
Association of CARD14 Single-Nucleotide Polymorphisms with Psoriasis

 , ,
, ,
Abstract
:1. Introduction
2. Results
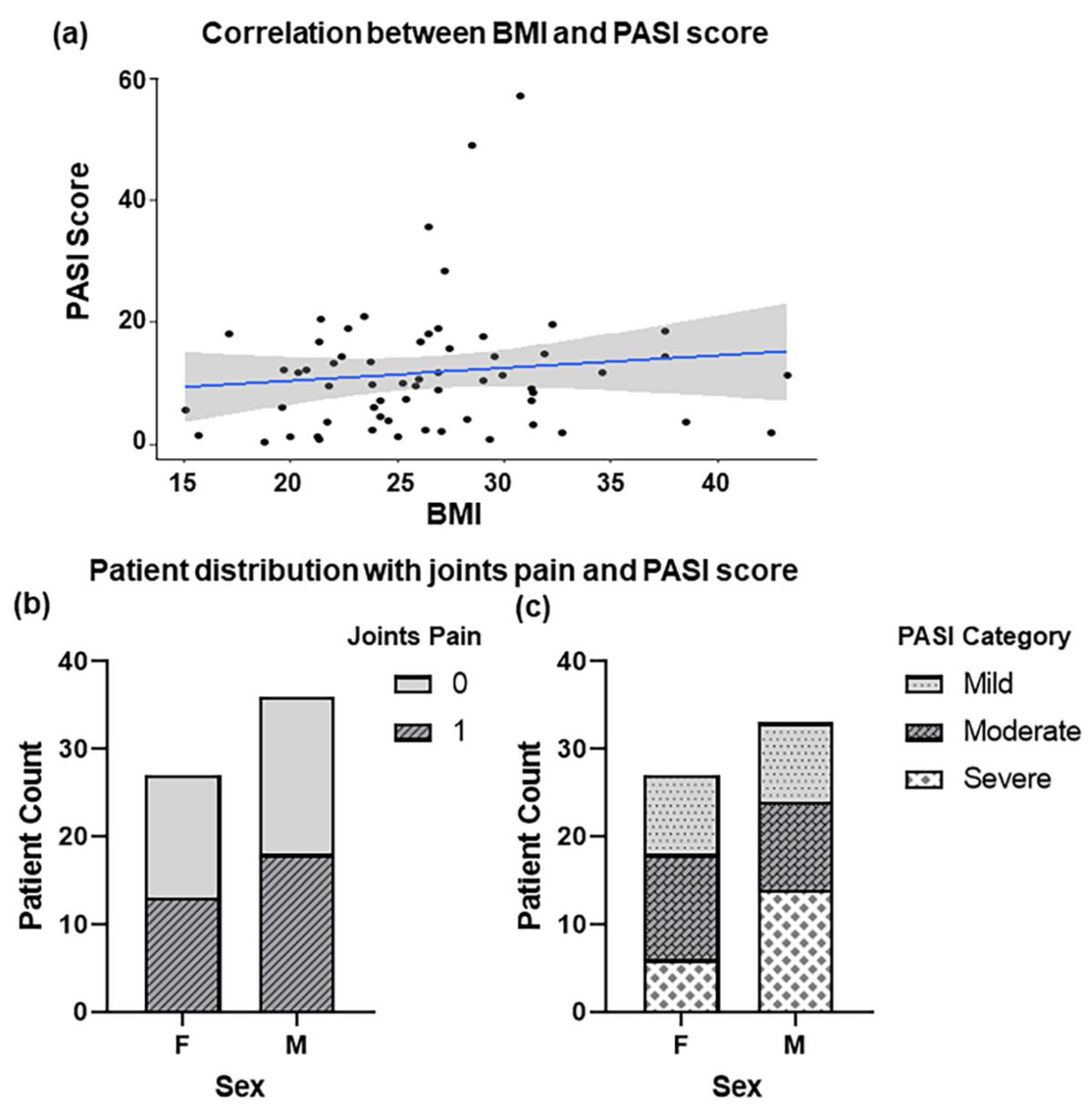
2.1. Clinical Presentation of Psoriasis Patients
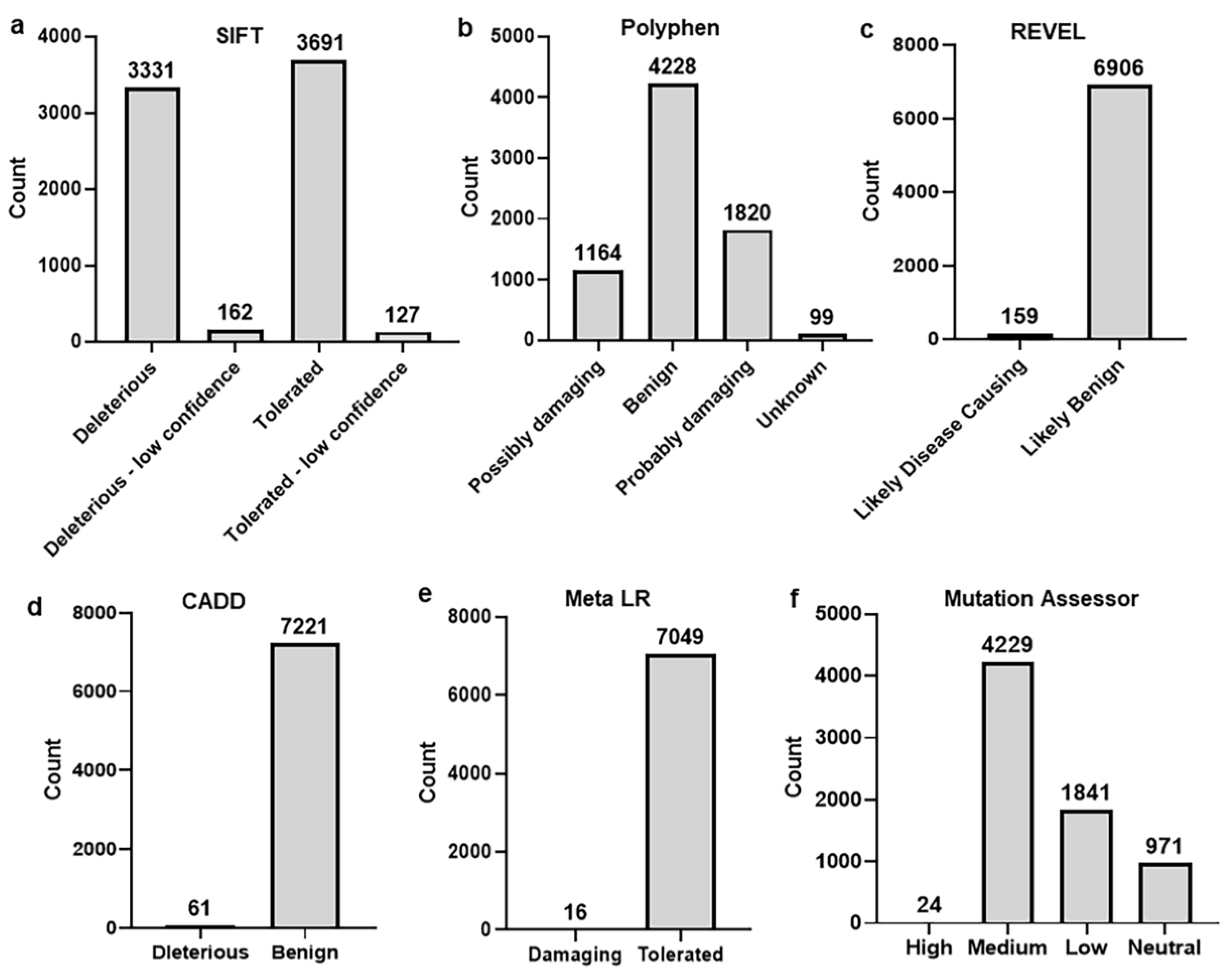
2.2. Identification and Selection of nsSNPs
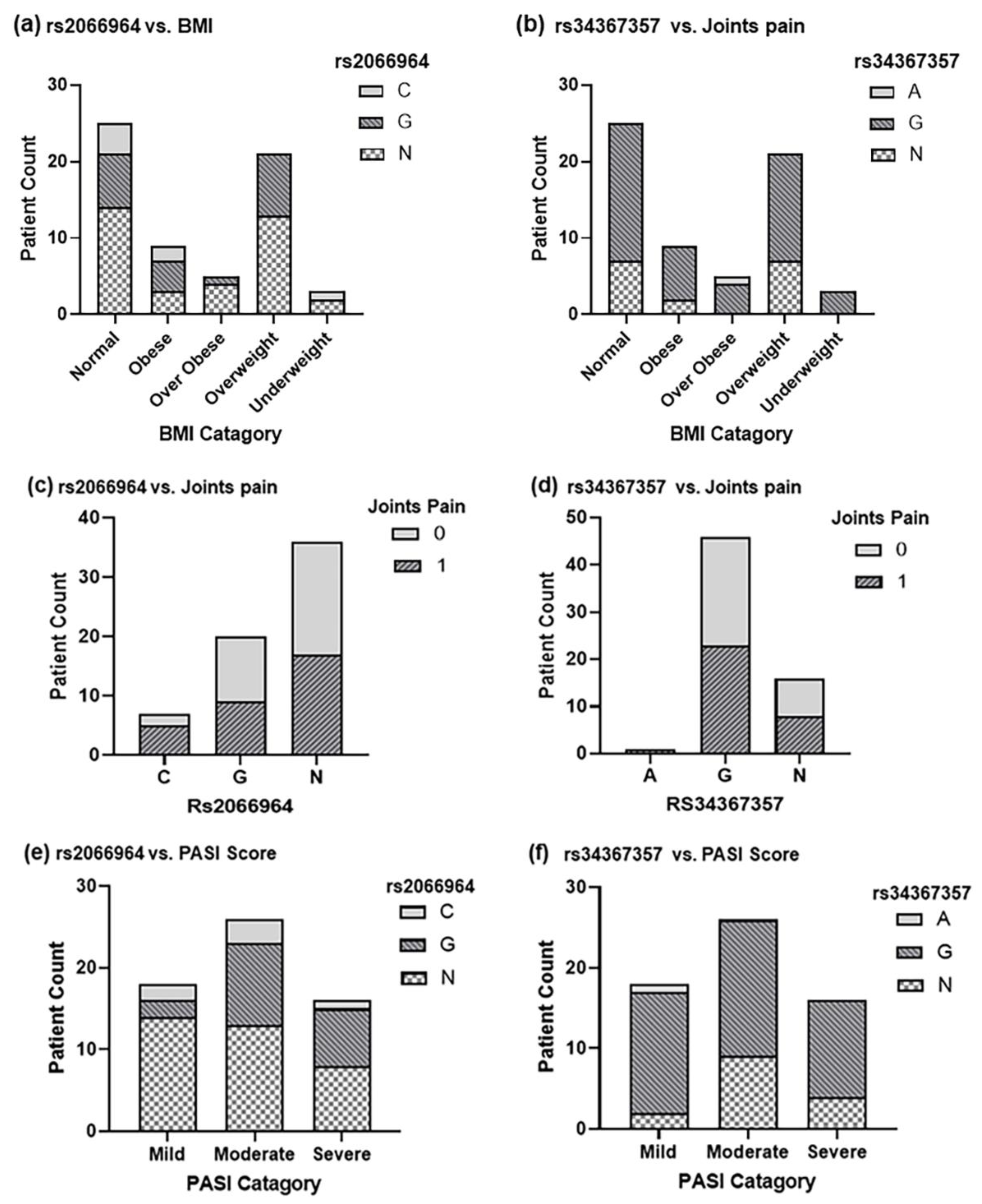
2.3. Genotyping of CARD14 in Diseased and Control Samples
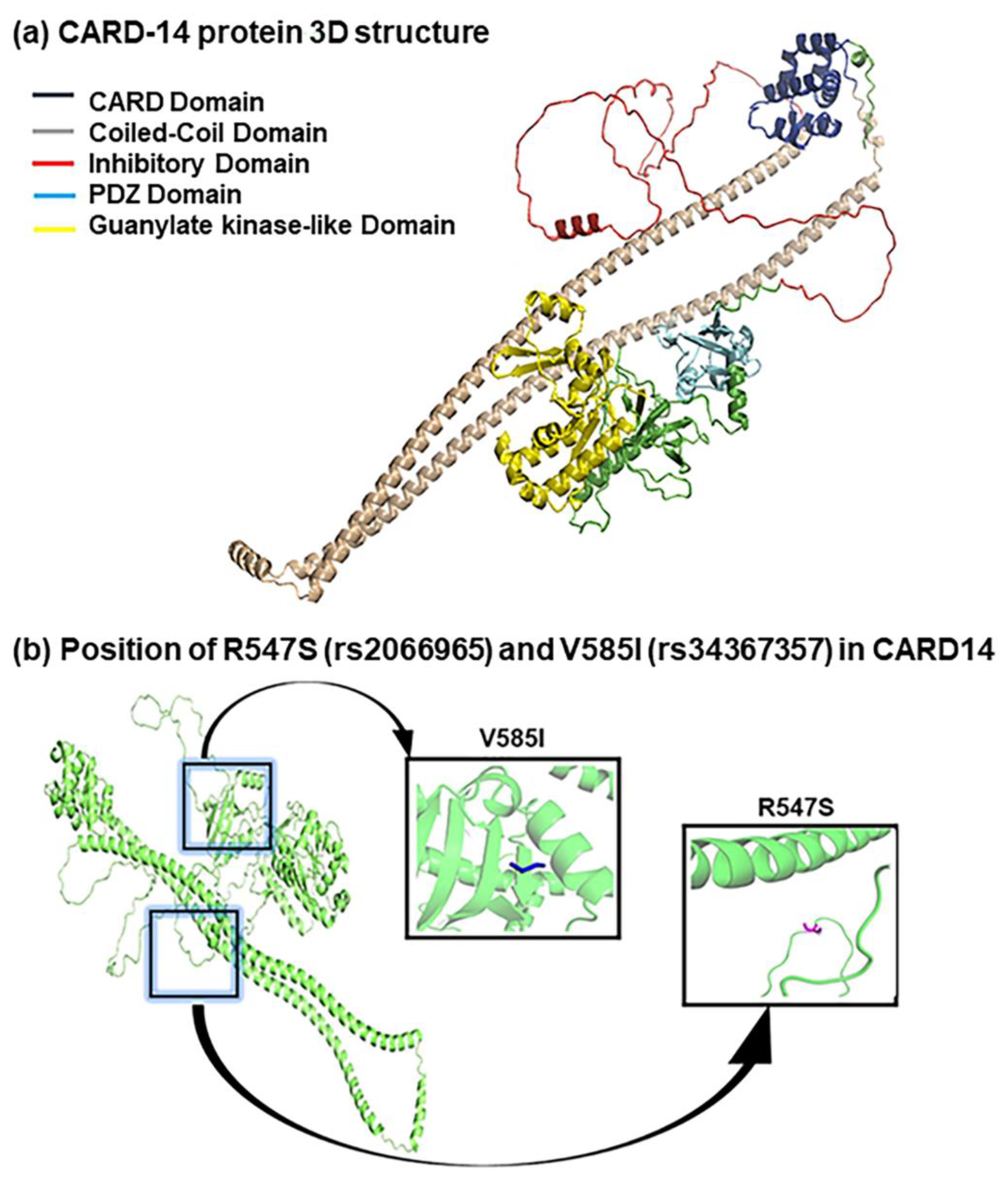
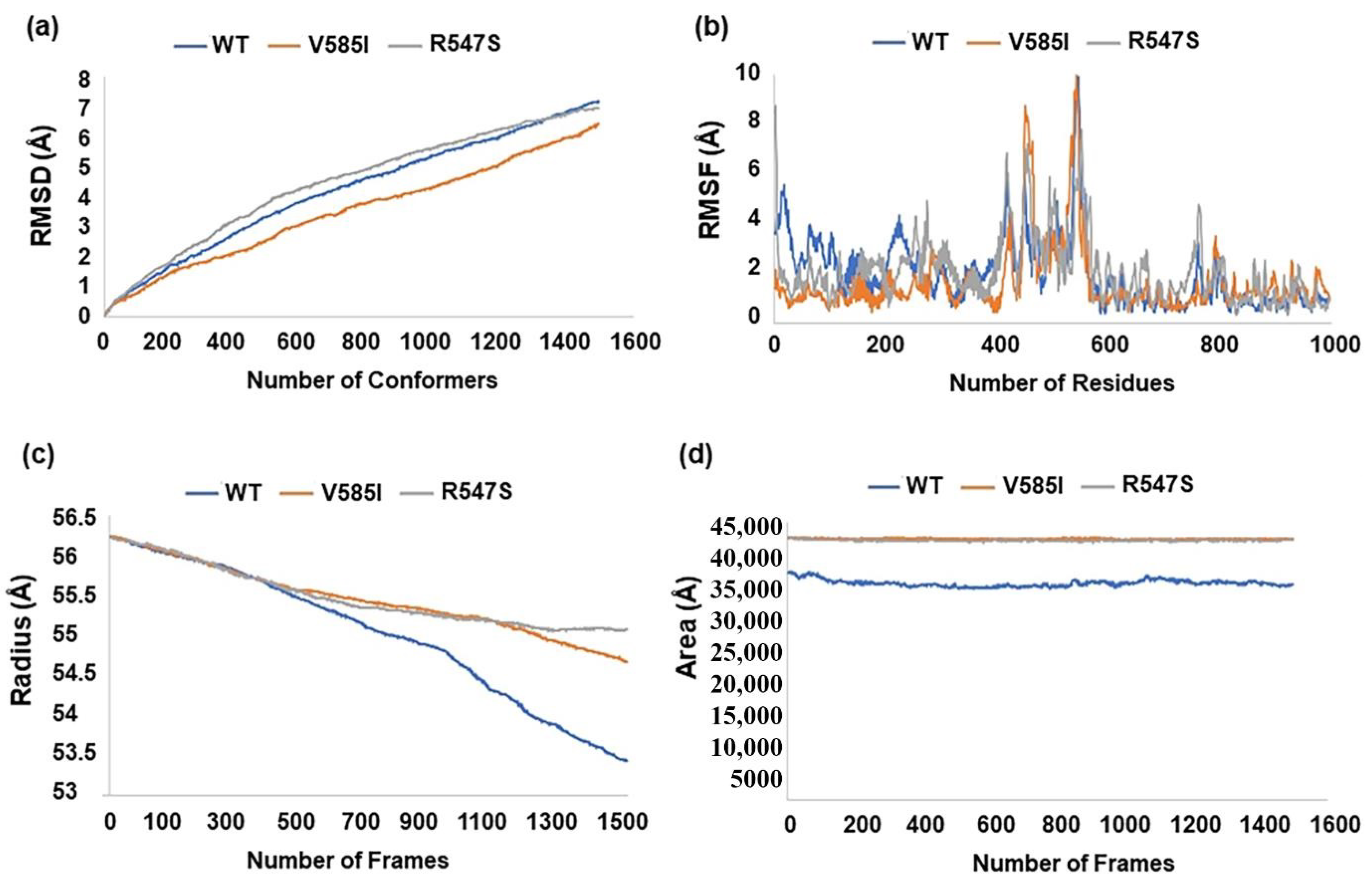
2.4. Structural Analysis of SNPs Associated with Psoriasis
3. Discussion
4. Materials and Methods
4.1. Study Subjects and Sample Collection
4.2. Selection of nsSNPs
4.3. PCR Amplification and Sanger Sequencing
4.4. Statistical Analysis
4.5. 3D Structure Prediction and Simulations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rendon, A.; Schäkel, K. Psoriasis pathogenesis and treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, S. Skin disease and socioeconomic conditions in rural Africa: Tanzania. Int. J. Dermatol. 1996, 35, 633–639. [Google Scholar] [CrossRef] [PubMed]
- Chandran, V.; Raychaudhuri, S.P. Geoepidemiology and environmental factors of psoriasis and psoriatic arthritis. J. Autoimmun. 2010, 34, J314–J321. [Google Scholar] [CrossRef] [PubMed]
- Augustin, M.; Eissing, L.; Langenbruch, A.; Enk, A.; Luger, T.; Maaßen, D.; Mrowietz, U.; Reich, K.; Reusch, M.; Strömer, K. The German National Program on psoriasis health care 2005–2015: Results and experiences. Arch. Dermatol. Res. 2016, 308, 389–400. [Google Scholar] [CrossRef]
- Capon, F. The genetic basis of psoriasis. Int. J. Mol. Sci. 2017, 18, 2526. [Google Scholar] [CrossRef]
- Consortium, I.P.G. The International Psoriasis Genetics Study: Assessing linkage to 14 candidate susceptibility loci in a cohort of 942 affected sib pairs. Am. J. Hum. Genet. 2003, 73, 430–437. [Google Scholar]
