1. Introduction
Calcium plays an essential role in maintaining the stability of cell walls, cell membranes, and membrane-binding proteins. As a second messenger, calcium is widely involved in regulating plant growth, development, and responses to external stimuli [
1]. Signal transduction pathways are initiated at the cell’s perception of external signals. Different environmental stimuli are regarded as primary signals; they are recognized by specific receptors and are transmitted to intracellular second messengers, inducing a downstream cascade reaction [
2]. Plants respond to physiological and biochemical changes and elicit subsequent biological effects. Based on structure and function, two categories of receptors recognize Ca
2+ signals. One is the transitive receptor with a Ca
2+-binding domain, which has no kinase activity and which regulates the expression of downstream genes by interacting with other proteins, such as calmodulins, calmodulin-like protein, and calcineurin B-like protein (CBL). These regulate other effector proteins to induce downstream reactions and, thus, transmit signals. The second category comprises responsive receptors that bind to Ca
2+ and exhibit kinase activity. Their functions are mediated by binding to Ca
2+ and inducing conformational changes in calcium-dependent protein kinases [
3].
CBLs belong to the family of serine/threonine protein phosphatases and have a specific Ca
2+-binding domain, also called the EF-hand domain [
4]. In rice, each CBL protein contains four conserved EF-hand domains (
Supplementary Figure S1). In addition, myristoylation and palmitoylation sites exist at the N-terminal region and FPSF motifs at the C-terminus region. The myristoylation and palmitoylation sites at the N-terminal region of the CBL protein help localize the CBL protein [
5]. The serine residues in the FPSF motif are phosphorylated by the CBL-interacting protein kinase (CIPK) to enhance protein–protein interactions and activate the CBL–CIPK complex to target downstream proteins [
6]. During Ca
2+-mediated signal transduction, CBL interacts with Ca
2+-dependent serine/threonine protein kinases, namely CIPK, to transmit signals within cells [
7]. CIPKs are a class of CBL-dependent protein kinases with conserved sequences. Typical CIPK proteins contain an N-terminal kinase domain and C-terminal regulatory domain (
Supplementary Figure S2). An activation loop exists in the N-terminal kinase domain between the Asp–Phe–Gly and Ala–Pro–Glu motifs [
8]. Protein kinases can be activated by phosphorylation of one or more amino acid residues in the activation loop [
9]. The C-terminal regulatory domain contains a NAF motif that specifically binds to CBL [
10]. Under normal conditions, CIPK activity is inhibited by the intramolecular binding of the NAF motif and kinase domain. After receiving a specific signal, the NAF motif binds to CBL, resulting in the activation of CIPK. In addition, toward the C-terminus of the NAF motif exists a protein phosphatase interaction motif that binds to protein phosphatase 2C (PP2C) [
11]. Studies have found that CBL proteins in plants can interact with specific CIPK proteins, CIPK proteins can also interact with specific CBL proteins, and these interactions play an essential role in response to stress as well as plant growth and development [
12]. For example, the interactions of CBL3 and CBL4 with CIPK24 and CBL10 with CIPK24 in
Arabidopsis are responsible for the response to salt stress [
13]. Further, CBL10 and CIPK6 are involved in immune reactions and disease-programmed cell death in tomatoes and tobacco [
14]. The formation of CBL–CIPK complexes, such as the CBL1 and CIPK1 complex, requires the presence of Ca
2+ [
15]. In contrast, other CBLs, such as CBL2 and CBL4, are Ca
2+-independent. For example, CBL2 interacts with CIPK14 and CBL4 with CIPK24 in the absence of Ca
2+ [
16]. CBLs can also interact with CIPKs from different species. For example, the CBL of rice can interact with the CIPK of
Arabidopsis [
17] and Bermuda grass [
18]. Therefore, CBLs and CIPKs are relatively conserved in the plant kingdom.
Gene replication can occur in several ways, including tandem duplication, segmental duplication, transposon element-mediated duplication, and whole-genome duplication (WGD). WGD is also known as polyploidy. Polyploidization of the genome leads to chromosome doubling and eventually the retention of many repetitive genes. These repetitive genes are essential driving forces that lead to the evolution of gene functions [
19]: although rapid gene deletions follow WGD, certain genes with specific functions will eventually remain [
20]. For example, after WGD in
Populus, the enriched genes were related to vesicle and protein-targeted transport, protein degradation, RNA transcription regulation, and protein post-translational modification [
21]. Additionally, transcription factors and signal transduction-related genes were significantly enriched after WGD in rice and
Arabidopsis [
22].
