
Genetic Variation in the MBL2 Gene Is Associated with Chlamydia trachomatis Infection and Host Humoral Response to Chlamydia trachomatis Infection

 , ,
, ,
Abstract
:1. Introduction
2. Results
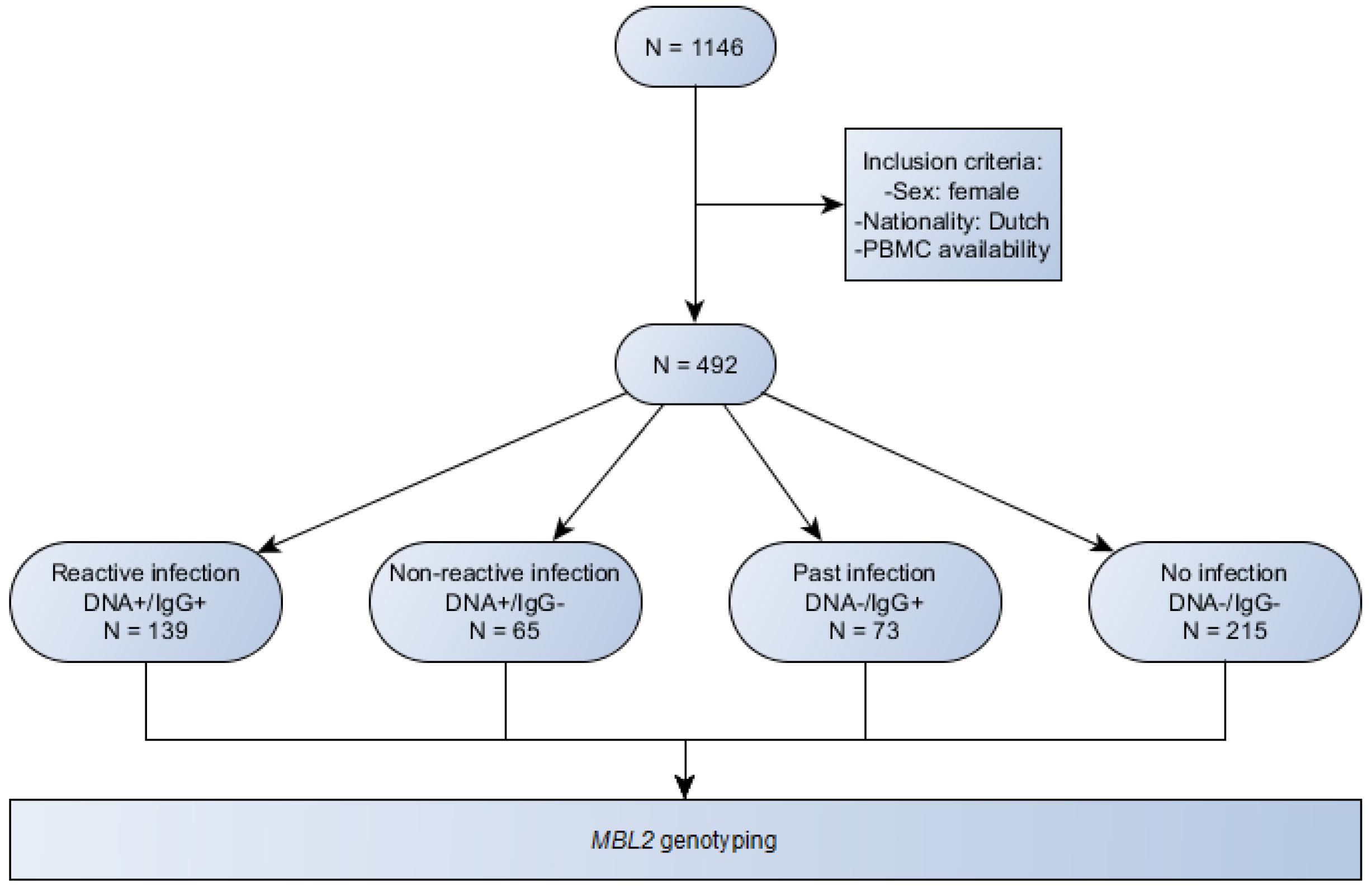
2.1. Study Sample Characteristics
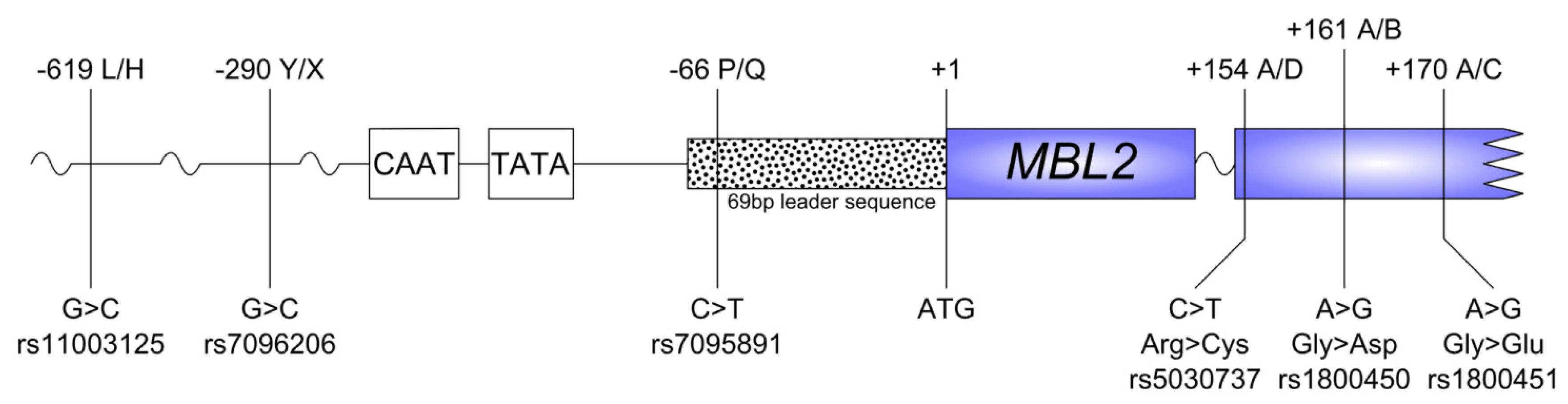
2.2. SNP Distribution
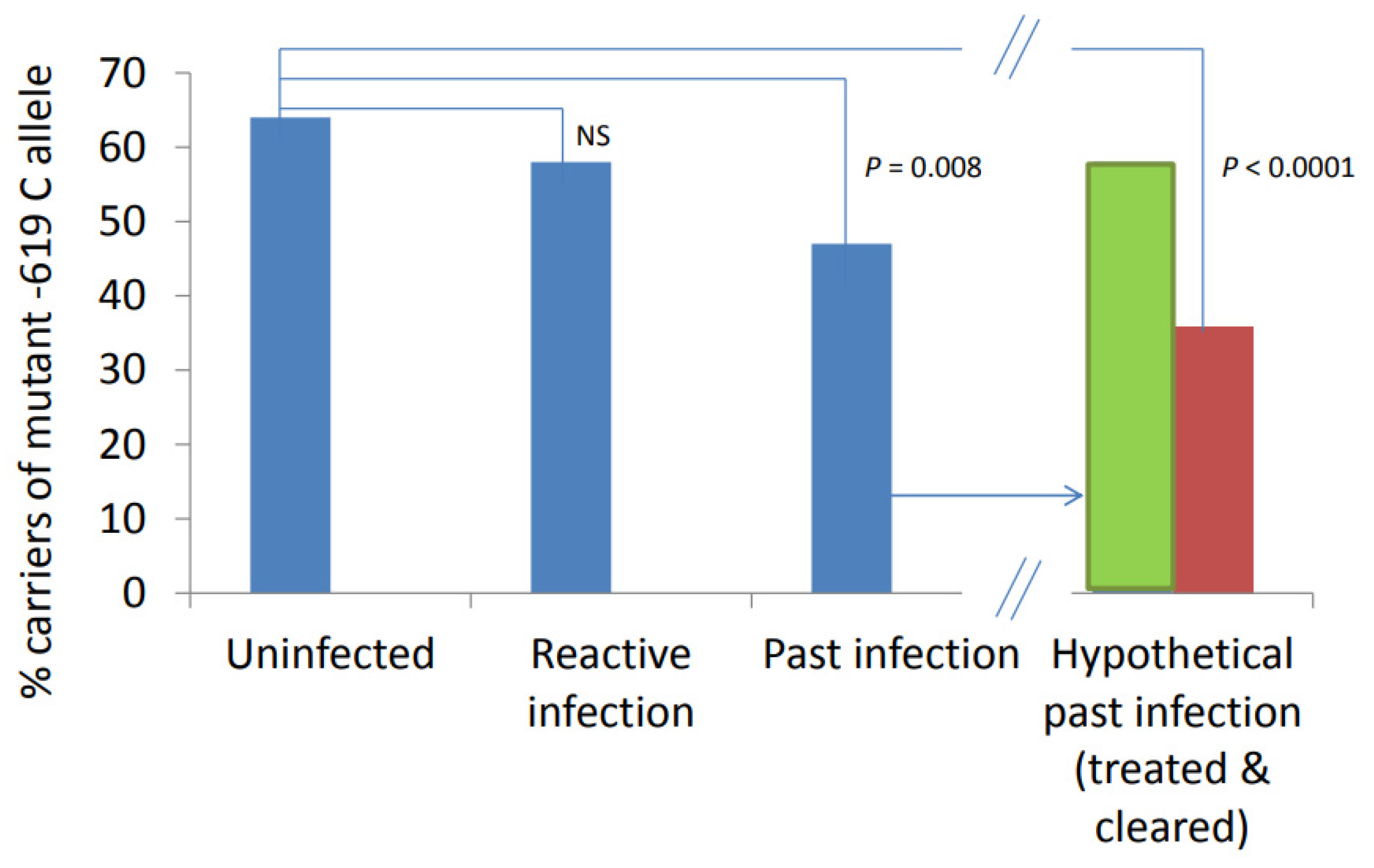
2.3. Association of SNPs and Stadium of Infection
2.4. Effect of Homozygous and Heterozygous Carriage of Alleles and Susceptibility to Infection
2.5. Haplotype Frequencies and Associations with Susceptibility
2.6. Association of SNPs with Immune Response
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Ethical Approval
4.3. Laboratory Tests
4.4. Genotype Analysis
4.5. Subgroups for Analyses
4.6. Statistical Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| SNP | single-nucleotide polymorphism |
| MBL | mannose-binding lectin |
| IgG | immunoglobulin G |
| ELISA | enzyme-linked immunosorbent assay |
| PCR | polymerase chain reaction |
| OR | odds ratio |
| CI | confidence interval |
References
- Morré, S.A.; van den Brule, A.J.; Rozendaal, L.; Boeke, A.J.; Voorhorst, F.J.; de Blok, S.; Meijer, C.J.L.M. The natural course of asymptomatic Chlamydia trachomatis infections: 45% clearance and no development of clinical PID after one-year follow-up. Int. J. STD AIDS 2002, 13 (Suppl. S2), 12–18. [Google Scholar] [CrossRef] [PubMed]
- Janeway, C.A., Jr.; Medzhitov, R. Innate immune recognition. Annu. Rev. Immunol. 2002, 20, 197–216. [Google Scholar] [CrossRef]
- Super, M.; Thiel, S.; Lu, J.; Levinsky, R.J.; Turner, M.W. Association of low levels of mannan-binding protein with a common defect of opsonisation. Lancet 1989, 2, 1236–1239. [Google Scholar] [CrossRef]
