
Paclobutrazol Ameliorates Low-Light-Induced Damage by Improving Photosynthesis, Antioxidant Defense System, and Regulating Hormone Levels in Tall Fescue



Abstract
:1. Introduction
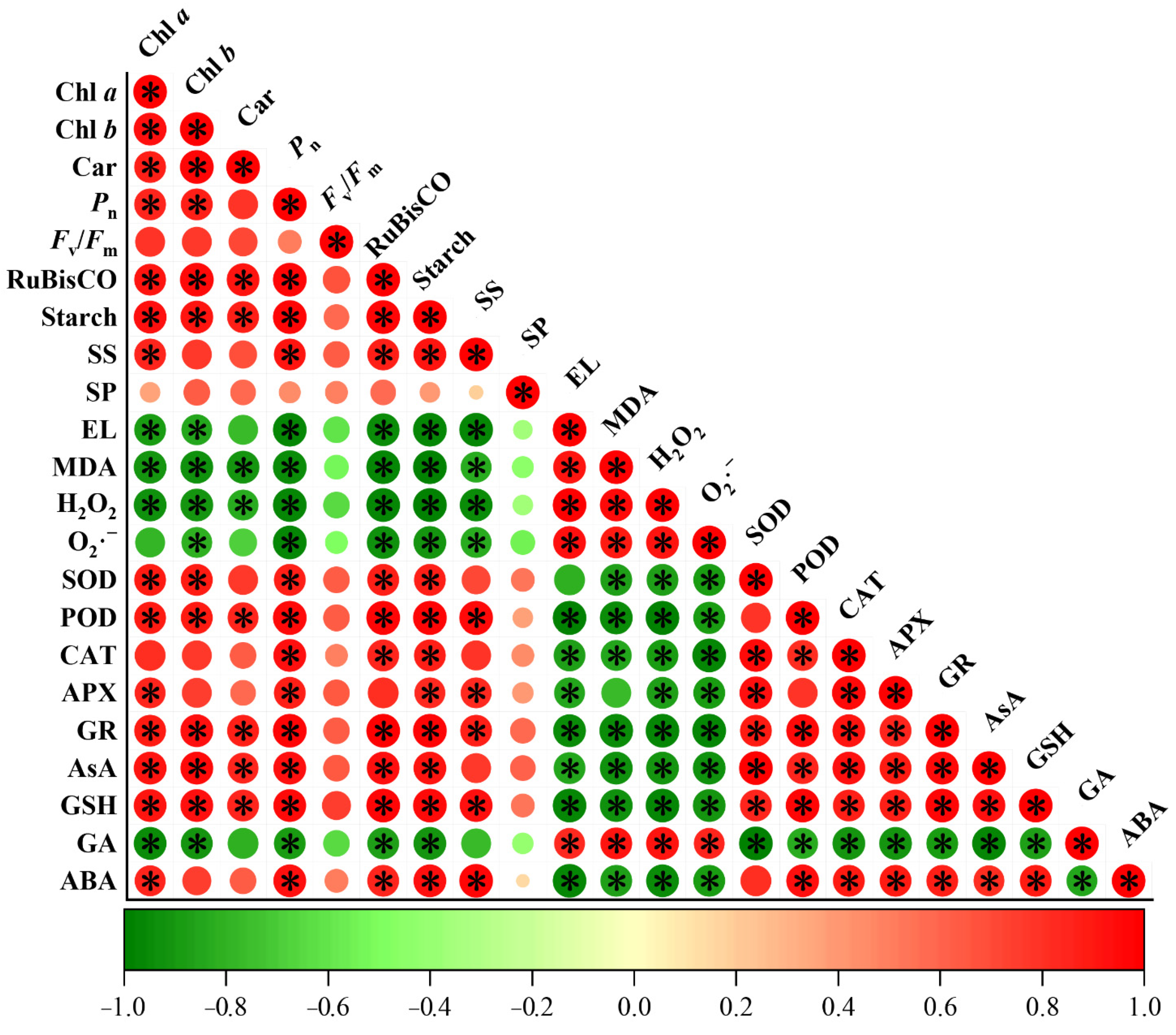
2. Results
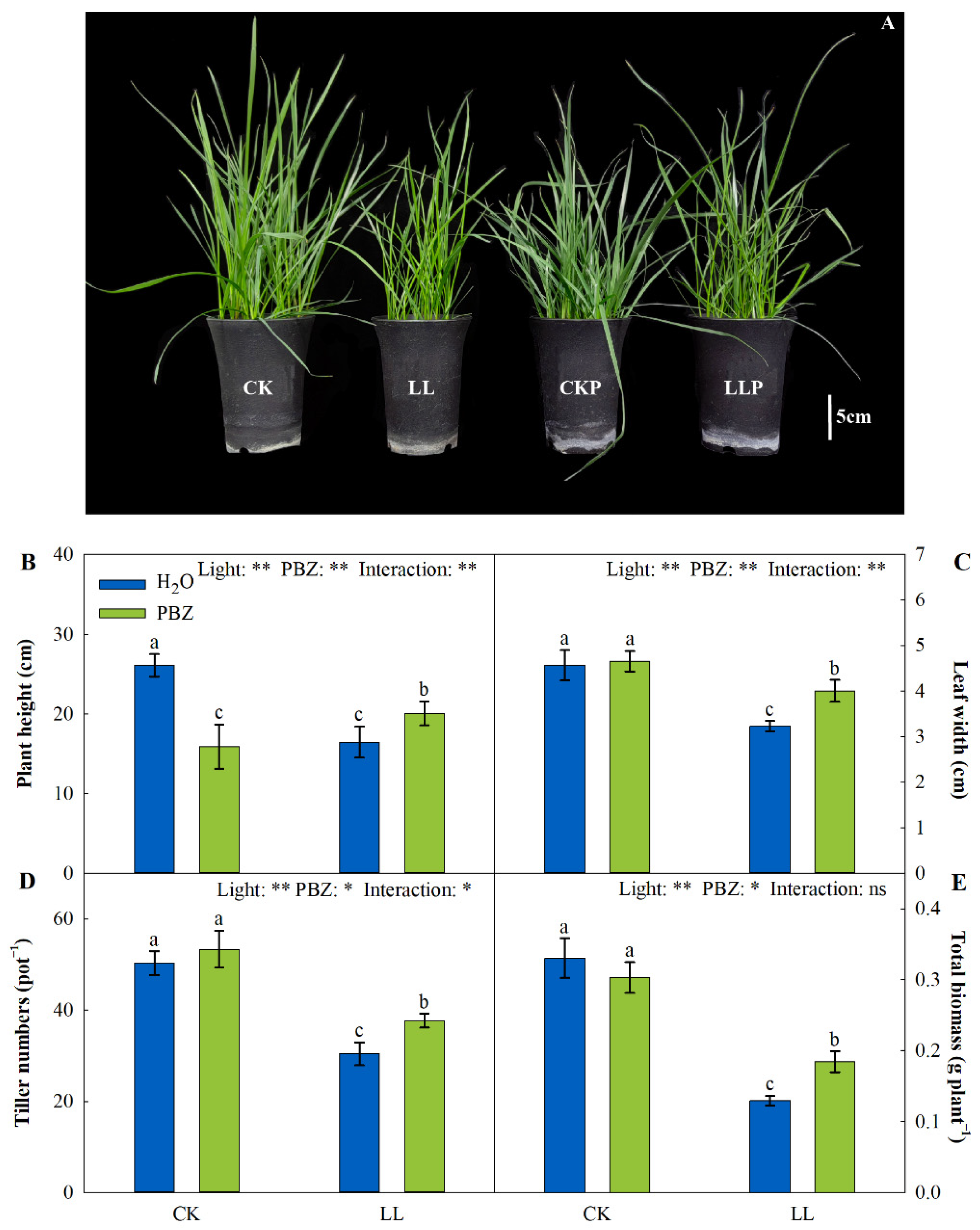
2.1. Optimal PBZ Concentration and Morphological Characteristics
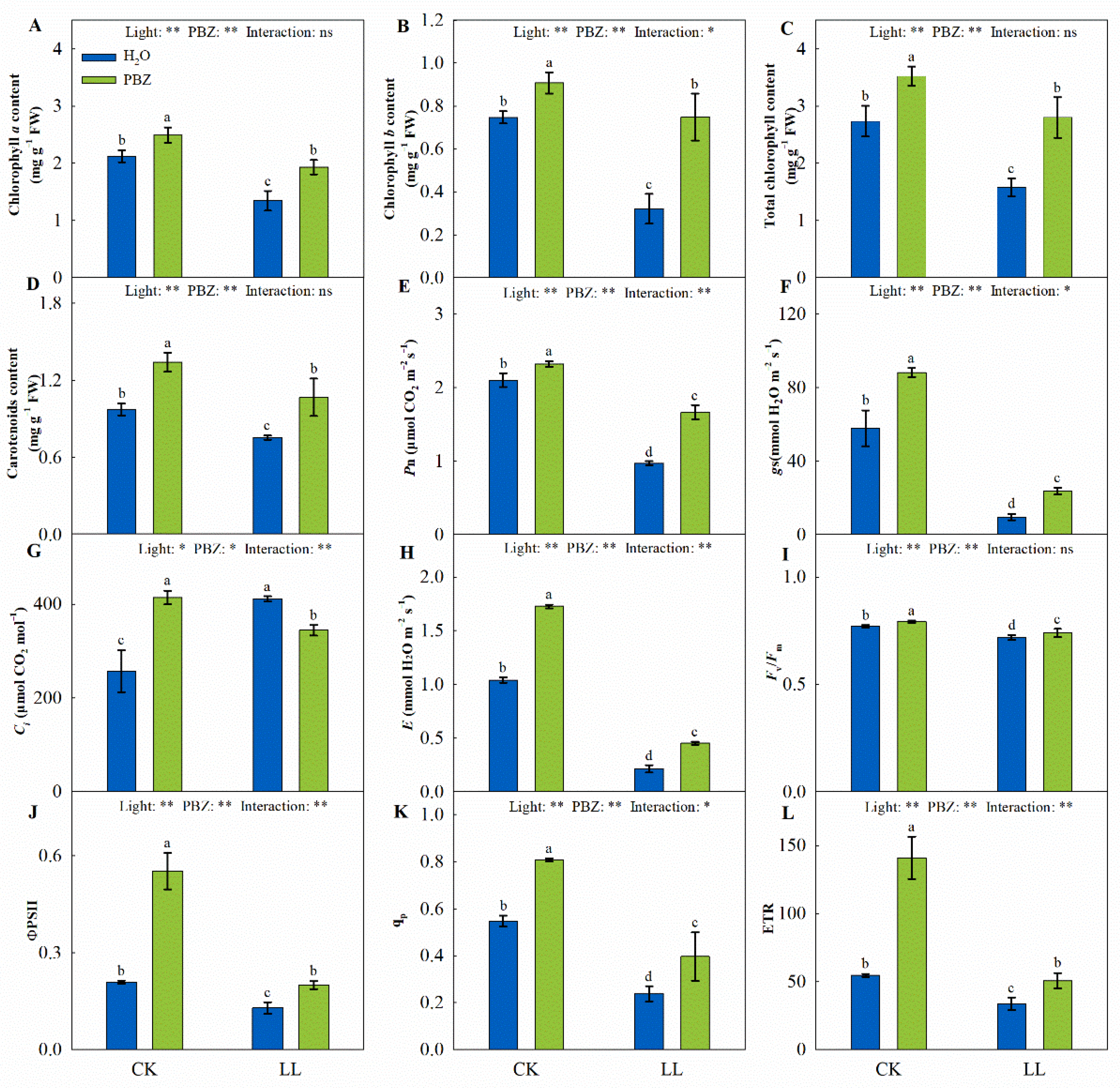
2.2. Chlorophyll Content, Gas Exchange, and Chlorophyll Fluorescence Parameters
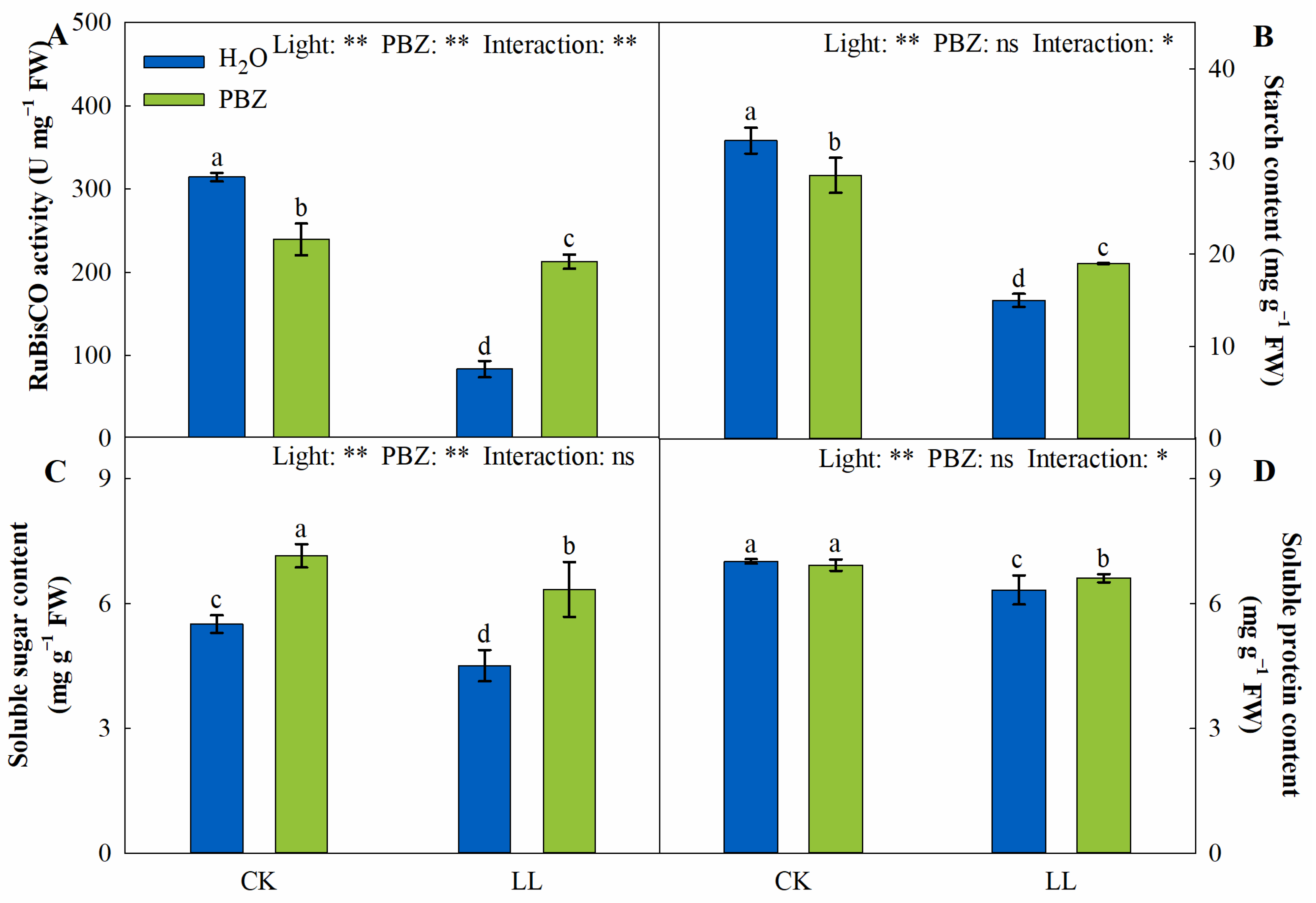
2.3. RuBisCO Activity and Photosynthates
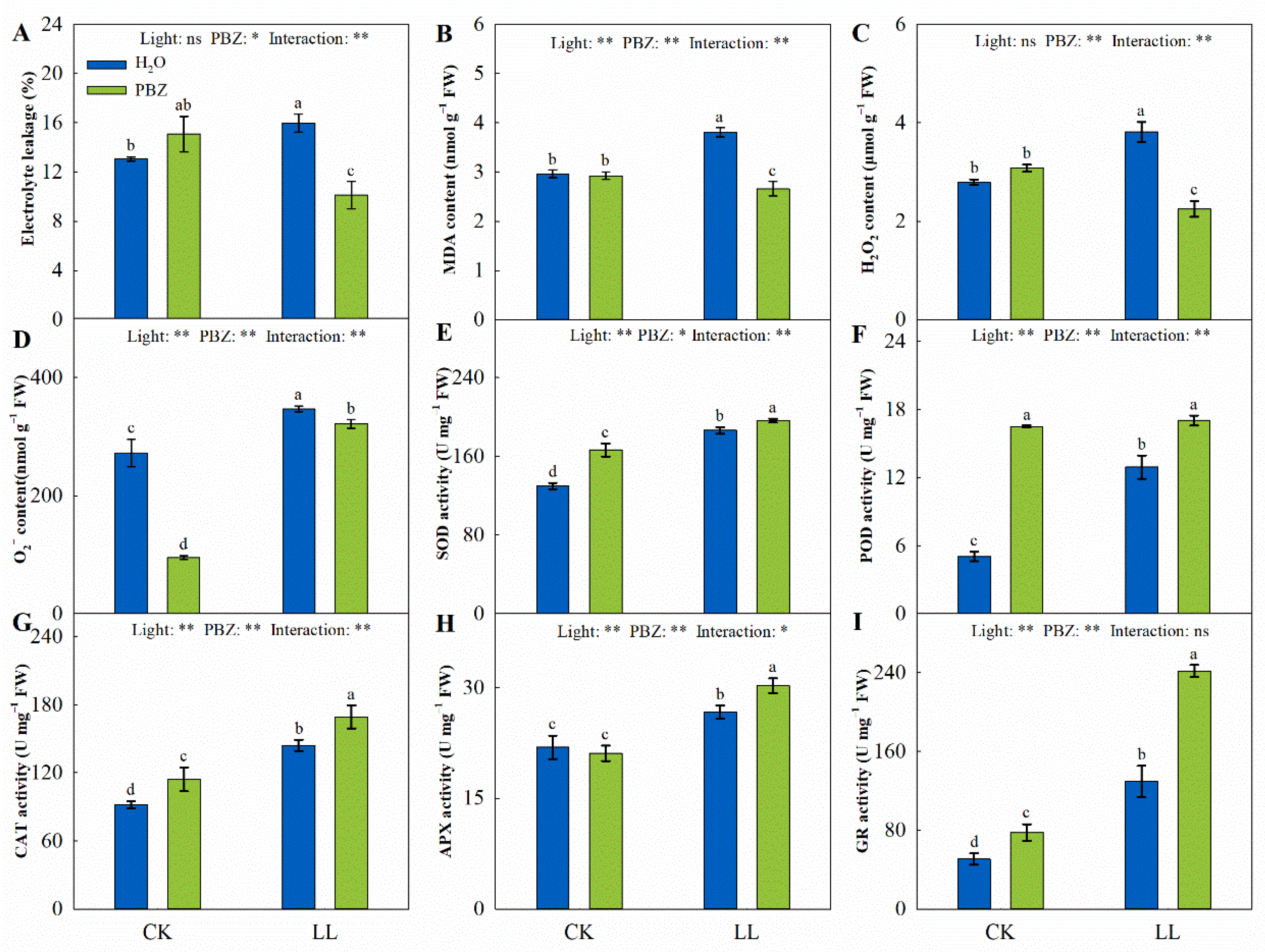
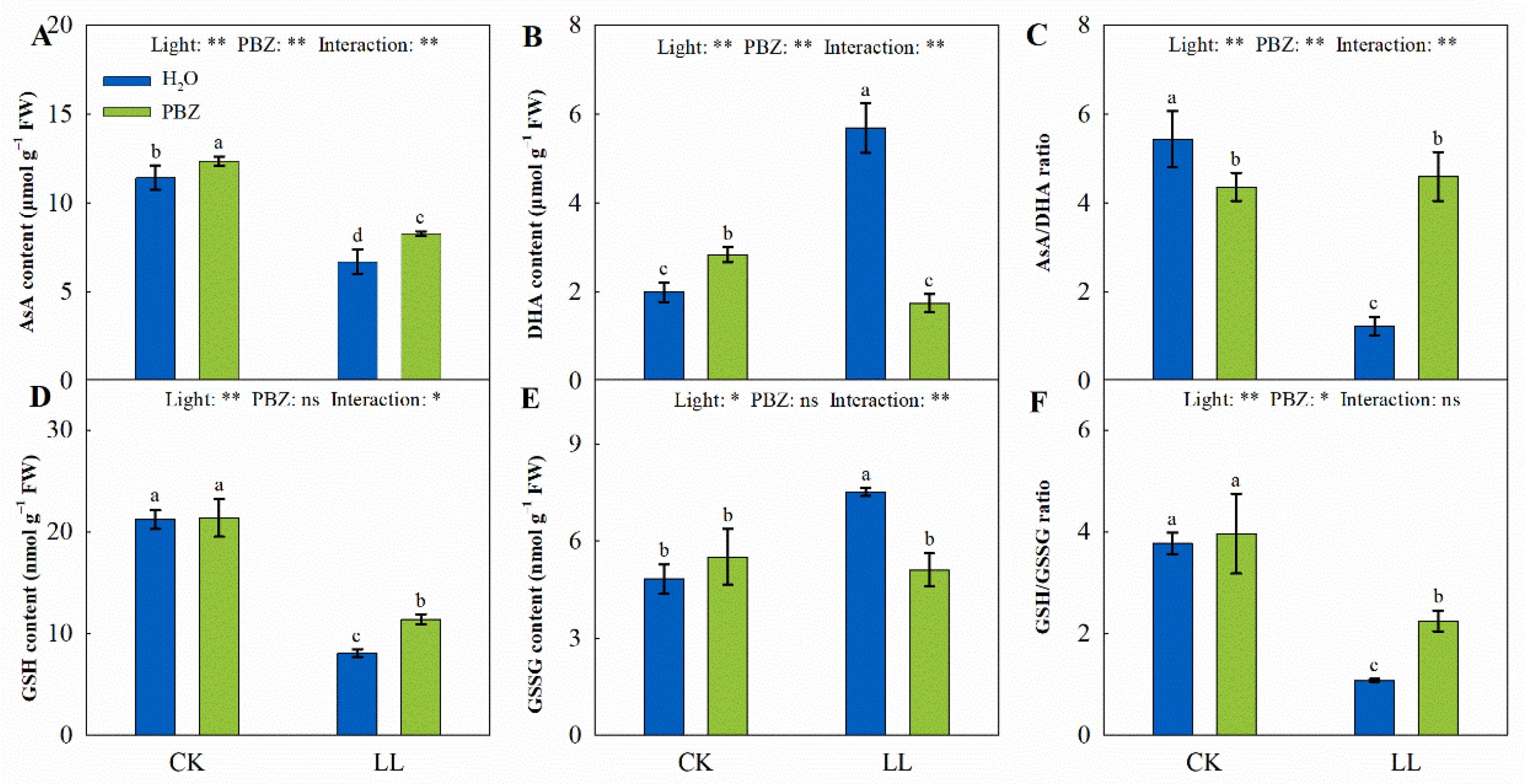
2.4. Membrane Damage and Oxidative Stress
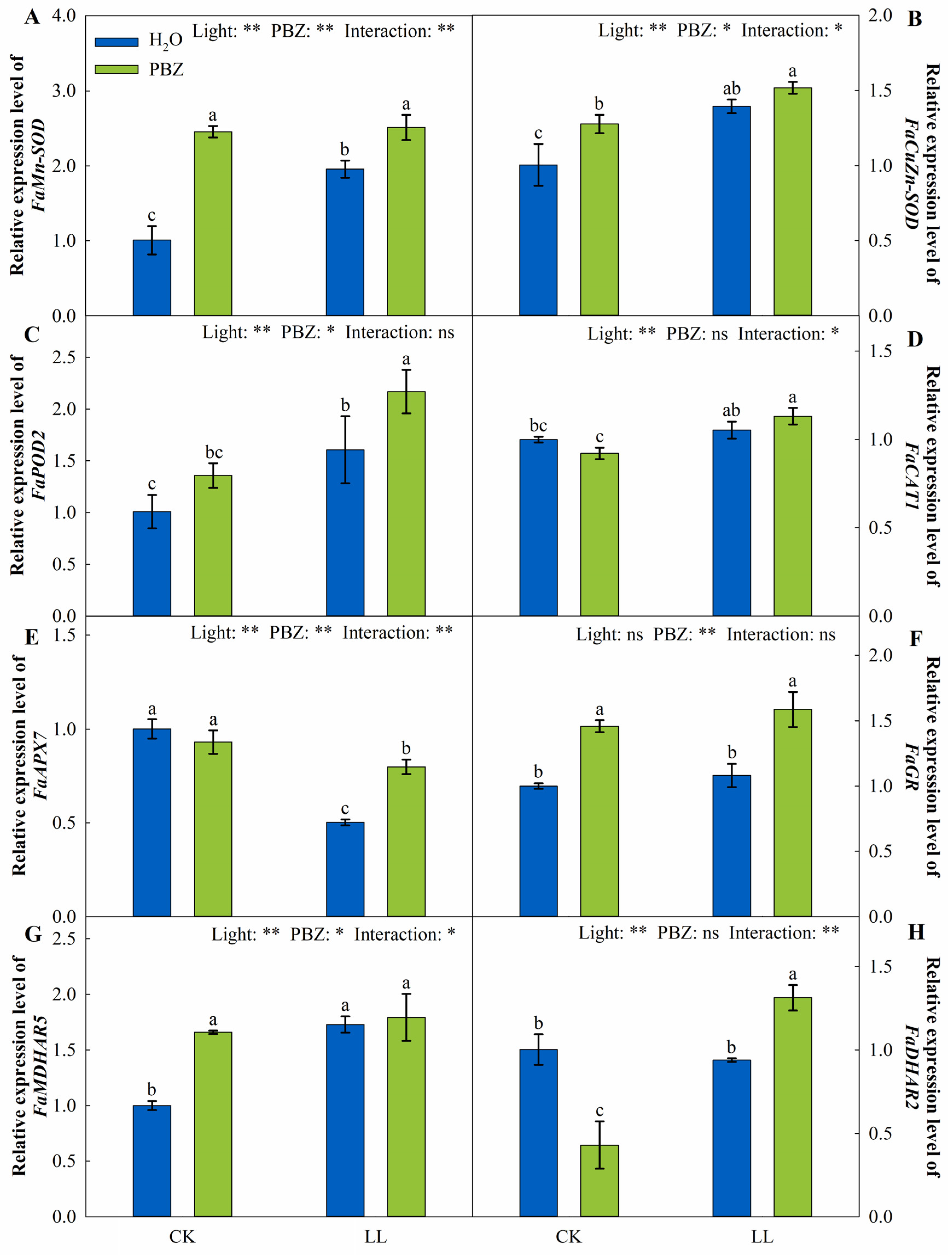
2.5. Antioxidant System
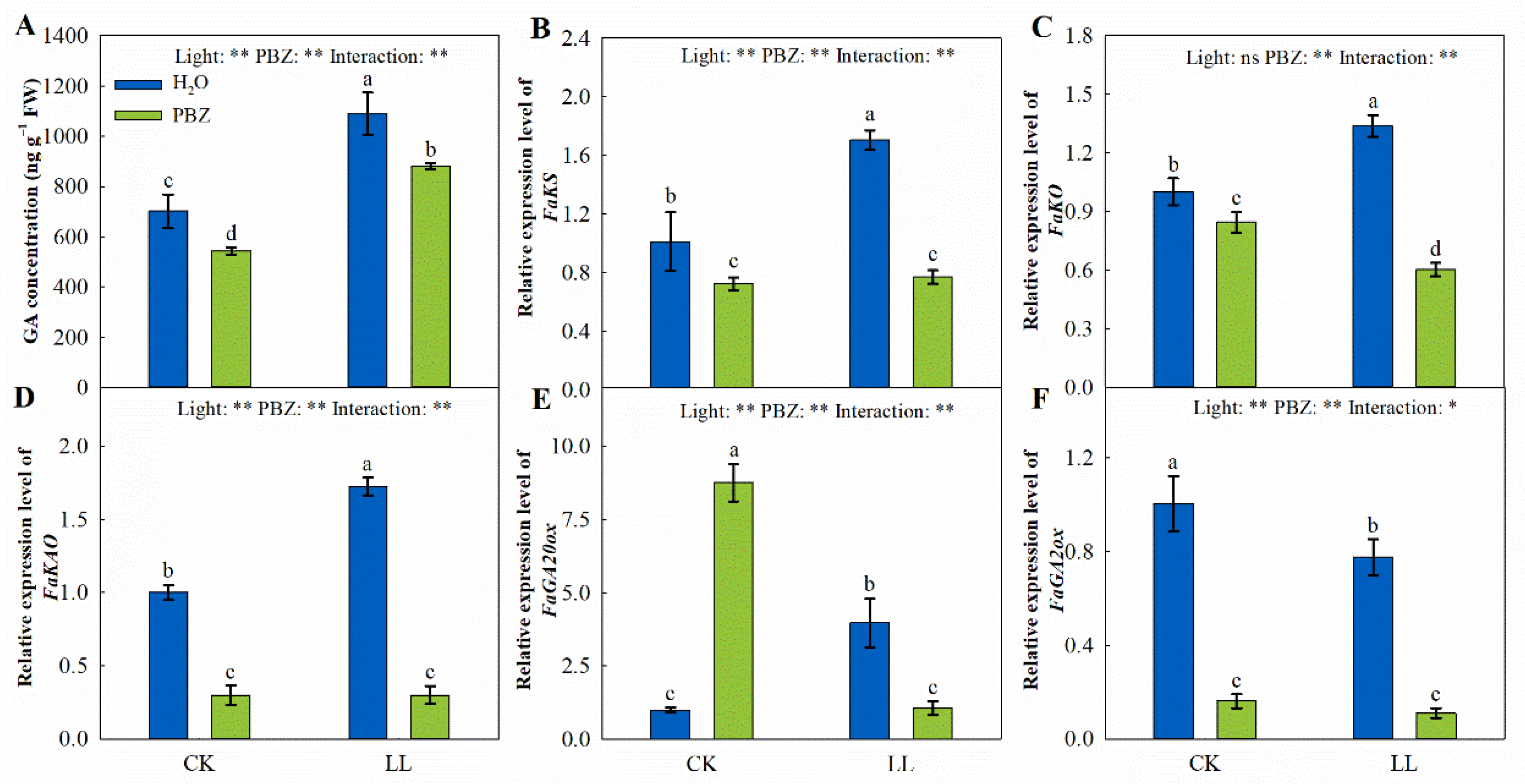
2.6. Endogenous GA Concentration and GA Metabolism
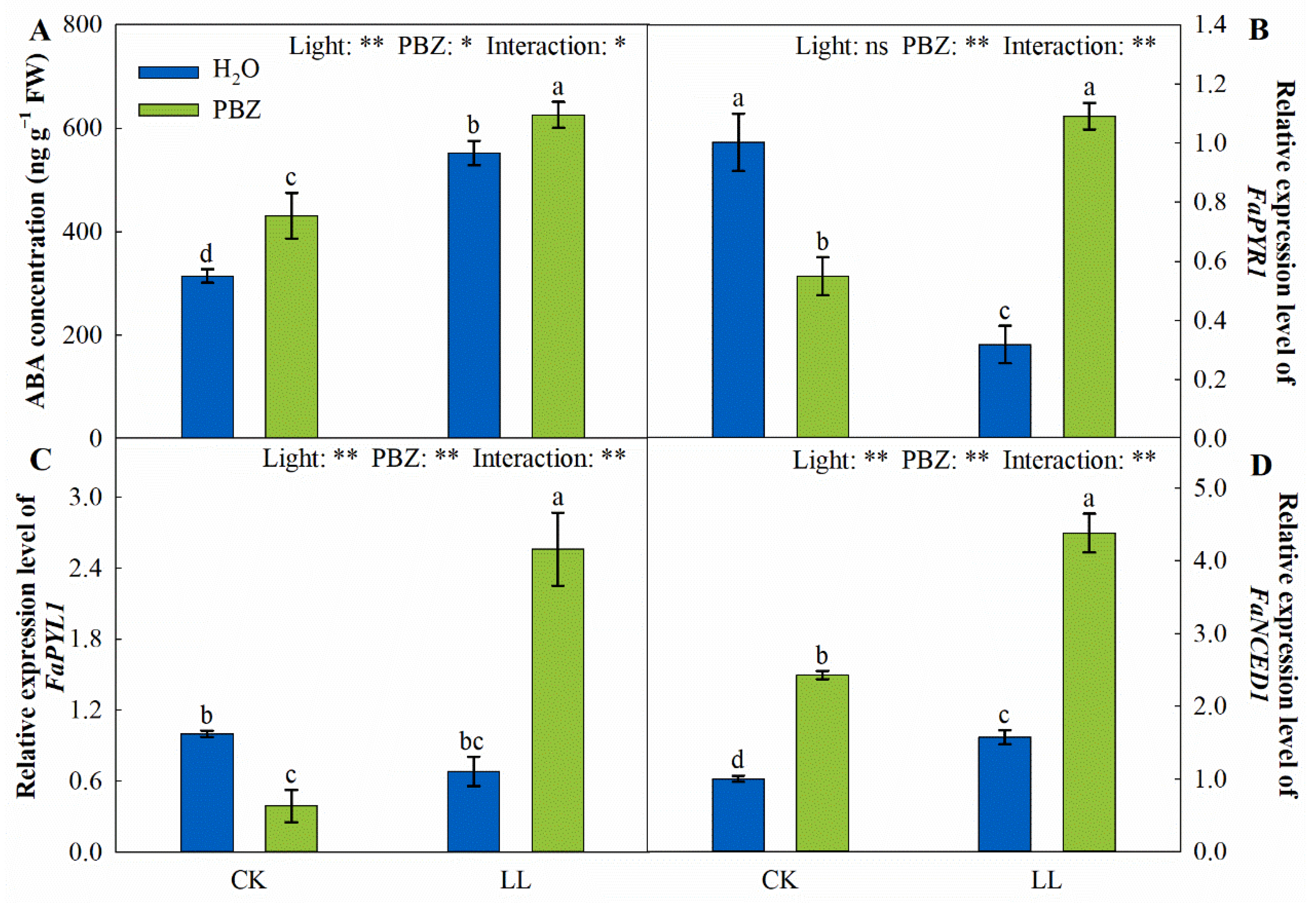
2.7. Endogenous ABA Concentration and ABA-Related Genes Expression
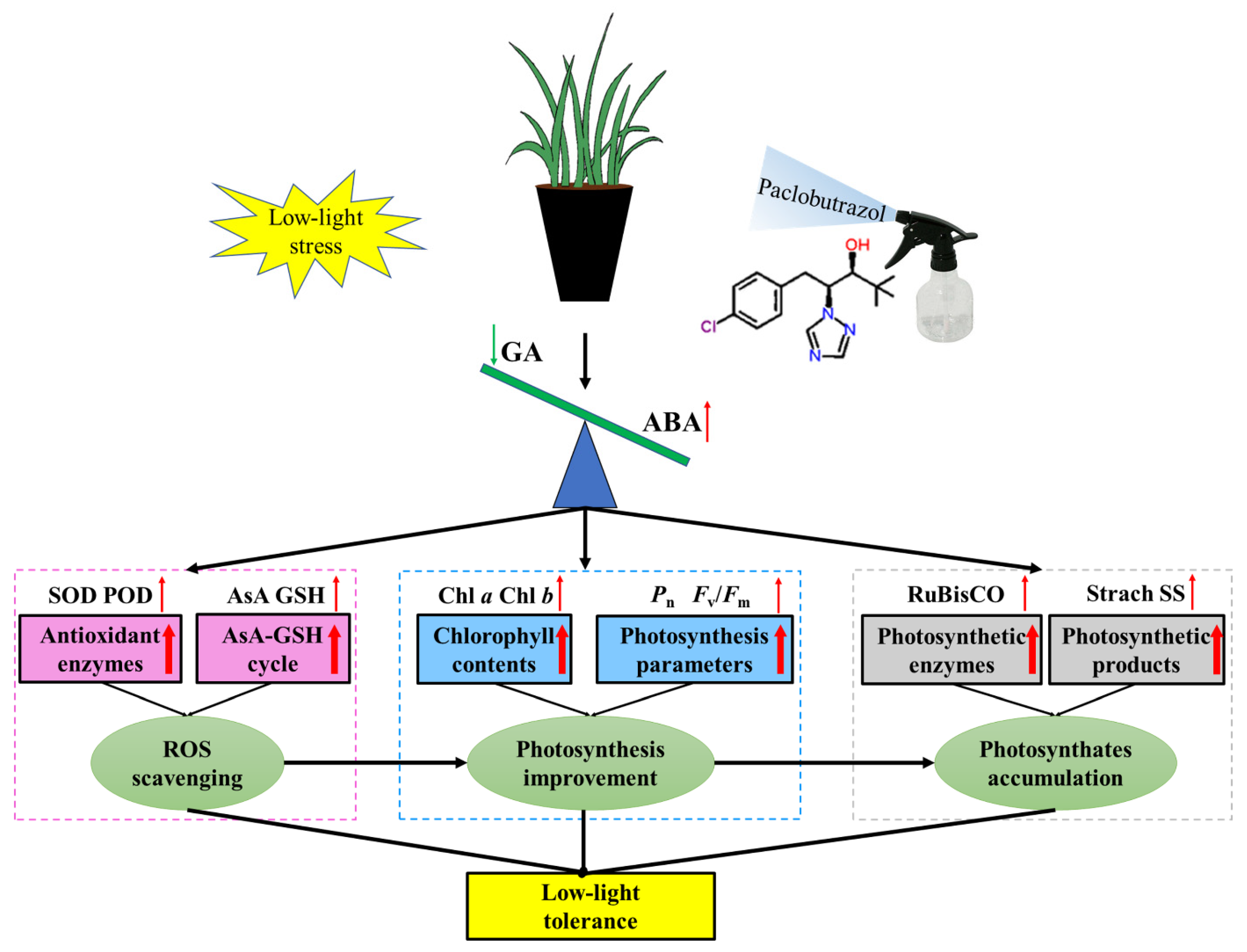
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Experimental Designs and Treatments
4.3. Measurements of Growth and Morphological Parameters
4.4. Determination of Chlorophyll Content, Photosynthesis, and Chlorophyll Fluorescence Parameters
4.5. Quantification of RuBisCO Activity and Photosynthetic Products
4.6. Estimation of the Ion Leakage and Oxidative Damage
4.7. Determination of the Activity of the Antioxidant Enzymes
4.8. Determination of Non-Enzymatic Antioxidants
4.9. Estimation of GA and ABA Concentrations
4.10. Quantitative Real-Time PCR (qRT-PCR) Analysis
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, L.L.; Yu, J.H.; Liao, W.B.; Zhang, G.B.; Xie, J.M.; Lv, J.; Xiao, X.M.; Yang, B.L.; Zhou, R.H.; Bu, R.F. Moderate ammonium: Nitrate alleviates low light intensity stress in mini Chinese cabbage seedling by regulating root architecture and photosynthesis. Sci. Hortic. 2015, 186, 143–153. [Google Scholar] [CrossRef]
- Shao, Q.S.; Wang, H.Z.; Guo, H.P.; Zhou, A.C.; Huang, Y.Q.; Sun, Y.L.; Li, M.Y. Effects of shade treatments on photosynthetic characteristics, chloroplast ultrastructure, and physiology of Anoectochilus roxburghii. PLoS ONE 2014, 9, e85996. [Google Scholar] [CrossRef]
- Emmons, R.; Rossi, F. Turfgrass Science and Management; Cengage: Stamford, CT, USA, 2016. [Google Scholar]
- Yao, X.D.; Zhou, H.L.; Zhu, Q.; Li, C.H.; Zhang, H.J.; Wu, J.J.; Xie, F.T. Photosynthetic response of soybean leaf to wide light-fluctuation in maize-soybean intercropping system. Front. Plant Sci. 2017, 8, 1695. [Google Scholar] [CrossRef] [PubMed]