In this study, the phylogenetic and synteny analysis of CBL and CIPK were carried out to determine whether CBL and CIPK could be retained in WGD. OsCBL2 and OsCBL8 were selected with great differences of the promoter region and gene structure, were identified from their interacting CIPK proteins using yeast two hybrid (Y2H) and bimolecular fluorescence complementation (BiFC), and were analyzed for their spatiotemporal expression pattern. CBL8 can regulate seed germination through gene editing materials of rice. Transcriptome analysis with overexpression lines of OsCBL8 and OsCIPK17 can understand the potential mechanism underlying the regulation of seedling development. Furthermore, the downstream protein of OsCIPK17 was further screened and verified for its response to high temperature and hormones. In general, this study aims to deepen our understanding of the mechanism by which the CBL–CIPK module regulates rice growth, development, and resistance, and it provides a theoretical basis for breeding excellent varieties.
3. Discussion
Cellular calcium signaling regulates almost every aspect of eukaryotic physiology (Tang et al., 2020). CBL–CIPK, an important calcium-signaling module, has attracted considerable attention from botanists since it was first discovered in
Arabidopsis [
29]. We investigated the number of
CBLs and
CIPKs in some plants, including monocotyledons, dicotyledons, bryophytes, and ferns. The 222 copies of CBL and 711 copies of
CIPK in these plants show significant quantitative differences among different plant species. A single pair of CBL–CIPK is usually present in green algae [
30], and it appears that an increase in CBL–CIPK genes accompanied plant evolution. During evolution, higher plants showed a greater extent of adaptation than lower plants. For example, multiple CBL–CIPK modules play a role in pollen tube growth, a special developmental process unique to flowering plants [
31]. The amplification of
CBL and
CIPK in
Arabidopsis during evolution has been studied in detail in a recent report [
32]. In this study, we demonstrated an increase in CBL–CIPK numbers in rice. Overall, evolutionary forces have determined and maintained the number of CBL–CIPK in different plants.
