
GhCYP710A1 Participates in Cotton Resistance to Verticillium Wilt by Regulating Stigmasterol Synthesis and Plasma Membrane Stability

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effects of Verticillium dahliae Infection on Sterol Content and Gene Expression in Cotton Roots
2.2. Characterization of GhCYP710A1 in Upland Cotton
2.3. Expression Pattern and Subcellular Location of GhCYP710A1
2.4. Ectopic Expression of GhCYP710A1 Gene Promoted the Synthesis of Stigmasterol in Arabidopsis thaliana
2.5. Ectopic Expression of GhCYP710A1 Gene Enhanced the Resistance to V991 in Arabidopsis
2.6. The Transgenic Cotton Plants Overexpressing GhCYP710A1 Enhanced the Resistance to V991
2.7. Silencing GhCYP710A1 Reduced Cotton Resistance to V991
2.8. Modifying GhCYP710A1 Expression Resulted in the Alteration of Plasma Membrane Permeability and ROS Accumulation in Transgenic Cotton Plants
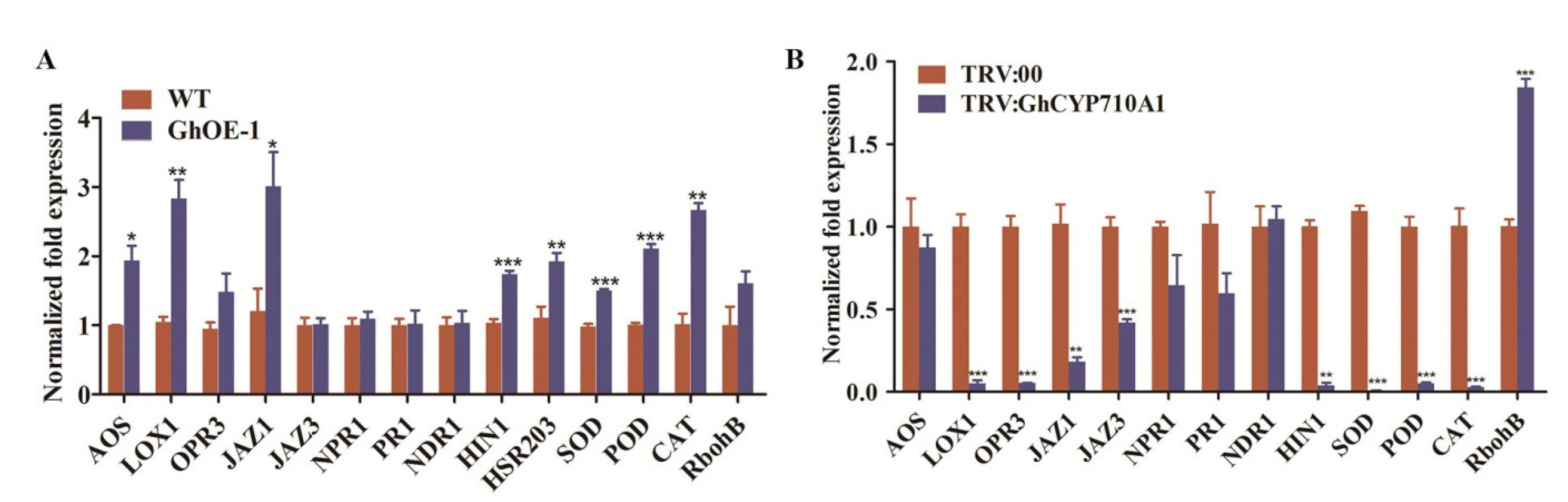
2.9. Modifying GhCYP710A1 Expression Led to the Expression Changes of Disease Resistance Genes
3. Discussion
3.1. GhCYP710A1 and Stigmasterol Positively Regulated Cotton Resistance to Verticillium Wilt
3.2. Effect of Membrane Lipid Changes on the Infection Process of Verticillium dahliae
3.3. GhCYP710A1 Regulates Cotton Resistance to Verticillium Wilt by Regulating ROS, HR and JA Signaling Pathways
4. Materials and Methods
4.1. Plant and Verticillium dahliae Materials and Growth Conditions
4.2. Subcellular Localization of GhCYP710A1
4.3. Vectors Construction and Transformation
4.4. RNA Extraction and qRT-PCR Assay
4.5. Investigation of Plant Resistance to Verticillium dahliae
4.6. Trypan Blue Staining
4.7. Reactive Oxygen Species (ROS) Detection in Cotton Leaves
4.8. Verticillium Dahliae Recovery Assay
4.9. Di-4-ANEPPDHQ Stain and Observation
4.10. GUS Histochemical Staining
4.11. Determination of Relative Conductivity
4.12. Sample Collection, Lipid Extraction and Lipidomics
4.13. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Inderbitzin, P.; Bostock, R.M.; Davis, R.M.; Usami, T.; Platt, H.W.; Subbarao, K.V. Phylogenetics and Taxonomy of the Fungal Vascular Wilt Pathogen Verticillium, with the Descriptions of Five New Species. PLoS ONE 2011, 6, e28341. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk-Fradin, E.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef] [PubMed]
- Kong, G.A. Plant Pathology 4th edition. Australas Plant Pathol. 1997, 26, 133. [Google Scholar] [CrossRef]
- Inderbitzin, P.; Subbarao, K.V. Verticillium Systematics and Evolution: How Confusion Impedes Verticillium Wilt Management and How to Resolve It. Phytopathology 2014, 104, 564–574. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fang, H.; Zhou, H.; Sanogo, S.; Ma, Z. Genetics, Breeding, and Marker-Assisted Selection for Verticillium Wilt Resistance in Cotton. Crop. Sci. 2014, 54, 1289–1303. [Google Scholar] [CrossRef]
- Vallad, G.E.; Qin, Q.-M.; Grube, R.; Hayes, R.J.; Subbarao, K.V. Characterization of Race-Specific Interactions Among Isolates of Verticillium dahliae Pathogenic on Lettuce. Phytopathology 2006, 96, 1380–1387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, Pathogenicity, and Management of Verticillium Species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [Green Version]
- Abdelraheem, A.; Esmaeili, N.; O’Connell, M.; Zhang, J. Progress and perspective on drought and salt stress tolerance in cotton. Ind. Crop. Prod. 2018, 130, 118–129. [Google Scholar] [CrossRef]
