
Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons


Abstract
:1. Introduction
2. Results
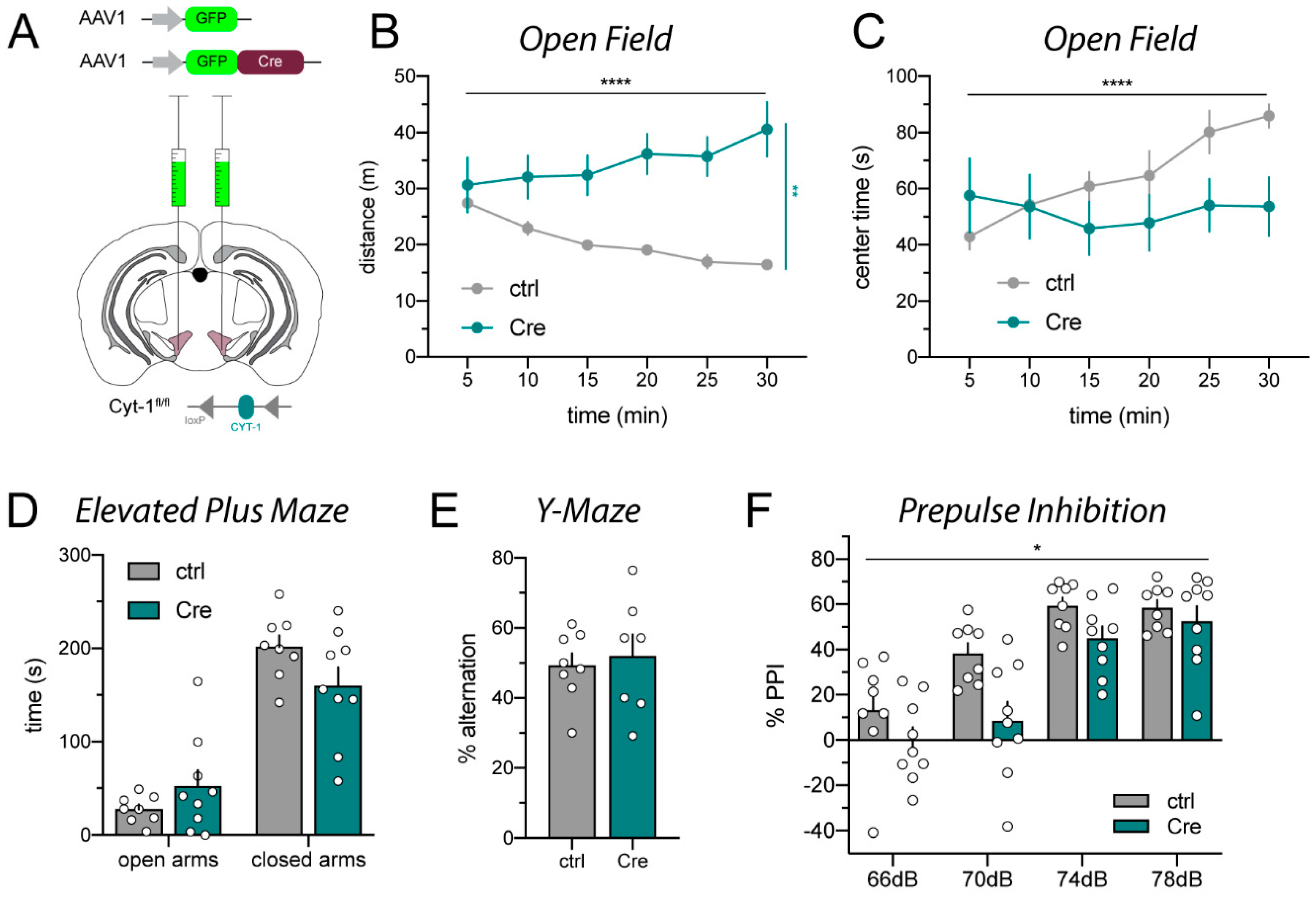
2.1. AAV_Cre Microinjections into the VTA Provoke Hyperactivity
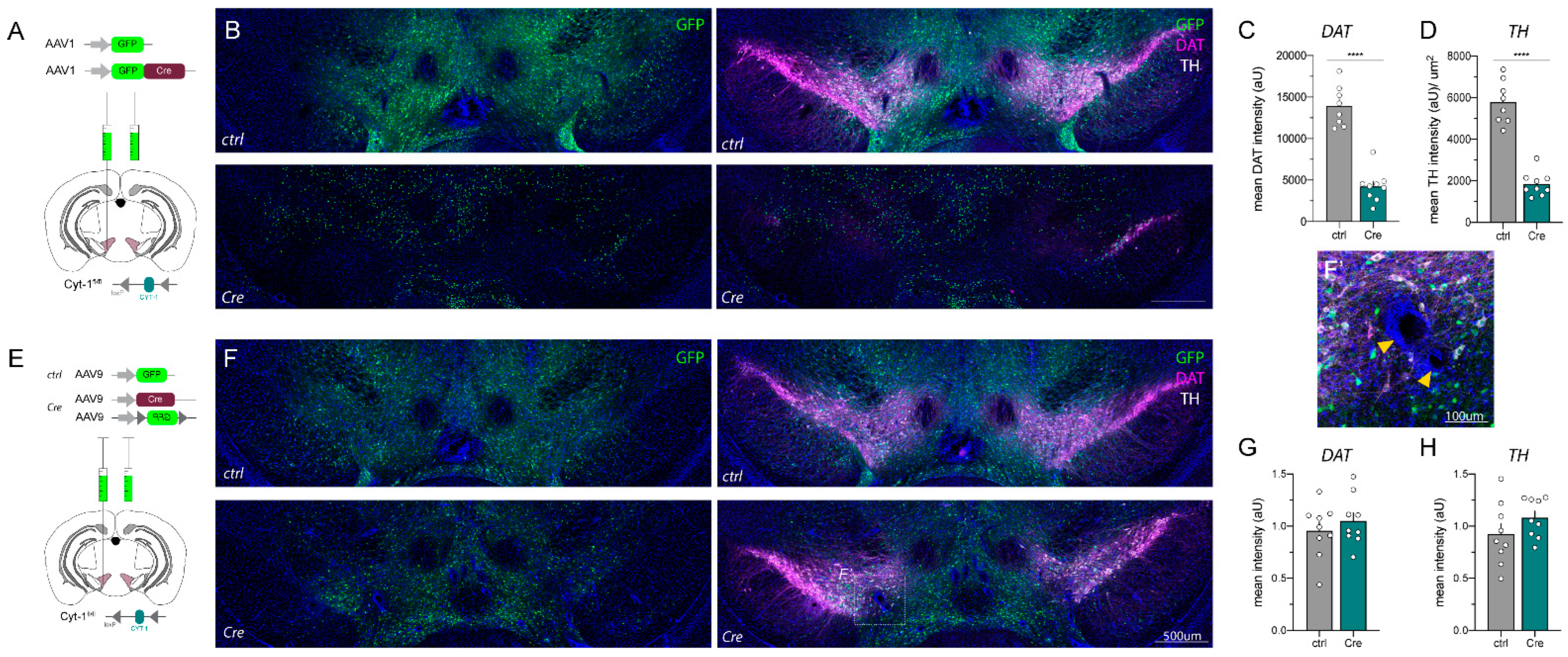
2.2. AAV_Cre Injections in the Mesencephalon Can Cause Loss of Key Protein Expression and Lesions
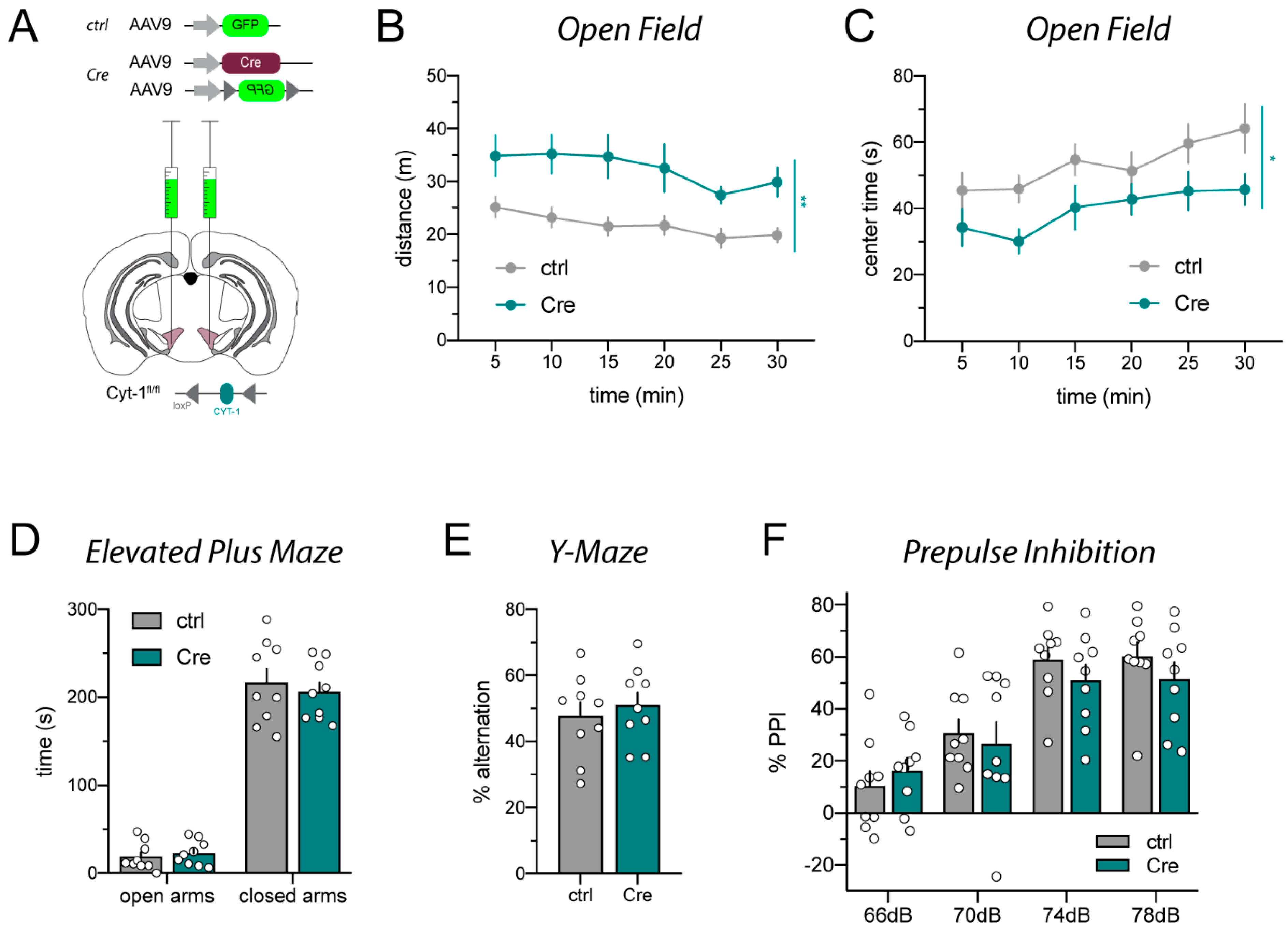
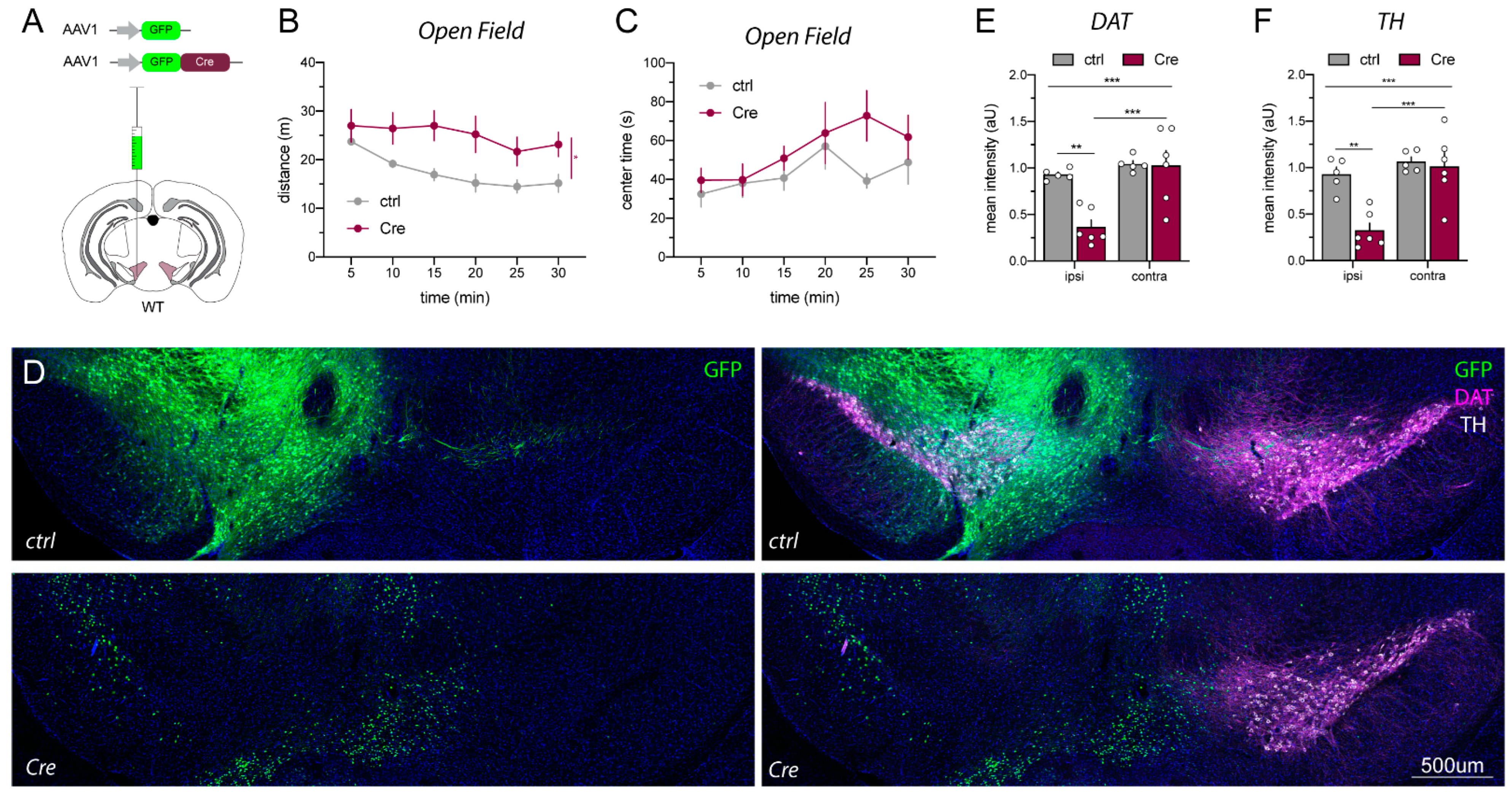
2.3. AAV1_Cre Injections in Wild-Type Mice Confirm That Loss of Protein Expression and Hyperactivity Are Unspecific
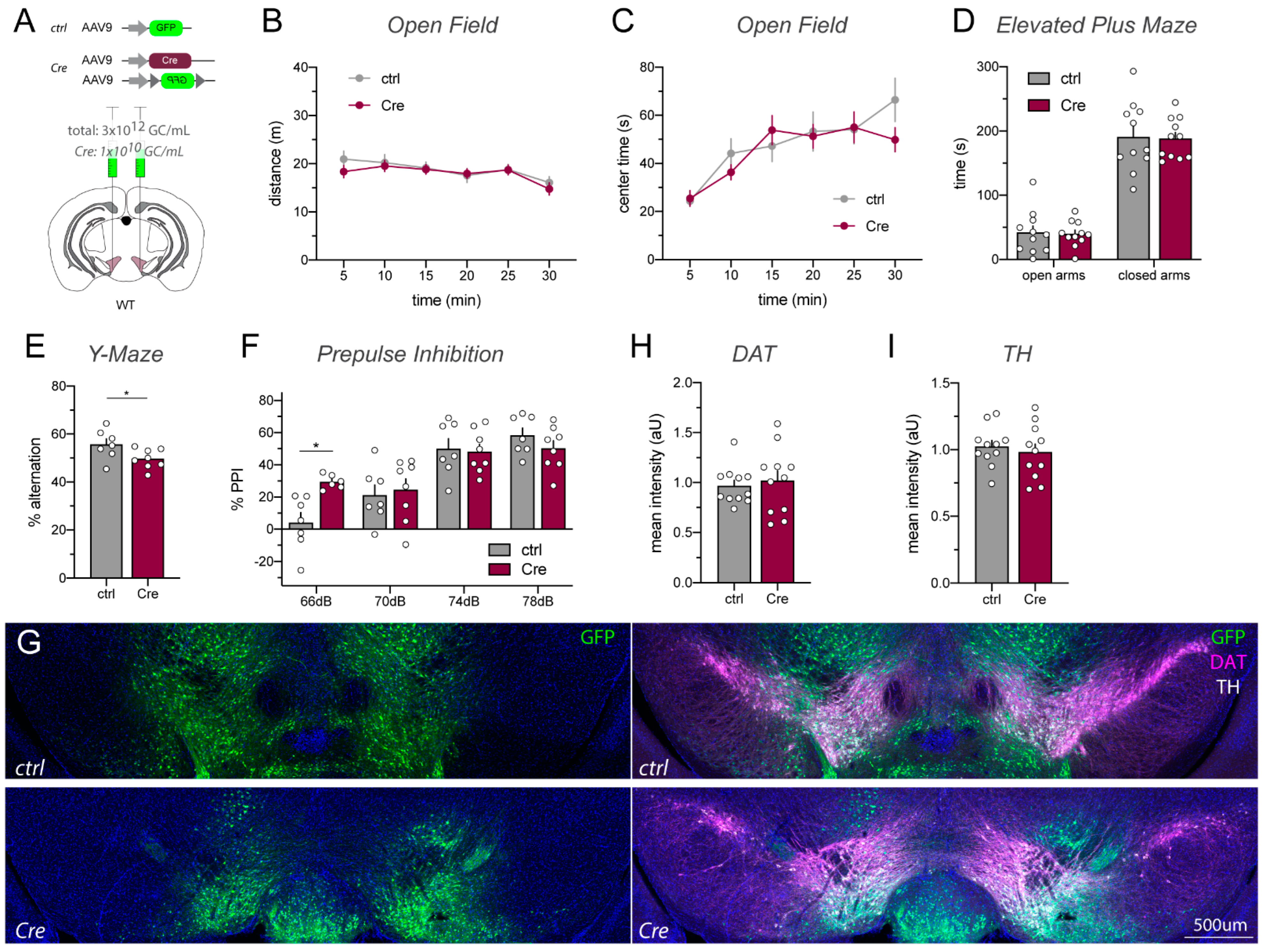
2.4. Lowering AAV_Cre Titer Prevents Toxicity While Retaining Cre/loxP Recombination Activity
3. Discussion
3.1. Toxicity Associated with High AAV_Cre Recombinase Expression Levels
3.2. Different AAV_Cre Viral Vectors Can Elicit Overlapping and Distinct Phenotypes
3.3. High Cre Recombinase Levels Can Cause Damage That Is Preventable by Reducing Expression
4. Materials and Methods
4.1. Animals
4.2. Stereotaxic Injections and Post-hoc Confirmation of Injection Sites
4.3. Battery of Behavior Tests
4.3.1. Open Field
4.3.2. Elevated Plus Maze
4.3.3. Y-Maze
4.3.4. Prepulse Inhibition (PPI)
4.4. Immunofluorescence Histochemistry (IHC)
4.5. Experimental Design and Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| RRID | Research Resource Identifier, see scicrunch.org (accessed 25 August 2020) |
| AAV | adeno-associated virus |
| AAV_GFP | GFP expressing AAV |
| AAV_Cre | Cre expressing AAV |
| DIO | double-floxed inverse orientation |
| NRG | neuregulin |
| VTA | ventral tegmental area |
| SNC | substantia nigra compacta |
| TH | tyrosine hydroxylase |
| DAT | dopamine transporter |
| ErbB4 Cyt-1fl/fl | ErbB4 Cyt-1-floxed mice |
| PPI | prepulse inhibition |
| IHC | immunohistochemistry |
References
- Buonanno, A.; Fischbach, G.D. Neuregulin and ErbB receptor signaling pathways in the nervous system. Curr. Opin. Neurobiol. 2001, 11, 287–296. [Google Scholar] [CrossRef]