- Du, J.B.; Jiang, H.K.; Sun, X.; Li, Y.; Liu, Y.; Sun, M.Y.; Fan, Z.; Cao, Q.L.; Feng, L.Y.; Shang, J.; et al. Auxin and gibberellins are required for the receptor-like kinase ERECTA regulated hypocotyl elongation in shade avoidance in Arabidopsis. Front. Plant Sci. 2018, 9, 124. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.J.; Feng, B.L.; Zhou, Y.F.; Liu, C.; Gong, X.W. Exogenous brassinosteroids increases tolerance to shading by altering stress responses in mung bean (Vigna radiata L.). Photosynth. Res. 2022, 151, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Wherley, B.G.; Gardner, D.S.; Metzger, J.D. Tall fescue photomorphogenesis as influenced by changes in the spectral composition and light intensity. Crop Sci. 2005, 45, 562–568. [Google Scholar] [CrossRef]
- Trappe, J.M.; Karcher, D.E.; Richardson, M.D.; Patton, A.J. Shade and traffic tolerance varies for bermudagrass and zoysiagrass cultivars. Crop Sci. 2011, 51, 870–877. [Google Scholar] [CrossRef]
- Suzuki, N.; Koussevitzky, S.; Mittler, R.; Miller, G. ROS and redox signalling in the response of plants to abiotic stress. Plant Cell Environ. 2012, 35, 259–270. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Bhuyan, M.H.M.B.; Zulfiqar, F.; Raza, A.; Mohsin, S.M.; Fujita, M.; Fotopoulos, V. Reactive oxygen species and antioxidant defense in plants under abiotic stress: Revisiting the crucial role of a universal defense regulator. Antioxidants 2020, 9, 681. [Google Scholar] [CrossRef]
- Glab, T.; Szewczyk, W.; Gondek, K.; Knaga, J.; Tomasik, M.; Kowalik, K. Effect of plant growth regulators on visual quality of turfgrass. Sci. Hortic. 2020, 267, 109314. [Google Scholar] [CrossRef]
- Desta, B.; Amare, G. Paclobutrazol as a plant growth regulator. Chem. Biol. Technol. Agric. 2021, 8, 1–15. [Google Scholar] [CrossRef]
- March, S.R.; Martins, D.; McElroy, J.S. Growth inhibitors in turfgrass. Planta Daninha 2013, 31, 733–747. [Google Scholar] [CrossRef]
- Zheng, K.J.; Wang, Y.T.; Wang, S.C. The non-DNA binding bHLH transcription factor Paclobutrazol Resistances are involved in the regulation of ABA and salt responses in Arabidopsis. Plant Physiol. Biochem. 2019, 139, 239–245. [Google Scholar] [CrossRef] [PubMed]
- Sasi, M.; Awana, M.; Samota, M.K.; Tyagi, A.; Kumar, S.; Sathee, L.; Krishnan, V.; Praveen, S.; Singh, A. Plant growth regulator induced mitigation of oxidative burst helps in the management of drought stress in rice (Oryza sativa L.). Environ. Exp. Bot. 2021, 185, 104413. [Google Scholar] [CrossRef]
- Babarashi, E.; Rokhzadi, A.; Pasari, B.; Mohammadi, K. Ameliorating effects of exogenous paclobutrazol and putrescine on mung bean [Vigna radiata (L.) Wilczek] under water deficit stress. Plant Soil Environ. 2021, 67, 40–45. [Google Scholar] [CrossRef]