- Qin, J.; Wang, K.; Sun, L.; Xing, H.; Wang, S.; Li, L.; Chen, S.; Guo, H.-S.; Zhang, J. The plant-specific transcription factors CBP60g and SARD1 are targeted by a Verticillium secretory protein VdSCP41 to modulate immunity. eLife 2018, 7, e34902. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liang, C.; Wu, S.; Zhang, X.; Tang, J.; Jian, G.; Jiao, G.; Li, F.; Chu, C.J. Significant Improvement of Cotton Verticillium Wilt Resistance by Manipulating the Expression of Gastrodia Antifungal Proteins—ScienceDirect. Mol. Plant 2016, 9, 1436–1439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Zhao, Y.-L.; Jin, Y.; Zhang, T.; Guo, H.-S. Colonization process of Arabidopsis thaliana roots by a green fluorescent protein-tagged isolate of Verticillium dahliae. Protein Cell 2014, 5, 94–98. [Google Scholar] [CrossRef]
- Zhou, T.-T.; Zhao, Y.-L.; Guo, H.-S. Secretory proteins are delivered to the septin-organized penetration interface during root infection by Verticillium dahliae. PLoS Pathog. 2017, 13, e1006275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Xie, C.; Dong, J.; Yang, X.; Sui, A. Interactions between Verticillium dahliae and its host: Vegetative growth, pathogenicity, plant immunity. Appl. Microbiol. Biotechnol. 2014, 98, 6921–6932. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.-D.; Wang, X.-Y.; Chen, J.-Y.; Kong, Z.-Q.; Gui, Y.-J.; Li, N.-Y.; Bao, Y.-M.; Dai, X.-F. Identification and characterization of a pathogenicity-related gene VdCYP1 from Verticillium dahliae. Sci. Rep. 2016, 6, 27979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Huang, L.; Wang, J.; Ma, C.; Tan, Y.; Wang, F.; Fan, Y.; Luo, M. Sphingolipid synthesis inhibitor fumonisin B1 causes verticillium wilt in cotton. J. Integr. Plant Biol. 2022, 64, 836–842. [Google Scholar] [CrossRef] [PubMed]
- Valitova, J.N.; Sulkarnayeva, A.G.; Minibayeva, F.V. Plant sterols: Diversity, biosynthesis, and physiological functions. Biochemistry 2016, 81, 819–834. [Google Scholar] [CrossRef]
- Breslow, D.K.; Weissman, J.S. Membranes in Balance: Mechanisms of Sphingolipid Homeostasis. Mol. Cell 2010, 40, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Cacas, J.-L.; Buré, C.; Grosjean, K.; Gerbeau-Pissot, P.; Lherminier, J.; Rombouts, Y.; Maes, E.; Bossard, C.; Gronnier, J.; Furt, F.; et al. Revisiting Plant Plasma Membrane Lipids in Tobacco: A Focus on Sphingolipids. Plant Physiol. 2015, 170, 367–384. [Google Scholar] [CrossRef]
- Simon-Plas, F.; Perraki, A.; Bayer, E.; Gerbeau-Pissot, P.; Mongrand, S. An update on plant membrane rafts. Curr. Opin. Plant Biol. 2011, 14, 642–649. [Google Scholar] [CrossRef]
- Grosjean, K.; Mongrand, S.; Beney, L.; Simon-Plas, F.; Gerbeau-Pissot, P. Differential Effect of Plant Lipids on Membrane Organization: Specificities of phytosphingolipids and phytosterols. J. Biol. Chem. 2015, 290, 5810–5825. [Google Scholar] [CrossRef] [Green Version]
- Grunwald, C. Effects of Free Sterols, Steryl Ester, and Steryl Glycoside on Membrane Permeability. Plant Physiol. 1971, 48, 653–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, M.-A. Plant sterols and the membrane environment. Trends Plant Sci. 1998, 3, 170–175. [Google Scholar] [CrossRef]