- Swanson, A.F.; Ezekowitz, R.A.; Lee, A.; Kuo, C.C. Human mannose-binding protein inhibits infection of HeLa cells by Chlamydia trachomatis. Infect. Immun. 1998, 66, 1607–1612. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, D.C. Mannan-binding lectin and its role in innate immunity. Transfus. Med. 2002, 12, 335–352. [Google Scholar] [CrossRef]
- Thiel, S.; Vorup-Jensen, T.; Stover, C.M.; Schwaeble, W.; Laursen, S.B.; Poulsen, K.; Willis, A.C.; Eggleton, P.; Hansen, S.; Holmskov, U.; et al. A second serine protease associated with mannan-binding lectin that activates complement. Nature 1997, 386, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Turner, M.W. Mannose-binding lectin (MBL) in health and disease. Immunobiology 1998, 199, 327–339. [Google Scholar] [CrossRef]
- Garred, P.; Honore, C.; Ma, Y.J.; Munthe-Fog, L.; Hummelshoj, T. MBL2, FCN1, FCN2 and FCN3-The genes behind the initiation of the lectin pathway of complement. Mol. Immunol. 2009, 46, 2737–2744. [Google Scholar] [CrossRef]
- Taylor, M.E.; Brickell, P.M.; Craig, R.K.; Summerfield, J.A. Structure and evolutionary origin of the gene encoding a human serum mannose-binding protein. Biochem. J. 1989, 262, 763–771. [Google Scholar] [CrossRef] [PubMed]
- Sastry, K.; Herman, G.A.; Day, L.; Deignan, E.; Bruns, G.; Morton, C.C.; Ezekowitz, R.A. The human mannose-binding protein gene. Exon structure reveals its evolutionary relationship to a human pulmonary surfactant gene and localization to chromosome 10. J. Exp. Med. 1989, 170, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Madsen, H.O.; Garred, P.; Thiel, S.; Kurtzhals, J.A.; Lamm, L.U.; Ryder, L.P.; Svejgaard, A. Interplay between promoter and structural gene variants control basal serum level of mannan-binding protein. J. Immunol. 1995, 155, 3013–3020. [Google Scholar] [PubMed]