- Balendran, N.; Clough, R.L.; Arguello, J.R.; Barber, R.; Veal, C.; Jones, A.B.; Rosbotham, J.L.; Little, A.-M.; Madrigal, A.; Barker, J.N. Characterization of the major susceptibility region for psoriasis at chromosome. J. Investig. Dermatol. 1999, 113, 322–328. [Google Scholar] [CrossRef]
- Tomfohrde, J.; Silverman, A.; Barnes, R.; Fernandez-Vina, M.A.; Young, M.; Lory, D.; Morris, L.; Wuepper, K.D.; Stastny, P.; Menter, A. Gene for familial psoriasis susceptibility mapped to the distal end of human chromosome 17q. Science 1994, 264, 1141–1145. [Google Scholar] [CrossRef]
- De Cid, R.; Riveira-Munoz, E.; Zeeuwen, P.L.; Robarge, J.; Liao, W.; Dannhauser, E.N.; Giardina, E.; Stuart, P.E.; Nair, R.; Helms, C. Deletion of the late cornified envelope LCE3B and LCE3C genes as a susceptibility factor for psoriasis. Nat. Genet. 2009, 41, 211–215. [Google Scholar] [CrossRef]
- Nair, R.P.; Stuart, P.E.; Nistor, I.; Hiremagalore, R.; Chia, N.V.; Jenisch, S.; Weichenthal, M.; Abecasis, G.R.; Lim, H.W.; Christophers, E. Sequence and haplotype analysis supports HLA-C as the psoriasis susceptibility 1 gene. Am. J. Hum. Genet. 2006, 78, 827–851. [Google Scholar] [CrossRef]
- Scudiero, I.; Zotti, T.; Ferravante, A.; Vessichelli, M.; Vito, P.; Stilo, R. Alternative splicing of CARMA2/CARD14 transcripts generates protein variants with differential effect on NF-κB activation and endoplasmic reticulum stress-induced cell death. J. Cell. Physiol. 2011, 226, 3121–3131. [Google Scholar] [CrossRef] [PubMed]
- Jordan, C.T.; Cao, L.; Roberson, E.D.; Pierson, K.C.; Yang, C.-F.; Joyce, C.E.; Ryan, C.; Duan, S.; Helms, C.A.; Liu, Y. PSORS2 is due to mutations in CARD14. Am. J. Hum. Genet. 2012, 90, 784–795. [Google Scholar] [CrossRef] [PubMed]