- Xu, Y.F.; Fu, J.J.; Chu, X.T.; Sun, Y.F.; Zhou, H.; Hu, T.M. Nitric oxide mediates abscisic acid induced light-tolerance in leaves of tall fescue under high-light stress. Sci. Hortic. 2013, 162, 1–10. [Google Scholar] [CrossRef]
- Dunne, J.C.; Miller, G.L.; Arellano, C.; Brandenburg, R.L.; Schoeman, A.; Milla-Lewis, S.R. Shade response of bermudagrass accessions under different management practices. Urban For. Urban Green. 2017, 26, 169–177. [Google Scholar] [CrossRef]
- Wang, Q.M.; Hou, F.Y.; Dong, S.X.; Xie, B.T.; Li, A.X.; Zhang, H.Y.; Zhang, L.M. Effects of shading on the photosynthetic capacity, endogenous hormones and root yield in purple-fleshed sweetpotato (Ipomoea batatas (L.) Lam). Plant Growth Regul. 2014, 72, 113–122. [Google Scholar] [CrossRef]
- Xie, F.C.; Shi, Z.J.; Zhang, G.Y.; Zhang, C.T.; Sun, X.Y.; Yan, Y.; Zhao, W.; Guo, Z.X.; Zhang, L.; Fahad, S.; et al. Quantitative leaf anatomy and photophysiology systems of C3 and C4 turfgrasses in response to shading. Sci. Hortic. 2020, 274, 109674. [Google Scholar] [CrossRef]
- Falcioni, R.; Moriwaki, T.; Bonato, C.M.; de Souza, L.A.; Nanni, M.R.; Antunes, W.C. Distinct growth light and gibberellin regimes alter leaf anatomy and reveal their influence on leaf optical properties. Environ. Exp. Bot. 2017, 140, 86–95. [Google Scholar] [CrossRef]
- Yan, D.L.; Song, F.; Li, Z.; Sharma, A.; Xie, X.T.; Wu, T.T.; Wang, X.F.; He, Y.; Chen, J.B.; Huang, Q.Y.; et al. Application of titanium regulates the functional components of photosynthetic apparatus in grafted seedlings of Carya cathayensis Sarg. under shade. Chemosphere 2022, 290, 133301. [Google Scholar] [CrossRef] [PubMed]
- Cohen, I.; Netzer, Y.; Sthein, I.; Gilichinsky, M.; Tel-Or, E. Plant growth regulators improve drought tolerance, reduce growth and evapotranspiration in deficit irrigated Zoysia japonica under field conditions. Plant Growth Regul. 2019, 88, 9–17. [Google Scholar] [CrossRef]
- Namjoyan, S.; Rajabi, A.; Sorooshzadeh, A.; AghaAlikhani, M. The potential of tebuconazole for mitigating oxidative stress caused by limited irrigation and improving sugar yield and root quality traits in sugar beet. Plant Physiol. Biochem. 2021, 162, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.Y.; Zheng, Q.J.; Xiong, L.B.; Xie, F.C.; Li, X.; Li, Y.; Zhang, L.; Saud, S.; Guo, Z.X.; Yan, Y.; et al. Nitrogen assimilation and gene regulation of two Kentucky bluegrass cultivars differing in response to nitrate supply. Sci. Hortic. 2021, 288, 110315. [Google Scholar] [CrossRef]
- Mohan, R.; Kaur, T.; Bhat, H.A.; Khajuria, M.; Pal, S.; Vyas, D. Paclobutrazol induces photochemical efficiency in mulberry (Morus alba L.) under water stress and affects leaf yield without influencing biotic interactions. J. Plant Growth Regul. 2020, 39, 205–215. [Google Scholar] [CrossRef]