- Boldt, A.B.; Petzl-Erler, M.L. A new strategy for mannose-binding lectin gene haplotyping. Hum. Mutat. 2002, 19, 296–306. [Google Scholar] [CrossRef] [PubMed]
- Steffensen, R.; Thiel, S.; Varming, K.; Jersild, C.; Jensenius, J.C. Detection of structural gene mutations and promoter polymorphisms in the mannan-binding lectin (MBL) gene by polymerase chain reaction with sequence-specific primers. J. Immunol. Methods 2000, 241, 33–42. [Google Scholar] [CrossRef]
- Madsen, H.O.; Satz, M.L.; Hogh, B.; Svejgaard, A.; Garred, P. Different molecular events result in low protein levels of mannan-binding lectin in populations from southeast Africa and South America. J. Immunol. 1998, 161, 3169–3175. [Google Scholar] [PubMed]
- Yokota, Y.; Arai, T.; Kawasaki, T. Oligomeric structures required for complement activation of serum mannan-binding proteins. J. Biochem. 1995, 117, 414–419. [Google Scholar] [CrossRef] [PubMed]
- Wallis, R. Dominant effects of mutations in the collagenous domain of mannose-binding protein. J. Immunol. 2002, 168, 4553–4558. [Google Scholar] [CrossRef]
- Eisen, D.P.; Minchinton, R.M. Impact of mannose-binding lectin on susceptibility to infectious diseases. Clin. Infect. Dis. 2003, 37, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Mayilyan, K.R. Complement genetics, deficiencies, and disease associations. Protein Cell 2012, 3, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Hibberd, M.L.; Sumiya, M.; Summerfield, J.A.; Booy, R.; Levin, M. Association of variants of the gene for mannose-binding lectin with susceptibility to meningococcal disease. Meningococcal Research Group. Lancet 1999, 353, 1049–1053. [Google Scholar] [CrossRef]