- Hodzic, A.; Rappolt, M.; Amenitsch, H.; Laggner, P.; Pabst, G. Differential Modulation of Membrane Structure and Fluctuations by Plant Sterols and Cholesterol. Biophys. J. 2008, 94, 3935–3944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, P.; Hirsch-Hoffmann, M.; Hennig, L.; Gruissem, W. GENEVESTIGATOR. Arabidopsis Microarray Database and Analysis Toolbox. Plant Physiol. 2004, 136, 2621–2632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabro, G.; Di Rienzo, J.A.; Voigt, C.A.; Savchenko, T.; Dehesh, K.; Somerville, S.; Alvarez, M.E. Genome-Wide Expression Profiling Arabidopsis at the Stage of Golovinomyces cichoracearum Haustorium Formation. Plant Physiol. 2008, 146, 1421–1439. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Senthil-Kumar, M.; Ryu, C.-M.; Kang, L.; Mysore, K.S. Phytosterols Play a Key Role in Plant Innate Immunity against Bacterial Pathogens by Regulating Nutrient Efflux into the Apoplast. Plant Physiol. 2012, 158, 1789–1802. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Nagegowda, D.A.; Rawat, R.; Bouvier-Navé, P.; Guo, D.; Bach, T.J.; Chye, M.-L. Overexpression of Brassica juncea wild-type and mutant HMG-CoA synthase 1 in Arabidopsis up-regulates genes in sterol biosynthesis and enhances sterol production and stress tolerance. Plant Biotechnol. J. 2011, 10, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Gamir, J.; Darwiche, R.; Hof, P.V.; Choudhary, V.; Stumpe, M.; Schneiter, R.; Mauch, F. The sterol-binding activity of PATHOGENESIS-RELATED PROTEIN 1 reveals the mode of action of an antimicrobial protein. Plant J. 2017, 89, 502–509. [Google Scholar] [CrossRef] [Green Version]
- Melotto, M.; Underwood, W.; He, S.Y. Role of Stomata in Plant Innate Immunity and Foliar Bacterial Diseases. Annu. Rev. Phytopathol. 2008, 46, 101–122. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.-Q.; Hou, B.-H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.-Q.; Guo, W.-J.; Kim, J.-G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef] [Green Version]
- Simons, K.; Sampaio, J.L. Membrane Organization and Lipid Rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, a004697. [Google Scholar] [CrossRef] [PubMed]
- Rogowska, A.; Szakiel, A. The role of sterols in plant response to abiotic stress. Phytochem. Rev. 2020, 19, 1525–1538. [Google Scholar] [CrossRef]
- Niu, Q.; Tan, K.; Zang, Z.; Xiao, Z.; Chen, K.; Hu, M.; Luo, M. Modification of phytosterol composition influences cotton fiber cell elongation and secondary cell wall deposition. BMC Plant Biol. 2019, 19, 208. [Google Scholar] [CrossRef] [PubMed]
- Gotoh, O. Substrate recognition sites in cytochrome P450 family 2 (CYP2) proteins inferred from comparative analyses of amino acid and coding nucleotide sequences. J. Biol. Chem. 1992, 267, 83–90. [Google Scholar] [CrossRef]
- Meunier, B.; de Visser, S.P.; Shaik, S. Mechanism of Oxidation Reactions Catalyzed by Cytochrome P450 Enzyme. ChemInform 2004, 104, 3947–3980. [Google Scholar] [CrossRef]
- Browse, J. Jasmonate Passes Muster: A Receptor and Targets for the Defense Hormone. Annu. Rev. Plant Biol. 2009, 60, 183–205. [Google Scholar] [CrossRef]
- Ellendorff, U.; Fradin, E.F.; de Jonge, R.; Thomma, B.P.H.J. RNA silencing is required for Arabidopsis defence against Verticillium wilt disease. J. Exp. Bot. 2008, 60, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Gayoso, C.; Pomar, F.; Novo-Uzal, E.; Merino, F.; de Ilárduya, Ó.M. The Ve-mediated resistance response of the tomato to Verticillium dahliae involves H2O2, peroxidase and lignins and drives PALgene expression. BMC Plant Biol. 2010, 10, 232. [Google Scholar] [CrossRef] [Green Version]
- Jambunathan, N. Determination and Detection of Reactive Oxygen Species (ROS), Lipid Peroxidation, and Electrolyte Leakage in Plants. Methods Mol. Biol. 2010, 639, 291–297. [Google Scholar]