- Anderson, J.M.; Park, Y.I.; Chow, W.S. Photoinactivation and photoprotection of photosystem II in nature. Physiol. Plant. 1997, 100, 214–223. [Google Scholar] [CrossRef]
- Fu, W.G.; Li, P.P.; Wu, Y.Y. Effects of different light intensities on chlorophyll fluorescence characteristics and yield in lettuce. Sci. Hortic. 2012, 135, 45–51. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef]
- Hou, J.F.; Xu, Y.; Wang, Z.X.; Chen, F.; Yuan, L.Y.; Zhu, S.D.; Shan, G.L.; Wang, C.G. Exogenous paclobutrazol can relieve the low irradiance stress in Capsicum annuum seedlings. Biol. Plant. 2021, 65, 297–306. [Google Scholar] [CrossRef]
- Sage, R.F.; Monson, R.K.; Ehleringer, J.R.; Adachi, S.; Pearcy, R.W. Some like it hot: The physiological ecology of C4 plant evolution. Oecologia 2018, 187, 941–966. [Google Scholar] [CrossRef]
- Raines, C.A. Transgenic approaches to manipulate the environmental responses of the C3 carbon fixation cycle. Plant Cell Environ. 2006, 29, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Taylor, S.H.; Gonzalez-Escobar, E.; Page, R.; Parry, M.A.J.; Long, S.P.; Carmo-Silva, E. Faster than expected Rubisco deactivation in shade reduces cowpea photosynthetic potential in variable light conditions. Nat. Plants 2022, 8, 118–124. [Google Scholar] [CrossRef] [PubMed]
- Wen, S.Y.; Liu, B.W.; Long, S.; Gao, S.H.; Liu, Q.; Liu, T.Y.; Xu, Y.F. Low nitrogen level improves low-light tolerance in tall fescue by regulating carbon and nitrogen metabolism. Environ. Exp. Bot. 2022, 194, 104749. [Google Scholar] [CrossRef]
- Gollan, P.J.; Lima-Melo, Y.; Tiwari, A.; Tikkanen, M.; Aro, E.M. Interaction between photosynthetic electron transport and chloroplast sinks triggers protection and signalling important for plant productivity. Philos. Trans. R. Soc. B 2017, 372, 20160390. [Google Scholar] [CrossRef]
- Virsile, A.; Brazaityte, A.; Vastakaite-Kairiene, V.; Miliauskiene, J.; Jankauskiene, J.; Novickovas, A.; Samuoliene, G. Lighting intensity and photoperiod serves tailoring nitrate assimilation indices in red and green baby leaf lettuce. J. Sci. Food Agric. 2019, 99, 6608–6619. [Google Scholar] [CrossRef]
- Adeel, M.; Ma, C.X.; Ullah, S.; Rizwan, M.; Hao, Y.; Chen, C.Y.; Jilani, G.; Shakoor, N.; Li, M.S.; Wang, L.H.; et al. Exposure to nickel oxide nanoparticles insinuates physiological, ultrastructural and oxidative damage: A life cycle study on Eisenia fetida. Environ. Pollut. 2019, 254, 113032. [Google Scholar] [CrossRef]