- Thye, T.; Niemann, S.; Walter, K.; Homolka, S.; Intemann, C.D.; Chinbuah, M.A.; Enimil, A.; Gyapong, J.; Osei, I.; Owusu-Dabo, E.; et al. Variant G57E of mannose binding lectin associated with protection against tuberculosis caused by Mycobacterium africanum but not by M. tuberculosis. PLoS ONE 2011, 6, e20908. [Google Scholar] [CrossRef] [PubMed]
- Darville, T.; Hiltke, T.J. Pathogenesis of genital tract disease due to Chlamydia trachomatis. J. Infect. Dis. 2010, 201 (Suppl. S2), S114–S125. [Google Scholar] [CrossRef]
- Sziller, I.; Babula, O.; Ujhazy, A.; Nagy, B.; Hupuczi, P.; Papp, Z.; Linhares, I.M.; Ledger, W.J.; Witkin, S.S. Chlamydia trachomatis infection, Fallopian tube damage and a mannose-binding lectin codon 54 gene polymorphism. Hum. Reprod. 2007, 22, 1861–1865. [Google Scholar] [CrossRef] [PubMed]
- Laisk, T.; Peters, M.; Saare, M.; Haller-Kikkatalo, K.; Karro, H.; Salumets, A. Association of CCR5, TLR2, TLR4 and MBL genetic variations with genital tract infections and tubal factor infertility. J. Reprod. Immunol. 2010, 87, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Laisk, T.; Peters, M.; Salumets, A. Mannose-binding lectin genotypes: Potential role in tubal damage and adverse IVF outcome. J. Reprod. Immunol. 2011, 92, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Roos, A.; Garred, P.; Wildenberg, M.E.; Lynch, N.J.; Munoz, J.R.; Zuiverloon, T.C.; Bouwman, L.H.; Schlagwein, N.; van den Houten, F.C.F.; Faber-Krol, M.C.; et al. Antibody-mediated activation of the classical pathway of complement may compensate for mannose-binding lectin deficiency. Eur. J. Immunol. 2004, 34, 2589–2598. [Google Scholar] [CrossRef] [PubMed]