- Mei, L.; Nave, K.A. Neuregulin-ERBB signaling in the nervous system and neuropsychiatric diseases. Neuron 2014, 83, 27–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stefansson, H.; Sigurdsson, E.; Steinthorsdottir, V.; Bjornsdottir, S.; Sigmundsson, T.; Ghosh, S.; Brynjolfsson, J.; Gunnarsdottir, S.; Ivarsson, O.; Chou, T.T.; et al. Neuregulin 1 and susceptibility to schizophrenia. Am. J. Hum. Genet. 2002, 71, 877–892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silberberg, G.; Darvasi, A.; Pinkas-Kramarski, R.; Navon, R. The involvement of ErbB4 with schizophrenia: Association and expression studies. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2006, 141, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, T.A.; Light, G.A.; Swerdlow, N.R.; Radant, A.D.; Braff, D.L. Association analysis of 94 candidate genes and schizophrenia-related endophenotypes. PLoS ONE 2012, 7, e29630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostaid, M.S.; Lloyd, D.; Liberg, B.; Sundram, S.; Pereira, A.; Pantelis, C.; Karl, T.; Weickert, C.S.; Everall, I.P.; Bousman, C.A. Neuregulin-1 and schizophrenia in the genome-wide association study era. Neurosci. Biobehav. Rev. 2016, 68, 387–409. [Google Scholar] [CrossRef]
- Steiner, H.; Blum, M.; Kitai, S.T.; Fedi, P. Differential expression of ErbB3 and ErbB4 neuregulin receptors in dopamine neurons and forebrain areas of the adult rat. Exp. Neurol. 1999, 159, 494–503. [Google Scholar] [CrossRef]
- Thuret, S.; Alavian, K.N.; Gassmann, M.; Lloyd, C.K.; Smits, S.M.; Smidt, M.P.; Klein, R.; Dyck, R.H.; Simon, H.H. The neuregulin receptor, ErbB4, is not required for normal development and adult maintenance of the substantia nigra pars compacta. J. Neurochem. 2004, 91, 1302–1311. [Google Scholar] [CrossRef]