- Zhang, X.H.; Liu, Y.H.; Liu, Q.; Zong, B.; Yuan, X.P.; Sun, H.E.; Wang, J.; Zang, L.; Ma, Z.Z.; Liu, H.M.; et al. Nitric oxide is involved in abscisic acid-induced photosynthesis and antioxidant system of tall fescue seedlings response to low-light stress. Environ. Exp. Bot. 2018, 155, 226–238. [Google Scholar] [CrossRef]
- Zhou, X.P.; Tan, Z.M.; Zhou, Y.G.; Guo, S.R.; Sang, T.; Wang, Y.; Shu, S. Physiological mechanism of strigolactone enhancing tolerance to low light stress in cucumber seedlings. BMC Plant Biol. 2022, 22, 30. [Google Scholar] [CrossRef]
- Davari, K.; Rokhzadi, A.; Mohammadi, K.; Pasari, B. Paclobutrazol and amino acid-based biostimulant as beneficial compounds in alleviating the drought stress effects on Safflower (Carthamus tinctorius L.). J. Soil. Sci. Plant Nut. 2021, 22, 1–17. [Google Scholar] [CrossRef]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Sharma, I.; Ching, E.; Saini, S.; Bhardwaj, R.; Pati, P.K. Exogenous application of brassinosteroid offers tolerance to salinity by altering stress responses in rice variety Pusa Basmati-1. Plant Physiol. Biochem. 2013, 69, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Srivastav, M.; Kishor, A.; Dahuja, A.; Sharma, R.R. Effect of paclobutrazol and salinity on ion leakage, proline content and activities of antioxidant enzymes in mango (Mangifera indica L.). Sci. Hortic. 2010, 125, 785–788. [Google Scholar] [CrossRef]
- Nagar, S.; Singh, V.P.; Arora, A.; Dhakar, R.; Singh, N.; Singh, G.P.; Meena, S.; Kumar, S.; Ramakrishnan, R.S. Understanding the role of gibberellic acid and paclobutrazol in terminal heat stress tolerance in wheat. Front. Plant Sci. 2021, 12, 692252. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.Q.; Zhang, H.J.; Cao, Y.Y.; Weeda, S.; Ren, S.X.; Guo, Y.D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2015, 66, 647–656. [Google Scholar] [CrossRef] [PubMed]
- ElSayed, A.I.; Rafudeen, M.S.; Gomaa, A.M.; Hasanuzzama, M. Exogenous melatonin enhances the reactive oxygen species metabolism, antioxidant defense-related gene expression, and photosynthetic capacity of Phaseolus vulgaris L. to confer salt stress tolerance. Physiol. Plant. 2021, 173, 1369–1381. [Google Scholar] [CrossRef]
- Sofy, M.R.; Elhindi, K.M.; Farouk, S.; Alotaibi, M.A. Zinc and paclobutrazol mediated regulation of growth, upregulating antioxidant aptitude and plant productivity of pea plants under salinity. Plants 2020, 9, 1197. [Google Scholar] [CrossRef] [PubMed]
- Hajihashemi, S.; Kiarostami, K.; Saboora, A.; Enteshari, S. Exogenously applied paclobutrazol modulates growth in salt-stressed wheat plants. Plant Growth Regul. 2007, 53, 117–128. [Google Scholar] [CrossRef]