- Garred, P.; Harboe, M.; Oettinger, T.; Koch, C.; Svejgaard, A. Dual role of mannan-binding protein in infections: Another case of heterosis? Eur. J. Immunogenet. 1994, 21, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Verdu, P.; Barreiro, L.B.; Patin, E.; Gessain, A.; Cassar, O.; Kidd, J.R.; Kidd, K.K.; Behar, D.M.; Froment, A.; Heyer, E.; et al. Evolutionary insights into the high worldwide prevalence of MBL2 deficiency alleles. Hum. Mol. Genet. 2006, 15, 2650–2658. [Google Scholar] [CrossRef] [PubMed]
- Öhman, H.; Natividad, A.; Bailey, R.; Ragoussis, J.; Johnson, L.L.; Tiitinen, A.; Halttunen, M.; Paavonen, J.; Surcel, H.-M. Contribution of IL-12A and IL-12B polymorphisms to Chlamydia trachomatis-specific cell-mediated immune responses. Scand. J. Immunol. 2015, 81, 209–213. [Google Scholar] [CrossRef] [PubMed]
- Catsburg, A.; van der Zwet, W.C.; Morré, S.A.; Ouburg, S.; Vandenbroucke-Grauls, C.M.; Savelkoul, P.H. Analysis of multiple single nucleotide polymorphisms (SNP) on DNA traces from plasma and dried blood samples. J. Immunol. Methods 2007, 321, 135–141. [Google Scholar] [CrossRef]
- Verweij, S.P.; Bax, C.J.; Quint, K.D.; Quint, W.G.; van Leeuwen, A.P.; Peters, R.P.; Oostvogel, P.M.; Mutsaers, J.A.; Dörr, P.J.; Pleijster, J.; et al. Significantly higher serologic responses of Chlamydia trachomatis B group serovars versus C and I serogroups. Drugs Today 2009, 45 (Suppl. B), 135–140. [Google Scholar]
- Verweij, S.P.; Lanjouw, E.; Bax, C.J.; Quint, K.D.; Oostvogel, P.M.; Dorr, P.J.; Pleijster, J.; de Vries, H.J.C.; Peters, R.P.H.; Ouburg, S.; et al. Serovar D and E of serogroup B induce highest serological responses in urogenital Chlamydia trachomatis infections. BMC Infect. Dis. 2014, 14, 3. [Google Scholar] [CrossRef] [PubMed]
- Burke, W.; Khoury, M.J.; Stewart, A.; Zimmern, R.L. The path from genome-based research to population health: Development of an international public health genomics network. Genet. Med. 2006, 8, 451–458. [Google Scholar] [CrossRef] [PubMed]