- Fuchs-Telem, D.; Sarig, O.; van Steensel, M.A.; Isakov, O.; Israeli, S.; Nousbeck, J.; Richard, K.; Winnepenninckx, V.; Vernooij, M.; Shomron, N. Familial pityriasis rubra pilaris is caused by mutations in CARD14. Am. J. Hum. Genet. 2012, 91, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Shaiq, P.; Stuart, P.; Latif, A.; Schmotzer, C.; Kazmi, A.; Khan, M.; Azam, M.; Tejasvi, T.; Voorhees, J.; Raja, G. Genetic associations of psoriasis in a Pakistani population. Br. J. Dermatol. 2013, 169, 406–411. [Google Scholar] [CrossRef]
- Munir, S.; ber Rahman, S.; Rehman, S.; Saba, N.; Ahmad, W.; Nilsson, S.; Mazhar, K.; Naluai, Å.T. Association analysis of GWAS and candidate gene loci in a Pakistani population with psoriasis. Mol. Immunol. 2015, 64, 190–194. [Google Scholar] [CrossRef]
- Kishimoto, M.; Deshpande, G.A.; Fukuoka, K.; Kawakami, T.; Ikegaya, N.; Kawashima, S.; Komagata, Y.; Kaname, S. Clinical features of psoriatic arthritis. Best Pract. Res. Clin. Rheumatol. 2021, 35, 101670. [Google Scholar] [CrossRef]
- Gerdes, S.; Körber, A.; Biermann, M.; Karnthaler, C.; Reinhardt, M. Absolute and relative psoriasis area and severity index (PASI) treatment goals and their association with health-related quality of life. J. Dermatol. Treat. 2020, 31, 470–475. [Google Scholar] [CrossRef]
- Hassan, M.S.; Shaalan, A.; Dessouky, M.; Abdelnaiem, A.E.; ElHefnawi, M. Evaluation of computational techniques for predicting non-synonymous single nucleotide variants pathogenicity. Genomics 2019, 111, 869–882. [Google Scholar] [CrossRef]
- Adzhubei, I.; Jordan, D.M.; Sunyaev, S.R. Predicting functional effect of human missense mutations using PolyPhen-2. Curr. Protoc. Hum. Genet. 2013, 76, 7. [Google Scholar] [CrossRef]
- Ng, P.C.; Henikoff, S. SIFT: Predicting amino acid changes that affect protein function. Nucleic Acids Res. 2003, 31, 3812–3814. [Google Scholar] [CrossRef]