- Skirzewski, M.; Karavanova, I.; Shamir, A.; Erben, L.; Garcia-Olivares, J.; Shin, J.H.; Vullhorst, D.; Alvarez, V.A.; Amara, S.G.; Buonanno, A. ErbB4 signaling in dopaminergic axonal projections increases extracellular dopamine levels and regulates spatial/working memory behaviors. Mol. Psychiatry 2018, 23, 2227–2237. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.; Shamir, A.; Skirzewski, M.; Leiva-Salcedo, E.; Kwon, O.B.; Karavanova, I.; Paredes, D.; Malkesman, O.; Bailey, K.R.; Vullhorst, D.; et al. Neuregulin-2 ablation results in dopamine dysregulation and severe behavioral phenotypes relevant to psychiatric disorders. Mol. Psychiatry 2018, 23, 1233–1243. [Google Scholar] [CrossRef] [Green Version]
- Kwon, O.B.; Paredes, D.; Gonzalez, C.M.; Neddens, J.; Hernandez, L.; Vullhorst, D.; Buonanno, A. Neuregulin-1 regulates LTP at CA1 hippocampal synapses through activation of dopamine D4 receptors. Proc. Natl. Acad. Sci. USA 2008, 105, 15587–15592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno, M.; Sotoyama, H.; Namba, H.; Shibuya, M.; Eda, T.; Wang, R.; Okubo, T.; Nagata, K.; Iwakura, Y.; Nawa, H. ErbB inhibitors ameliorate behavioral impairments of an animal model for schizophrenia: Implication of their dopamine-modulatory actions. Transl. Psychiatry 2013, 3, e252. [Google Scholar] [CrossRef] [PubMed]
- Namba, H.; Okubo, T.; Nawa, H. Perinatal Exposure to Neuregulin-1 Results in Disinhibition of Adult Midbrain Dopaminergic Neurons: Implication in Schizophrenia Modeling. Sci. Rep. 2016, 6, 22606. [Google Scholar] [CrossRef] [Green Version]
- Skirzewski, M.; Cronin, M.E.; Murphy, R.; Fobbs, W.; Kravitz, A.V.; Buonanno, A. ErbB4 Null Mice Display Altered Mesocorticolimbic and Nigrostriatal Dopamine Levels as well as Deficits in Cognitive and Motivational Behaviors. eNeuro 2020, 7. [Google Scholar] [CrossRef] [PubMed]
- Elenius, K.; Choi, C.J.; Paul, S.; Santiestevan, E.; Nishi, E.; Klagsbrun, M. Characterization of a naturally occurring ErbB4 isoform that does not bind or activate phosphatidyl inositol 3-kinase. Oncogene 1999, 18, 2607–2615. [Google Scholar] [CrossRef] [Green Version]