- Lal, J.A.; Malogajski, J.; Verweij, S.P.; de Boer, P.; Ambrosino, E.; Brand, A.; Ouburg, S.; Morré, S.A. Chlamydia trachomatis Infections and Subfertility: Opportunities to Translate Host Pathogen Genomic Data into Public Health. Public Health Genom. 2013, 16, 50–61. [Google Scholar] [CrossRef] [PubMed]
- Haasnoot, A.; Koedijk, F.D.; Op de Coul, E.L.M.; Gotz, H.M.; van der Sande, M.A.; van den Broek, I.V. Comparing two definitions of ethnicity for identifying young persons at risk for chlamydia. Epidemiol. Infect. 2012, 140, 951–958. [Google Scholar] [CrossRef] [PubMed]
- Ouburg, S.; Spaargaren, J.; den Hartog, J.E.; Land, J.A.; Fennema, J.S.; Pleijster, J.; Peña, A.S.; Morré, S.A. The CD14 functional gene polymorphism -260 C>T is not involved in either the susceptibility to Chlamydia trachomatis infection or the development of tubal pathology. BMC Infect. Dis. 2005, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- van der Zwet, W.C.; Catsburg, A.; van Elburg, R.M.; Savelkoul, P.H.; Vandenbroucke-Grauls, C.M. Mannose-binding lectin (MBL) genotype in relation to risk of nosocomial infection in pre-term neonates in the neonatal intensive care unit. Clin. Microbiol. Infect. 2008, 14, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Smith, N.J.; Donnelly, P. A new statistical method for haplotype reconstruction from population data. Am. J. Hum. Genet. 2001, 68, 978–989. [Google Scholar] [CrossRef] [PubMed]
- Stephens, M.; Donnelly, P. A comparison of bayesian methods for haplotype reconstruction from population genotype data. Am. J. Hum. Genet. 2003, 73, 1162–1169. [Google Scholar] [CrossRef] [PubMed]
- David, G. Clayton. SNPHAP. 2002. Available online: https://mybiosoftware.com/snphap-1-3-1-estimate-frequencies-haplotypes-snps.html (accessed on 17 February 2022).