- Bhat, R.A.; Panstruga, R. Lipid rafts in plants. Planta 2005, 223, 5–19. [Google Scholar] [CrossRef]
- Lindsey, K.; Pullen, M.L.; Topping, J.F. Importance of plant sterols in pattern formation and hormone signalling. Trends Plant Sci. 2003, 8, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.L.; Ueda, T.; Hara-Nishimura, I. Excess sterol accumulation affects seed morphology and physiology in Arabidopsis thaliana. Plant Signal. Behav. 2021, 16, 1872217. [Google Scholar] [CrossRef] [PubMed]
- Vriet, C.; Russinova, E.; Reuzeau, C. From Squalene to Brassinolide: The Steroid Metabolic and Signaling Pathways across the Plant Kingdom. Mol. Plant 2013, 6, 1738–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schrick, K.; Fujioka, S.; Takatsuto, S.; Stierhof, Y.-D.; Stransky, H.; Yoshida, S.; Jürgens, G. A link between sterol biosynthesis, the cell wall, and cellulose in Arabidopsis. Plant J. 2004, 38, 227–243. [Google Scholar] [CrossRef] [PubMed]
- Benveniste, P. Biosynthesis and accumulation of sterols. Annu. Rev. Plant Biol. 2004, 55, 429–457. [Google Scholar] [CrossRef]
- Peng, L.; Kawagoe, Y.; Hogan, P.; Delmer, D. Sitosterol-β-glucoside as Primer for Cellulose Synthesis in Plants. Science 2002, 295, 147–150. [Google Scholar] [CrossRef]
- Xin, X.-F.; Kvitko, B.; He, S.Y. Pseudomonas syringae: What it takes to be a pathogen. Nat. Rev. Genet. 2018, 16, 316–328. [Google Scholar] [CrossRef] [PubMed]
- Griebel, T.; Zeier, J. A role for β-sitosterol to stigmasterol conversion in plant-pathogen interactions. Plant J. 2010, 63, 254–268. [Google Scholar] [CrossRef]
- Malinsky, J.; Opekarová, M.; Grossmann, G.; Tanner, W. Membrane Microdomains, Rafts, and Detergent-Resistant Membranes in Plants and Fungi. Annu. Rev. Plant Biol. 2013, 64, 501–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerbeau-Pissot, P.; Der, C.; Thomas, D.; Anca, I.-A.; Grosjean, K.; Roche, Y.; Perrier-Cornet, J.M.; Mongrand, S.; Simon-Plas, F. Modification of Plasma Membrane Organization in Tobacco Cells Elicited by Cryptogein. Plant Physiol. 2014, 164, 273–286. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Wang, C.; Wang, X.; Yang, X. Proteomics-based analysis reveals that Verticillium dahliae toxin induces cell death by modifying the synthesis of host proteins. J. Gen. Plant Pathol. 2013, 79, 335–345. [Google Scholar] [CrossRef] [Green Version]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. [Google Scholar] [CrossRef]
- Brisson, L.F.; Tenhaken, R.; Lamb, C. Function of Oxidative Cross-Linking of Cell Wall Structural Proteins in Plant Disease Resistance. Plant Cell 1994, 6, 1703–1712. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Malamy, J.; Henning, J.; Conrath, U.; Sánchez-Casas, P.; Silva, H.; Ricigliano, J.; Klessig, D.K. Induction, modification, and transduction of the salicylic acid signal in plant defense responses. Proc. Natl. Acad Sci. USA 1995, 92, 4134–4137. [Google Scholar] [CrossRef] [Green Version]
- Peng, M. Peroxidase-Generated Hydrogen Peroxide as a Source of Antifungal Activity In Vitro and on Tobacco Leaf Disks. Phytopathology 1992, 82, 696–699. [Google Scholar] [CrossRef]
- Greenberg, J.T.; Yao, N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell. Microbiol. 2004, 6, 201–211. [Google Scholar] [CrossRef]