- Law, A.J.; Wang, Y.; Sei, Y.; O’Donnell, P.; Piantadosi, P.; Papaleo, F.; Straub, R.E.; Huang, W.; Thomas, C.J.; Vakkalanka, R.; et al. Neuregulin 1-ErbB4-PI3K signaling in schizophrenia and phosphoinositide 3-kinase-p110delta inhibition as a potential therapeutic strategy. Proc. Natl. Acad. Sci. USA 2012, 109, 12165–12170. [Google Scholar] [CrossRef] [Green Version]
- Law, A.J.; Kleinman, J.E.; Weinberger, D.R.; Weickert, C.S. Disease-associated intronic variants in the ErbB4 gene are related to altered ErbB4 splice-variant expression in the brain in schizophrenia. Hum. Mol. Genet. 2007, 16, 129–141. [Google Scholar] [CrossRef] [Green Version]
- Joshi, D.; Fullerton, J.M.; Weickert, C.S. Elevated ErbB4 mRNA is related to interneuron deficit in prefrontal cortex in schizophrenia. J. Psychiatr. Res. 2014, 53, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Chung, D.W.; Volk, D.W.; Arion, D.; Zhang, Y.; Sampson, A.R.; Lewis, D.A. Dysregulated ErbB4 Splicing in Schizophrenia: Selective Effects on Parvalbumin Expression. Am. J. Psychiatry 2016, 173, 60–68. [Google Scholar] [CrossRef] [Green Version]
- Erben, L.; Welday, J.P.; Cronin, M.E.; Murphy, R.; Skirzewski, M.; Vullhorst, D.; Carroll, S.L.; Buonanno, A. Developmental, neurochemical, and behavioral analyses of ErbB4 Cyt-1 knockout mice. J. Neurochem. 2022, 161, 435–452. [Google Scholar] [CrossRef]
- Sauer, B. Functional expression of the cre-lox site-specific recombination system in the yeast Saccharomyces cerevisiae. Mol. Cell. Biol. 1987, 7, 2087–2096. [Google Scholar] [CrossRef] [PubMed]