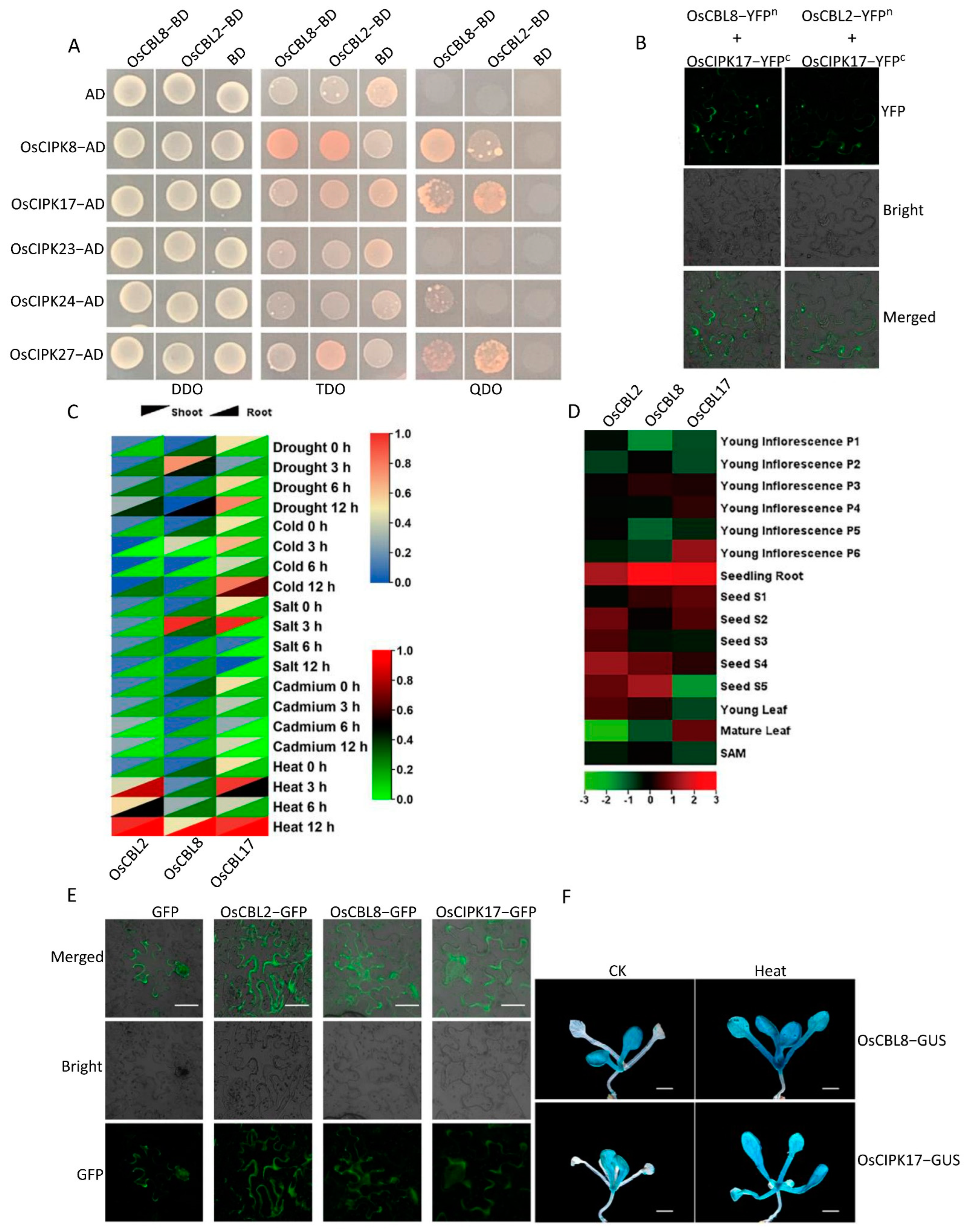
The interaction and expression patterns of CBL and CIPK in rice were analyzed to determine their identity and functional relationships after WGD. The results of the Y2H experiments showed that OsCBL8 interacts with OsCIPK24, whereas OsCBL2 does not. In addition, OsCBL8 and OsCBL2 interact with OsCIPK8, OsCIPK17, and OsCIPK27. However, they show varying degrees of interactions. For instance, the OsCBL8–OsCIPK8 interaction was more robust than that of OsCBL2–OsCIPK8, and the opposite was observed between OsCBL8–OsCIPK17 and OsCBL2–OSCIPK17. These results indicate that CBL and CIPK in rice have specific interactions. Even if the interaction objects between different CBLs are repeated, specificity is maintained by controlling the intensity of interaction. This specificity has also been demonstrated in
Arabidopsis and pumpkins [
32]. Specific interactions between CBL and CIPK have been observed in different species [
17]. To verify this, we selected OsCIPK17, which interacts with both OsCBL8 and OsCBL2, and studied its expression patterns in detail. OsCBL8–OsCIPK17 and OsCBL2–OsCIPK17 showed various trends when exposed to different stresses. For example,
OsCBL2 and
OsCIPK17 showed similar expression patterns in shoots under drought conditions, whereas
OsCBL8 and
OsCIPK17 showed similar expression patterns in roots.
OsCBL8 and
OsCIPK17 exhibited identical expression patterns in young inflorescences at different developmental stages. In addition, the subcellular localization results showed that both OsCBL8 and OsCBL2 were expressed on the cell membrane and interacted with OsCIPK17. The results of GUS tissue staining also suggest that OsCBL8 can interact with OsCIPK17 to regulate plant growth and development and respond to high temperature stress. These results indicate that CBL and CIPK exhibit different spatial and temporal expression patterns at the gene, protein, and tissue levels. The specific interaction and different spatiotemporal expression patterns further imply that OsCBL2 and OsCBL8 have other functions.