- Colebrook, E.H.; Thomas, S.G.; Phillips, A.L.; Hedden, P. The role of gibberellin signalling in plant responses to abiotic stress. J. Exp. Biol. 2014, 217, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Hisamatsu, T.; King, R.W.; Helliwell, C.A.; Koshioka, M. The involvement of gibberellin 20-oxidase genes in phytochrome-regulated petiole elongation of Arabidopsis. Plant Physiol. 2005, 138, 1106–1116. [Google Scholar] [CrossRef] [Green Version]
- Hedden, P. The current status of research on gibberellin biosynthesis. Plant Cell Physiol. 2020, 61, 1832–1849. [Google Scholar] [CrossRef] [PubMed]
- Baninasab, B.; Ghobadi, C. Influence of paclobutrazol and application methods on high-temperature stress injury in cucumber seedlings. J. Plant Growth Regul. 2011, 30, 213–219. [Google Scholar] [CrossRef]
- Wang, P.C.; Du, Y.Y.; Hou, Y.J.; Zhao, Y.; Hsu, C.C.; Yuan, F.J.; Zhu, X.H.; Tao, W.A.; Song, C.P.; Zhu, J.K. Nitric oxide negatively regulates abscisic acid signaling in guard cells by S-nitrosylation of OST1. Proc. Natl. Acad. Sci. USA 2015, 112, 613–618. [Google Scholar] [CrossRef] [PubMed]
- Soumya, P.R.; Kumar, P.; Pal, M. Paclobutrazol: A novel plant growth regulator and multi-stress ameliorant. Indian J. Plant Physiol. 2017, 22, 267–278. [Google Scholar] [CrossRef]
- Liu, X.; Hou, X.L. Antagonistic regulation of ABA and GA in metabolism and signaling pathways. Front. Plant Sci. 2018, 9, 251. [Google Scholar] [CrossRef]
- Opio, P.; Tomiyama, H.; Saito, T.; Ohkawa, K.; Ohara, H.; Kondo, S. Paclobutrazol elevates auxin and abscisic acid, reduces gibberellins and zeatin and modulates their transporter genes in Marubakaido apple (Malus prunifolia Borkh. var. ringo Asami) rootstocks. Plant Physiol. Biochem. 2020, 155, 502–511. [Google Scholar] [CrossRef]
- Liu, B.W.; Zhang, X.H.; You, X.K.; Li, Y.Y.; Long, S.; Wen, S.Y.; Liu, Q.; Liu, T.Y.; Guo, H.; Xu, Y.F. Hydrogen sulfide improves tall fescue photosynthesis response to low-light stress by regulating chlorophyll and carotenoid metabolisms. Plant Physiol. Biochem. 2022, 170, 133–145. [Google Scholar] [CrossRef]
- Fu, J.J.; Sun, Y.F.; Chu, X.T.; Yang, L.Y.; Xu, Y.F.; Hu, T.M. Exogenous nitric oxide alleviates shade-induced oxidative stress in tall fescue (Festuca arundinacea Schreb.). J. Hortic. Sci. Biotechnol. 2014, 89, 193–200. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Van Kooten, O.; Snel, J.F.H. The use of chlorophyll fluorescence nomenclature in plant stress physiology. Photosynth. Res. 1990, 25, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Barrios, A.C.; Medina-Velo, I.A.; Zuverza-Mena, N.; Dominguez, O.E.; Peralta-Videa, J.R.; Gardea-Torresdey, J.L. Nutritional quality assessment of tomato fruits after exposure to uncoated and citric acid coated cerium oxide nanoparticles, bulk cerium oxide, cerium acetate and citric acid. Plant Physiol. Biochem. 