- Gaveriaux-Ruff, C.; Kieffer, B.L. Conditional gene targeting in the mouse nervous system: Insights into brain function and diseases. Pharmacol. Ther. 2007, 113, 619–634. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, M.; Im, S.K.; Fang, S. Mouse Cre-LoxP system: General principles to determine tissue-specific roles of target genes. Lab. Anim. Res. 2018, 34, 147–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murlidharan, G.; Samulski, R.J.; Asokan, A. Biology of adeno-associated viral vectors in the central nervous system. Front. Mol. Neurosci. 2014, 7, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedbrook, C.N.; Deverman, B.E.; Gradinaru, V. Viral Strategies for Targeting the Central and Peripheral Nervous Systems. Annu. Rev. Neurosci. 2018, 41, 323–348. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, A.; Brandon, E.P.; Kootstra, N.; Gage, F.H.; Verma, I.M. Delivery of the Cre recombinase by a self-deleting lentiviral vector: Efficient gene targeting in vivo. Proc. Natl. Acad. Sci. USA 2001, 98, 11450–11455. [Google Scholar] [CrossRef] [Green Version]
- Kaspar, B.K.; Vissel, B.; Bengoechea, T.; Crone, S.; Randolph-Moore, L.; Muller, R.; Brandon, E.P.; Schaffer, D.; Verma, I.M.; Lee, K.F.; et al. Adeno-associated virus effectively mediates conditional gene modification in the brain. Proc. Natl. Acad. Sci. USA 2002, 99, 2320–2325. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, B.Y.; Chakravarthy, S.; Eggers, R.; Hermens, W.T.; Zhang, J.Y.; Niclou, S.P.; Levelt, C.; Sablitzky, F.; Anderson, P.N.; Lieberman, A.R.; et al. Efficient delivery of Cre-recombinase to neurons in vivo and stable transduction of neurons using adeno-associated and lentiviral vectors. BMC Neurosci. 2004, 5, 4. [Google Scholar] [CrossRef] [Green Version]
- Rezai Amin, S.; Gruszczynski, C.; Guiard, B.P.; Callebert, J.; Launay, J.M.; Louis, F.; Betancur, C.; Vialou, V.; Gautron, S. Viral vector-mediated Cre recombinase expression in substantia nigra induces lesions of the nigrostriatal pathway associated with perturbations of dopamine-related behaviors and hallmarks of programmed cell death. J. Neurochem. 2019, 150, 330–340. [Google Scholar] [CrossRef]