- Rentzsch, P.; Witten, D.; Cooper, G.M.; Shendure, J.; Kircher, M. CADD: Predicting the deleteriousness of variants throughout the human genome. Nucleic Acids Res. 2019, 47, D886–D894. [Google Scholar] [CrossRef] [PubMed]
- Ioannidis, N.M.; Rothstein, J.H.; Pejaver, V.; Middha, S.; McDonnell, S.K.; Baheti, S.; Musolf, A.; Li, Q.; Holzinger, E.; Karyadi, D. REVEL: An ensemble method for predicting the pathogenicity of rare missense variants. Am. J. Hum. Genet. 2016, 99, 877–885. [Google Scholar] [CrossRef] [PubMed]
- Alirezaie, N.; Kernohan, K.D.; Hartley, T.; Majewski, J.; Hocking, T.D. ClinPred: Prediction tool to identify disease-relevant nonsynonymous single-nucleotide variants. Am. J. Hum. Genet. 2018, 103, 474–483. [Google Scholar] [CrossRef]
- Israel, L.; Mellett, M. Clinical and genetic heterogeneity of CARD14 mutations in psoriatic skin disease. Front. Immunol. 2018, 9, 2239. [Google Scholar] [CrossRef] [PubMed]
- Chua, K.; Kee, B.; Tan, S.; Lian, L. Interleukin-6 promoter polymorphisms (-174 G/C) in Malaysian patients with systemic lupus erythematosus. Braz. J. Med. Biol. Res. 2009, 42, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Al Quraishi, M. AlphaFold at CASP13. Bioinformatics 2019, 35, 4862–4865. [Google Scholar] [CrossRef]
- Mulder, N.; Apweiler, R. Interpro and interproscan. In Comparative Genomics; Springer: Berlin/Heidelberg, Germany, 2007; pp. 59–70. [Google Scholar]
- Howes, A.; O’Sullivan, P.A.; Breyer, F.; Ghose, A.; Cao, L.; Krappmann, D.; Bowcock, A.M.; Ley, S.C. Psoriasis mutations disrupt CARD14 autoinhibition promoting BCL10-MALT1-dependent NF-κB activation. Biochem. J. 2016, 473, 1759–1768. [Google Scholar] [CrossRef]
- Zotti, T.; Polvere, I.; Voccola, S.; Vito, P. CARD14/CARMA2 signaling and its role in inflammatory skin disorders. Front. Immunol. 2018, 9, 2167. [Google Scholar] [CrossRef]