2017, 110, 100–107. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Xu, C.; Yin, Y.; Cai, R.; Wang, P.; Ni, Y.; Guo, J.; Chen, E.; Cai, T.; Cui, Z.; Liu, T.; et al. Responses of photosynthetic characteristics and antioxidative metabolism in winter wheat to post-anthesis shading. Photosynthetica 2013, 51, 139–150. [Google Scholar] [CrossRef]
- Song, L.L.; Ding, W.; Shen, J.; Zhang, Z.G.; Bi, Y.R.; Zhang, L.X. Nitric oxide mediates abscisic acid induced thermotolerance in the calluses from two ecotypes of reed under heat stress. Plant Sci. 2008, 175, 826–832. [Google Scholar] [CrossRef]
- Kumar, G.N.M.; Knowles, N.R. Changes in lipid peroxidation and lipolytic and free-radical scavenging enzyme activities during aging and sprouting of potato (Solanum tuberosum) seed-tubers. Plant Physiol. 1993, 102, 115–124. [Google Scholar] [CrossRef]
- Sun, C.L.; Lu, L.L.; Liu, L.J.; Liu, W.J.; Yu, Y.; Liu, X.X.; Hu, Y.; Jin, C.W.; Lin, X.Y. Nitrate reductase-mediated early nitric oxide burst alleviates oxidative damage induced by aluminum through enhancement of antioxidant defenses in roots of wheat (Triticum aestivum). New Phytol. 2014, 201, 1240–1250. [Google Scholar] [CrossRef]
- Ke, D.S.; Sun, G.C.; Wang, Z.X. Effects of superoxide radicals on ACC synthase activity in chilling-stressed etiolated mungbean seedlings. Plant Growth Regul. 2007, 51, 83–91. [Google Scholar] [CrossRef]
- Zhou, W.; Zhao, D.; Lin, X. Effects of waterlogging on nitrogen accumulation and alleviation of waterlogging damage by application of nitrogen fertilizer and mixtalol in winter rape (Brassica napus L.). J. Plant Growth Regul. 1997, 16, 47–53. [Google Scholar] [CrossRef]
- Quintanilla-Guerrero, F.; Duarte-Vazquez, M.A.; Garcia-Almendarez, B.E.; Tinoco, R.; Vazquez-Duhalt, R.; Regalado, C. Polyethylene glycol improves phenol removal by immobilized turnip peroxidase. Bioresour. Technol. 2008, 99, 8605–8611. [Google Scholar] [CrossRef]
- Nakano, Y.; Asada, K. Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol. 1981, 22, 867–880. [Google Scholar] [CrossRef]
- Zhu, H.; Cao, Z.X.; Zhang, L.; Trush, M.A.; Li, Y.B. Glutathione and glutathione-linked enzymes in normal human aortic smooth muscle cells: Chemical inducibility and protection against reactive oxygen and nitrogen species-induced injury. Mol. Cell. Biochem. 2007, 301, 47–59. [Google Scholar] [CrossRef]
- Zhang, J.X.; Kirkham, M.B. Antioxidant responses to drought in sunflower and sorghum seedlings. New Phytol. 1996, 132, 361–373. [Google Scholar] [CrossRef]