- Wang, H.; Liu, F.; Chen, W.; Sun, X.; Cui, W.; Dong, Z.; Zhao, K.; Zhang, H.; Li, H.; Xing, G.; et al. Genetic recovery of ErbB4 in adulthood partially restores brain functions in null mice. Proc. Natl. Acad. Sci. USA 2018, 115, 13105–13110. [Google Scholar] [CrossRef] [Green Version]
- Shamir, A.; Kwon, O.B.; Karavanova, I.; Vullhorst, D.; Leiva-Salcedo, E.; Janssen, M.J.; Buonanno, A. The importance of the NRG-1/ErbB4 pathway for synaptic plasticity and behaviors associated with psychiatric disorders. J. Neurosci. 2012, 32, 2988–2997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cearley, C.N.; Wolfe, J.H. A single injection of an adeno-associated virus vector into nuclei with divergent connections results in widespread vector distribution in the brain and global correction of a neurogenetic disease. J. Neurosci. 2007, 27, 9928–9940. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Jiang, C.; Liu, P.; Wang, F.; Ma, L. Mesolimbic leptin signaling negatively regulates cocaine-conditioned reward. Transl. Psychiatry 2016, 6, e972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forni, P.E.; Scuoppo, C.; Imayoshi, I.; Taulli, R.; Dastru, W.; Sala, V.; Betz, U.A.; Muzzi, P.; Martinuzzi, D.; Vercelli, A.E.; et al. High levels of Cre expression in neuronal progenitors cause defects in brain development leading to microencephaly and hydrocephaly. J. Neurosci. 2006, 26, 9593–9602. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higashi, A.Y.; Ikawa, T.; Muramatsu, M.; Economides, A.N.; Niwa, A.; Okuda, T.; Murphy, A.J.; Rojas, J.; Heike, T.; Nakahata, T.; et al. Direct hematological toxicity and illegitimate chromosomal recombination caused by the systemic activation of CreERT2. J. Immunol. 2009, 182, 5633–5640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thyagarajan, B.; Guimaraes, M.J.; Groth, A.C.; Calos, M.P. Mammalian genomes contain active recombinase recognition sites. Gene 2000, 244, 47–54. [Google Scholar] [CrossRef]