- Wang, M.; Zhang, S.; Zheng, G.; Huang, J.; Songyang, Z.; Zhao, X.; Lin, X. Gain-of-function mutation of Card14 leads to spontaneous psoriasis-like skin inflammation through enhanced keratinocyte response to IL-17A. Immunity 2018, 49, 66–79.e5. [Google Scholar] [CrossRef]
- Li, L.; You, J.; Fu, X.; Wang, Z.; Sun, Y.; Liu, H.; Zhang, F. Variants of CARD14 are predisposing factors for generalized pustular psoriasis (GPP) with psoriasis vulgaris but not for GPP alone in a Chinese population. Br. J. Dermatol. 2019, 180, 425–426. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, C.; Tao, L.; Zhou, J.; Zhu, J.; Yan, K.; Han, L.; Huang, Q.; Shi, J.; Zhang, Z. Updating and identifying three novel variants of the CARD14 gene in Chinese Han patients with psoriasis. J. Genet. 2020, 99, 1–4. [Google Scholar] [CrossRef]
- Sugiura, K.; Muto, M.; Akiyama, M. CARD14 c. 526G > C (p. Asp176His) is a significant risk factor for generalized pustular psoriasis with psoriasis vulgaris in the Japanese cohort. J. Investig. Dermatol. 2014, 134, 1755–1757. [Google Scholar] [CrossRef] [PubMed]
- González-Lara, L.; Coto-Segura, P.; Penedo, A.; Eiris, N.; Díaz, M.; Santos-Juanes, J.; Queiro, R.; Coto, E. SNP rs11652075 in the CARD14 gene as a risk factor for psoriasis (PSORS2) in a Spanish cohort. DNA Cell Biol. 2013, 32, 601–604. [Google Scholar] [CrossRef]
- Mössner, R.; Frambach, Y.; Wilsmann-Theis, D.; Löhr, S.; Jacobi, A.; Weyergraf, A.; Müller, M.; Philipp, S.; Renner, R.; Traupe, H. Palmoplantar pustular psoriasis is associated with missense variants in CARD14, but not with loss-of-function mutations in IL36RN in European patients. J. Investig. Dermatol. 2015, 135, 2538. [Google Scholar] [CrossRef] [PubMed]