- Mittler, R.; Vanderauwera, S.; Gollery, M.; Van Breusegem, F. Reactive oxygen gene network of plants. Trends Plant Sci. 2004, 9, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Mittler, R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Kazan, K.; Lyons, R. Intervention of Phytohormone Pathways by Pathogen Effectors. Plant Cell 2014, 26, 2285–2309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimova, O.; Shchennikova, A.; Kochieva, E.; Filyushin, M. Pathogenesis-Related Genes of PR1, PR2, PR4, and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021, 22, 6688. [Google Scholar] [CrossRef]
- Overmyer, K.; Tuominen, H.; Kettunen, R.; Betz, C.; Langebartels, C.; Sandermann, H., Jr.; Kangasjärvi, J. Ozone-Sensitive Arabidopsis rcd1 Mutant Reveals Opposite Roles for Ethylene and Jasmonate Signaling Pathways in Regulating Superoxide-Dependent Cell Death. Plant Cell 2000, 12, 1849–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovács, J.; Poór, P.; Szepesi, Á.; Tari, I. Salicylic acid induced cysteine protease activity during programmed cell death in tomato plants. Acta Biol. Hung. 2016, 67, 148–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Díaz, R.M.; Olivares-García, C.; Landa, B.B.; Jiménez-Gasco, M.D.M.; Navas-Cortés, J.A. Region-Wide Analysis of Genetic Diversity in Verticillium dahliae Populations Infecting Olive in Southern Spain and Agricultural Factors Influencing the Distribution and Prevalence of Vegetative Compatibility Groups and Pathotypes. Phytopathology 2011, 101, 304–315. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.-P.; Gurung, S.; Short, D.P.G.; Sandoya, G.V.; Shang, W.-J.; Hayes, R.J.; Davis, R.M.; Subbarao, K.V. Nondefoliating and Defoliating Strains from Cotton Correlate with Races 1 and 2 of Verticillium dahliae. Plant Dis. 2015, 99, 1713–1720. [Google Scholar] [CrossRef] [Green Version]
- Wroblewski, T.; Tomczak, A.; Michelmore, R. Optimization of Agrobacterium-mediated transient assays of gene expression in lettuce, tomato and Arabidopsis. Plant Biotechnol. J. 2005, 3, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Suo, X.; Li, F.; Bao, C.; He, S.; Huang, L.; Luo, M. Membrane lipid raft organization during cotton fiber development. J. Cotton Res. 2020, 3, 13. [Google Scholar] [CrossRef]
- Wang, Q.; Meng, Q.; Xu, F.; Chen, Q.; Ma, C.; Huang, L.; Li, G.; Luo, M. Comparative Metabolomics Analysis Reveals Sterols and Sphingolipids Play a Role in Cotton Fiber Cell Initiation. Int. J. Mol. Sci. 2021, 22, 11438. [Google Scholar] [CrossRef] [PubMed]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, L.; Li, G.; Wang, Q.; Meng, Q.; Xu, F.; Chen, Q.; Liu, F.; Hu, Y.; Luo, M. GhCYP710A1 Participates in Cotton Resistance to Verticillium Wilt by Regulating Stigmasterol Synthesis and Plasma Membrane Stability. Int. J. Mol. Sci. 2022, 23, 8437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158437
Huang L, Li G, Wang Q, Meng Q, Xu F, Chen Q, Liu F, Hu Y, Luo M. GhCYP710A1 Participates in Cotton Resistance to Verticillium Wilt by Regulating Stigmasterol Synthesis and Plasma Membrane Stability. International Journal of Molecular Sciences. 2022; 23(15):8437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158437
Chicago/Turabian StyleHuang, Li, Guiming Li, Qiaoling Wang, Qian Meng, Fan Xu, Qian Chen, Fang Liu, Yulin Hu, and Ming Luo. 2022. "GhCYP710A1 Participates in Cotton Resistance to Verticillium Wilt by Regulating Stigmasterol Synthesis and Plasma Membrane Stability" International Journal of Molecular Sciences 23, no. 15: 8437. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23158437