- Huh, W.J.; Mysorekar, I.U.; Mills, J.C. Inducible activation of Cre recombinase in adult mice causes gastric epithelial atrophy, metaplasia, and regenerative changes in the absence of “floxed” alleles. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, G368–G380. [Google Scholar] [CrossRef] [Green Version]
- Janbandhu, V.C.; Moik, D.; Fassler, R. Cre recombinase induces DNA damage and tetraploidy in the absence of loxP sites. Cell Cycle 2014, 13, 462–470. [Google Scholar] [CrossRef] [Green Version]
- Loonstra, A.; Vooijs, M.; Beverloo, H.B.; Allak, B.A.; van Drunen, E.; Kanaar, R.; Berns, A.; Jonkers, J. Growth inhibition and DNA damage induced by Cre recombinase in mammalian cells. Proc. Natl. Acad. Sci. USA 2001, 98, 9209–9214. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Nguyen, M.T.; Nakamura, E.; Yang, J.; Mackem, S. Cre-mediated recombination can induce apoptosis in vivo by activating the p53 DNA damage-induced pathway. Genesis 2012, 50, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Surova, O.; Zhivotovsky, B. Various modes of cell death induced by DNA damage. Oncogene 2013, 32, 3789–3797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmore, S. Apoptosis: A review of programmed cell death. Toxicol. Pathol. 2007, 35, 495–516. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jenkins, N.A.; Copeland, N.G. A highly efficient recombineering-based method for generating conditional knockout mutations. Genome Res. 2003, 13, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Heredia-Lopez, F.J.; Alvarez-Cervera, F.J.; Colli-Alfaro, J.G.; Bata-Garcia, J.L.; Arankowsky-Sandoval, G.; Gongora-Alfaro, J.L. An automated Y-maze based on a reduced instruction set computer (RISC) microcontroller for the assessment of continuous spontaneous alternation in rats. Behav. Res. Methods 2016, 48, 1631–1643. [Google Scholar] [CrossRef] [Green Version]