- Zhu, K.; Shi, G.; Liu, H.; Zhu, C.; Fan, Y. Variants of CARD14 gene and psoriasis vulgaris in southern Chinese cohort. An. Bras. De Dermatol. 2016, 91, 45–48. [Google Scholar] [CrossRef]
- Feng, C.; Wang, T.; Li, S.J.; Fan, Y.M.; Shi, G.; Zhu, K.J. CARD 14 gene polymorphism c. C2458T (p. Arg820Trp) is associated with clinical features of psoriasis vulgaris in a Chinese cohort. J. Dermatol. 2016, 43, 294–297. [Google Scholar] [CrossRef] [PubMed]
- Ammar, M.; Jordan, C.; Cao, L.; Lim, E.; Bouchlaka Souissi, C.; Jrad, A.; Omrane, I.; Kouidhi, S.; Zaraa, I.; Anbunathan, H. CARD 14 alterations in Tunisian patients with psoriasis and further characterization in European cohorts. Br. J. Dermatol. 2016, 174, 330–337. [Google Scholar] [CrossRef]
- Van Nuffel, E.; Afonina, I.S.; Beyaert, R. Psoriasis plays a wild CARD. J. Investig. Dermatol. 2018, 138, 1903–1905. [Google Scholar] [CrossRef]
- Mellett, M.; Meier, B.; Mohanan, D.; Schairer, R.; Cheng, P.; Satoh, T.K.; Kiefer, B.; Ospelt, C.; Nobbe, S.; Thome, M. CARD14 gain-of-function mutation alone is sufficient to drive IL-23/IL-17–mediated psoriasiform skin inflammation in vivo. J. Investig. Dermatol. 2018, 138, 2010–2023. [Google Scholar] [CrossRef]
- Danis, J.; Göblös, A.; Gál, B.; Sulák, A.; Farkas, K.; Török, D.; Varga, E.; Korom, I.; Kemény, L.; Széll, M. Nuclear factor κB activation in a Type V pityriasis rubra pilaris patient harboring multiple CARD14 variants. Front. Immunol. 2018, 9, 1564. [Google Scholar] [CrossRef]
- Qin, P.; Zhang, Q.; Chen, M.; Fu, X.A.; Wang, C.; Wang, Z.; Yu, G.; Yu, Y.; Li, X.; Sun, Y. Variant analysis of CARD14 in a Chinese Han population with psoriasis vulgaris and generalized pustular psoriasis. J. Investig. Dermatol. 2014, 134, 2994. [Google Scholar] [CrossRef] [PubMed]
