Microbiol. 2004, 155, 710–719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldberg, A.L.; Moerschell, R.P.; Chung, C.H.; Maurizi, M.R. Atp-Dependent Protease La (Lon) from Escherichia-Coli. Proteolytic Enzym. Serine Cysteine Pept. 1994, 244, 350–375. [Google Scholar]
- Maurizi, M.R.; Thompson, M.W.; Singh, S.K.; Kim, S.H. Endopeptidase-Clp—Atp-Dependent Clp Protease from Escherichia-Coli. Proteolytic Enzym. Serine Cysteine Pept. 1994, 244, 314–331. [Google Scholar]
- Wood, N.A.; Blocker, A.M.; Seleem, M.A.; Conda-Sheridan, M.; Fisher, D.J.; Ouellette, S.P. The ClpX and ClpP2 Orthologs of Chlamydia trachomatis Perform Discrete and Essential Functions in Organism Growth and Development. MBio 2020, 11, e02016-20. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.A.; Chung, K.Y.; Blocker, A.M.; de Almeida, N.R.; Conda-Sheridan, M.; Fisher, D.J.; Ouellette, S.P. Initial Characterization of the Two ClpP Paralogs of Chlamydia trachomatis Suggests Unique Functionality for Each. J. Bacteriol. 2019, 201, e00635-18. [Google Scholar] [CrossRef] [Green Version]
- Muz, B.; Ghazarian, R.N.; Ou, M.; Luderer, M.J.; Kusdono, H.D.; Azab, A.K. Spotlight on ixazomib: Potential in the treatment of multiple myeloma. Drug Des. Dev. Ther. 2016, 10, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Kupperman, E.; Lee, E.C.; Cao, Y.Y.; Bannerman, B.; Fitzgerald, M.; Berger, A.; Yu, J.; Yang, Y.; Hales, P.; Bruzzese, F.; et al. Evaluation of the Proteasome Inhibitor MLN9708 in Preclinical Models of Human Cancer. Cancer Res. 2010, 70, 1970–1980. [Google Scholar] [CrossRef] [Green Version]
- Dean, D.; Powers, V.C. Persistent Chlamydia trachomatis infections resist apoptotic stimuli. Infect. Immun. 2001, 69, 2442–2447. [Google Scholar] [CrossRef] [Green Version]
- Pushpakom, S.; Iorio, F.; Eyers, P.A.; Escott, K.J.; Hopper, S.; Wells, A.; Doig, A.; Guilliams, T.; Latimer, J.; McNamee, C.; et al. Drug repurposing: Progress, challenges and recommendations. Nat. Rev. Drug Discov. 2019, 18, 41–58. [Google Scholar] [CrossRef]
- Itoh, R.; Murakami, I.; Chou, B.; Ishii, K.; Soejima, T.; Suzuki, T.; Hiromatsu, K. Chlamydia pneumoniae harness host NLRP3 inflammasome-mediated caspase-1 activation for optimal intracellular growth in murine macrophages. Biochem. Biophys. Res. Commun. 2014, 452, 689–694. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Itoh, R.; Kurihara, Y.; Yoshimura, M.; Hiromatsu, K. Bortezomib Eliminates Persistent Chlamydia trachomatis Infection through Rapid and Specific Host Cell Apoptosis. Int. J. Mol. Sci. 2022, 23, 7434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137434
Itoh R, Kurihara Y, Yoshimura M, Hiromatsu K. Bortezomib Eliminates Persistent Chlamydia trachomatis Infection through Rapid and Specific Host Cell Apoptosis. International Journal of Molecular Sciences. 2022; 23(13):7434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137434
Chicago/Turabian StyleItoh, Ryota, Yusuke Kurihara, Michinobu Yoshimura, and Kenji Hiromatsu. 2022. "Bortezomib Eliminates Persistent Chlamydia trachomatis Infection through Rapid and Specific Host Cell Apoptosis" International Journal of Molecular Sciences 23, no. 13: 7434. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23137434