- Vullhorst, D.; Ahmad, T.; Karavanova, I.; Keating, C.; Buonanno, A. Structural Similarities between Neuregulin 1–3 Isoforms Determine Their Subcellular Distribution and Signaling Mode in Central Neurons. J. Neurosci. 2017, 37, 5232–5249. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Catalog # (Addgene) | Plasmid RRID | Concentration (GC/mL) | Figure |
|---|---|---|---|---|
| AAV1.hSyn.HI.eGFP-Cre.WRPE.SV40 | 105540 | Addgene_105540 | 2.6 × 1013 | Figure 1, Figure 3 and Figure 4 |
| AAV1.hSyn.eGFP.WRPE.bGH | 105539 | Addgene_105539 | 2.7 × 1013 | Figure 1, Figure 3 and Figure 4 |
| AAV9.hSyn.Cre.hGH mixed with AAV9.synP.DIO.EGFP.WRPE.hGH | 105555 100043 | Addgene_105555 Addgene_100043 | 1.5 × 1013 1.5 × 1013 | Figure 2 and Figure 3 |
| AAV9.hSyn.eGFP.WRPE.bGH | 105539 | Addgene_105539 | 3.0 × 1013 | Figure 2 and Figure 3 |
| AAV9.hSyn.Cre.hGH mixed with AAV9.synP.DIO.EGFP.WRPE.hGH | 105555 100043 | Addgene_105555 Addgene_100043 | 1.0 × 1010 2.99 × 1012 | Figure 5 |
| AAV9.hSyn.eGFP.WRPE.bGH | 105539 | Addgene_105539 | 3.0 × 1012 | Figure 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erben, L.; Welday, J.P.; Murphy, R.; Buonanno, A. Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons. Int. J. Mol. Sci. 2022, 23, 9462. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169462
Erben L, Welday JP, Murphy R, Buonanno A. Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons. International Journal of Molecular Sciences. 2022; 23(16):9462. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169462
Chicago/Turabian StyleErben, Larissa, Jacqueline P. Welday, Ricardo Murphy, and Andres Buonanno. 2022. "Toxic and Phenotypic Effects of AAV_Cre Used to Transduce Mesencephalic Dopaminergic Neurons" International Journal of Molecular Sciences 23, no. 16: 9462. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23169462