- Gál, B.; Göblös, A.; Danis, J.; Farkas, K.; Sulák, A.; Varga, E.; Nagy, N.; Széll, M.; Kemény, L.; Bata-Csörgő, Z. The management and genetic background of pityriasis rubra pilaris: A single-centre experience. J. Eur. Acad. Dermatol. Venereol. 2019, 33, 944–949. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Fang, Y.; Cao, G.; Zhang, S.; Tian, M.; Shen, Q.; Xu, H.; Liu, C.; Rao, J. Genetic and pathological findings in a boy with psoriasis and C3 glomerulonephritis: A case report and literature review. Mol. Genet. Genom. Med. 2020, 8, e1430. [Google Scholar] [CrossRef] [PubMed]
- Fleming, P.; Kraft, J.; Gulliver, W.P.; Lynde, C. The Relationship of Obesity With the Severity of Psoriasis: A Systematic Review. J. Cutan. Med. Surg. 2015, 19, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Sobhan, M.; Farshchian, M. Associations between body mass index and severity of psoriasis. Clin. Cosmet. Investig. Derm. 2017, 10, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Bouchier-Hayes, L.; Martin, S.J. CARD games in apoptosis and immunity. EMBO Rep. 2002, 3, 616–621. [Google Scholar] [CrossRef] [PubMed]
- Ponting, C.P.; Phillips, C.; Davies, K.E.; Blake, D.J. PDZ domains: Targeting signalling molecules to sub-membranous sites. Bioessays 1997, 19, 469–479. [Google Scholar] [CrossRef]
- Stehle, T.; Schulz, G.E. Refined structure of the complex between guanylate kinase and its substrate GMP at 20 Å resolution. J. Mol. Biol. 1992, 224, 1127–1141. [Google Scholar] [CrossRef]
- Yates, C.M.; Sternberg, M.J. The effects of non-synonymous single nucleotide polymorphisms (nsSNPs) on protein–protein interactions. J. Mol. Biol. 2013, 425, 3949–3963. [Google Scholar] [CrossRef]
- Untergasser, A.; Cutcutache, I.; Koressaar, T.; Ye, J.; Faircloth, B.C.; Remm, M.; Rozen, S.G. Primer3—New capabilities and interfaces. Nucleic Acids Res. 2012, 40, e115. [Google Scholar] [CrossRef]