- Rothman, K.J. No adjustments are needed for multiple comparisons. Epidemiology 1990, 1, 43–46. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Chlamydia trachomatis Infection | ||||||
|---|---|---|---|---|---|---|
| Overall SNP Distribution | Ct-DNA+/IgG+ | Ct-DNA+/IgG− | Ct-DNA−/IgG+ | Ct-DNA−/IgG− | ||
| n = 492 (%) | n = 139 (%) | n = 65 (%) | n = 73 (%) | n = 215 (%) | ||
| −619 G > C | GG | 203 (41) | 59 (42) | 28 (43) | 39 (53) * | 77 (36) |
| GC | 224 (46) | 67 (48) | 30 (46) | 24 (33) | 103 (48) | |
| CC | 65 (13) | 13 (9.4) | 7 (11) | 10 (14) | 35 (16) | |
| −290 G > C | GG | 301 (61) | 88 (63) | 35 (54) | 41 (56) | 137 (64) |
| GC | 167 (34) | 45 (32) | 29 (45) | 27 (37) | 66 (31) | |
| CC | 24 (4.9) | 6 (4.3) | 1 (1.5) | 5 (6.8) | 12 (5.6) | |
| −66 C > T | CC | 280 (57) | 76 (55) | 36 (55) | 41 (56) | 127 (59) |
| CT | 187 (38) | 51 (37) | 27 (42) | 28 (38) | 81 (38) | |
| TT | 25 (5.1) | 12 (8.6) | 2 (3.1) | 4 (5.5) | 7 (3.3) | |
| +154 C > T | CC | 430 (87) | 122 (88) | 58 (89) | 63 (86) | 187 (87) |
| CT | 60 (12) | 15 (11) | 7 (11) | 10 (14) | 28 (13) | |
| TT | 2 (0.4) | 2 (1.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| +161 A > G | AA | 386 (78) | 108 (78) | 49 (75) | 57 (78) | 172 (80) |
| AG | 99 (20) | 28 (20) | 15 (23) | 15 (21) | 41 (19) | |
| GG | 7 (1.4) | 3 (2.2) | 1 (1.5) | 1 (1.4) | 2 (0.9) | |
| +170 A > G | AA | 459 (93) | 123 (89) | 63 (97) | 69 (95) | 204 (95) |
| AG | 32 (7) | 15 (11) | 2 (3.1) | 4 (5.5) | 11 (5.1) | |
| GG | 1 (0.2) | 1 (0.7) | 0 (0.0) | 0 (0.0) | 0 (0.0) | |
| Ct-DNA+/IgG+ | Ct-DNA+/IgG− | Ct-DNA−/IgG+ | Ct-DNA−/IgG− | ||
|---|---|---|---|---|---|
| MBL Production | n = 139 (%) | n = 65 (%) | n = 73 (%) | n = 215 (%) | |
| HYA/HYA | High | 7 (5.0) * | 4 (6.2) | 6 (8.2) | 27 (13) |
| HYA/LYA | 22 (16) | 12 (19) | 8 (11) | 37 (17) | |
| HYA/LXA | 19 (14) | 8 (12) | 5 (6.8) | 26 (12) | |
| LYA/LYA | 10 (7.2) | 2 (3.1) | 8 (11) | 13 (6.0) | |
| LYA/LXA | 14 (10) | 12 (19) | 13 (18) | 23 (11) | |
| LXA/LXA | Low | 6 (4.3) | 1 (1.5) | 5 (6.8) | 12 (5.6) |
| HYA/O | 19 (14) | 9 (14) | 9 (12) | 28 (13) | |
| LYA/O | 21 (15) | 7 (11) | 7 (9.6) | 25 (12) | |
| LXA/O | Deficient | 12 (8.6) | 9 (14) | 9 (12) | 17 (7.9) |
| O/O | 9 (6.5) | 1 (1.5) | 3 (4.1) | 7 (3.3) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Verweij, S.P.; Peters, R.P.H.; Catsburg, A.; de Vries, H.J.C.; Ouburg, S.; Morré, S.A. Genetic Variation in the MBL2 Gene Is Associated with Chlamydia trachomatis Infection and Host Humoral Response to Chlamydia trachomatis Infection. Int. J. Mol. Sci. 2022, 23, 9292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169292
Verweij SP, Peters RPH, Catsburg A, de Vries HJC, Ouburg S, Morré SA. Genetic Variation in the MBL2 Gene Is Associated with Chlamydia trachomatis Infection and Host Humoral Response to Chlamydia trachomatis Infection. International Journal of Molecular Sciences. 2022; 23(16):9292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169292
Chicago/Turabian StyleVerweij, Stephan P., Remco P. H. Peters, Arnold Catsburg, Henry J. C. de Vries, Sander Ouburg, and Servaas A. Morré. 2022. "Genetic Variation in the MBL2 Gene Is Associated with Chlamydia trachomatis Infection and Host Humoral Response to Chlamydia trachomatis Infection" International Journal of Molecular Sciences 23, no. 16: 9292. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169292