- Rao, M.V.; Hale, B.A.; Ormrod, D.P. Amelioration of ozone-induced oxidative damage in wheat plants grown under high carbon dioxide-role of antioxidant enzymes. Plant Physiol. 1995, 109, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.C.; Li, M.L.; Jannasch, A.H.; Jiang, Y.W. Submergence stress alters fructan and hormone metabolism and gene expression in perennial ryegrass with contrasting growth habits. Environ. Exp. Bot. 2020, 179, 104202. [Google Scholar] [CrossRef]
- Li, M.L.; Jannasch, A.H.; Jiang, Y.W. Growth and hormone alterations in response to heat stress in perennial ryegrass accessions differing in heat tolerance. J. Plant Growth Regul. 2020, 39, 1022–1029. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Plant Height (cm) | Leaf Width (mm) | Total Biomass (g plant−1) | Total Chl Content (mg g−1 FW) | MDA Content (nmol g−1 FW) |
|---|---|---|---|---|---|
| CK | 26.27 ± 3.92 a | 4.75 ± 0.05 a | 0.59 ± 0.06 a | 3.73 ± 0.24 a | 2.46 ± 0.05 c d |
| LL | 16.53 ± 1.36 c | 2.74 ± 0.09 c | 0.09 ± 0.02 c | 2.32 ± 0.04 d | 2.73 ± 0.04 b |
| LL + 50 | 16.90 ± 1.31 c | 3.36 ± 0.19 c | 0.09 ± 0.01 c | 2.24 ± 0.17 d | 2.63 ± 0.14 b c |
| LL + 100 | 15.64 ± 3.06 c | 3.50 ± 0.16 c | 0.08 ± 0.03 c | 2.59 ± 0.09 c | 2.93 ± 0.04 a |
| LL + 200 | 21.07 ± 0.51 b | 3.78 ± 0.16 b | 0.16 ± 0.01 b | 3.01 ± 0.15 b | 2.30 ± 0.15 d |
| LL + 300 | 17.14 ± 2.45 c | 3.24 ± 0.08 c | 0.08 ± 0.02 c | 2.79 ± 0.05 c | 2.66 ± 0.14 b |
| LL + 500 | 16.14 ± 1.88 c | 2.99 ± 0.16 c | 0.09 ± 0.01 c | 2.28 ± 0.10 c | 2.62 ± 0.02 b c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Long, S.; Liu, K.; Zhu, T.; Gong, J.; Gao, S.; Wang, R.; Zhang, L.; Liu, T.; Xu, Y. Paclobutrazol Ameliorates Low-Light-Induced Damage by Improving Photosynthesis, Antioxidant Defense System, and Regulating Hormone Levels in Tall Fescue. Int. J. Mol. Sci. 2022, 23, 9966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179966
Liu B, Long S, Liu K, Zhu T, Gong J, Gao S, Wang R, Zhang L, Liu T, Xu Y. Paclobutrazol Ameliorates Low-Light-Induced Damage by Improving Photosynthesis, Antioxidant Defense System, and Regulating Hormone Levels in Tall Fescue. International Journal of Molecular Sciences. 2022; 23(17):9966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179966
Chicago/Turabian StyleLiu, Bowen, Si Long, Kening Liu, Tianqi Zhu, Jiongjiong Gong, Shuanghong Gao, Ruijia Wang, Liyun Zhang, Tieyuan Liu, and Yuefei Xu. 2022. "Paclobutrazol Ameliorates Low-Light-Induced Damage by Improving Photosynthesis, Antioxidant Defense System, and Regulating Hormone Levels in Tall Fescue" International Journal of Molecular Sciences 23, no. 17: 9966. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23179966