- Lewis, C.M. Genetic association studies: Design, analysis and interpretation. Brief. Bioinform. 2002, 3, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Cramer, P. AlphaFold2 and the future of structural biology. Nat. Struct. Mol. Biol. 2021, 28, 704–705. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | nsSNPs ID | Alleles | Amino Acid |
|---|---|---|---|
| 1 | rs146214639 | T/G | L150R |
| 2 | rs1598639617 | T/C | L124P |
| 3 | rs1598639659 | G/C | C127S |
| 4 | rs1598639974 | A/C | Q157P |
| 5 | rs281875215 | G/A | G117S |
| 6 | rs1567872320 | G/A | E138K |
| 7 | rs281875212 | G/A | E142K |
| 8 | rs281875213 | A/G | E142G |
| 9 | rs387907240 | T/C | L156P |
| 10 | rs281875216 | C/A | H171N |
| 11 | rs2066964 | G/C | R547S |
| 12 | rs34367357 | G/A | V585I |
| 13 | rs200102454 | C/T | T591M |
| 14 | rs281875220 | T/A | I593N |
| 15 | rs201285077 | C/T | S602L |
| 16 | rs117918077 | C/T | R682W |
| 17 | rs1567903243 | C/A | S802R |
| 18 | rs11652075 | C/T | R820W |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suleman, S.; Chhabra, G.; Raza, R.; Hamid, A.; Qureshi, J.A.; Ahmad, N. Association of CARD14 Single-Nucleotide Polymorphisms with Psoriasis. Int. J. Mol. Sci. 2022, 23, 9336. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169336
Suleman S, Chhabra G, Raza R, Hamid A, Qureshi JA, Ahmad N. Association of CARD14 Single-Nucleotide Polymorphisms with Psoriasis. International Journal of Molecular Sciences. 2022; 23(16):9336. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169336
Chicago/Turabian StyleSuleman, Saima, Gagan Chhabra, Rubab Raza, Arslan Hamid, Javed Anver Qureshi, and Nihal Ahmad. 2022. "Association of CARD14 Single-Nucleotide Polymorphisms with Psoriasis" International Journal of Molecular Sciences 23, no. 16: 9336. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169336