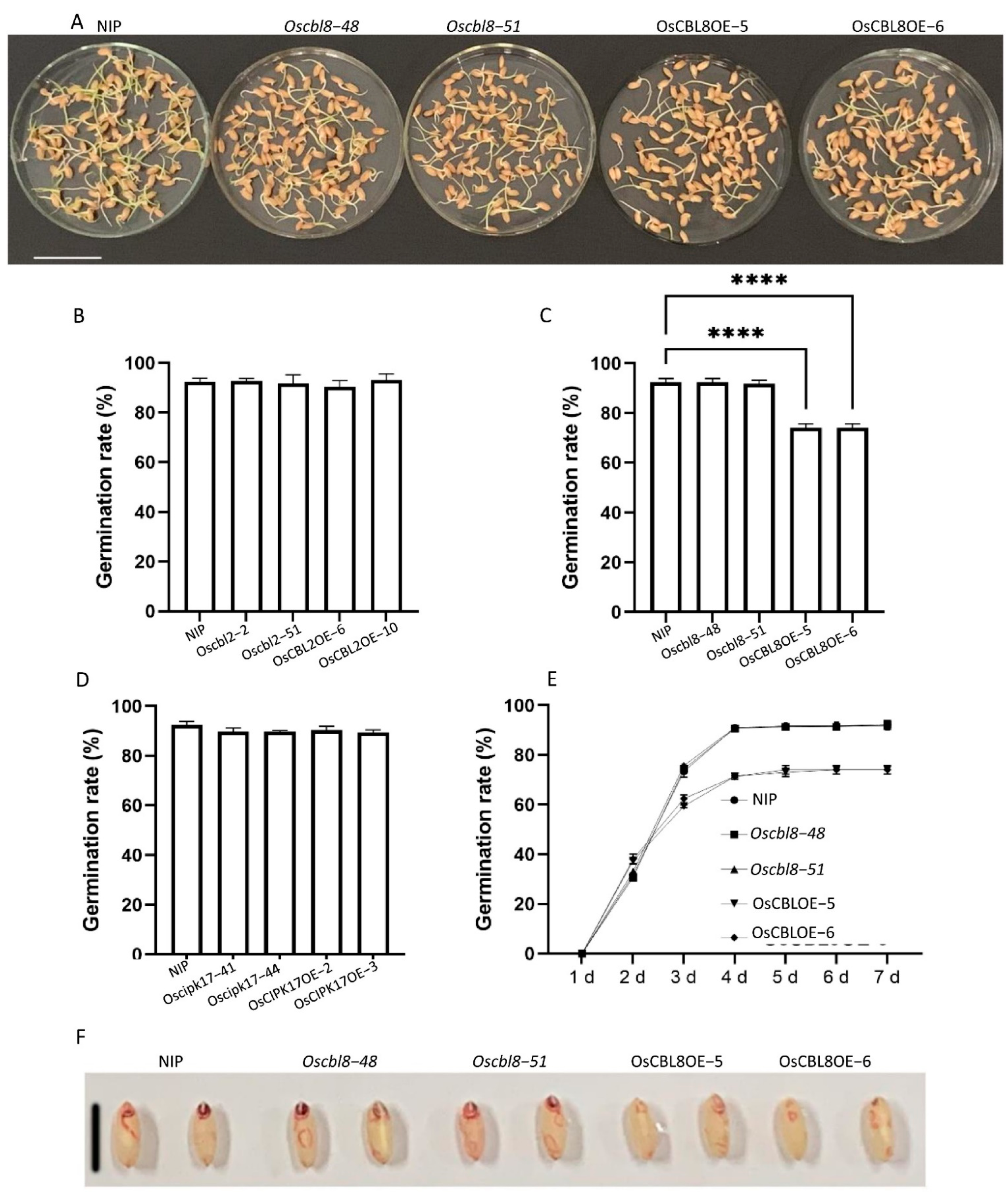
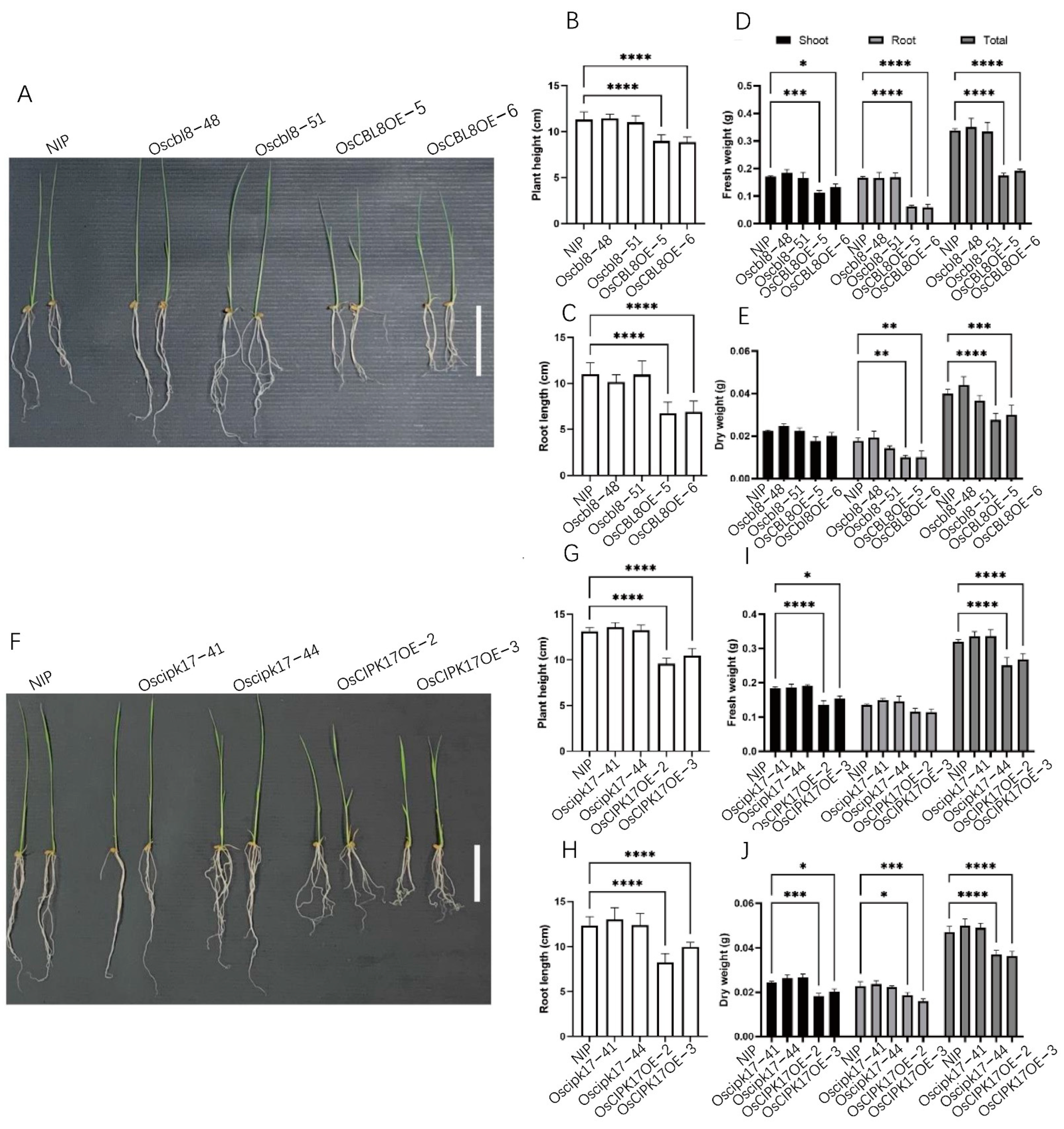
Subsequently, we conducted further studies to determine whether OsCBL8–OsCIPK17 and OsCBL2–OsCIPK17 showed clear functional differences. OsCBL8-OsCIPK17 regulated seedlings growth, whereas OsCBL2–OsCIPK17 did not. Overexpression of
OsCBL8 and
OsCIPK17 resulted in a decrease in physiological indices, such as plant height, root length, and seedlings weight, suggesting that OsCBL8–OsCIPK17 plays a negative regulatory role in seedlings growth. Similar effects of CBL and CIPK on plant growth have been widely reported. For example, CIPK9 is involved in NH4
+-dependent root growth in rice [
33].
CIPK3 is involved in flower development and regulates flowering time [
34]. In addition,
OsCBL8 regulates seed germination.
OsCBL8 overexpression caused a significant decrease in seed germination rates compared with NIP, whereas this phenomenon was not observed with OsCBL2 and OsCIPK17. This suggests that OsCBL8 may combine with a different OsCIPK to affect germination. A similar phenomenon, in which OsCBL10 affected seed germination under waterlogging conditions, has been reported, and variations in the promoter region of
OsCBL10 influenced this regulation [
35]. Overall, by measuring physiological indicators, we found that OsCBL8–OsCIPK17 regulates seedling growth. Additionally, although OsCBL8 and OsCBL2 can interact with OsCIPK17, their functions are significantly different.
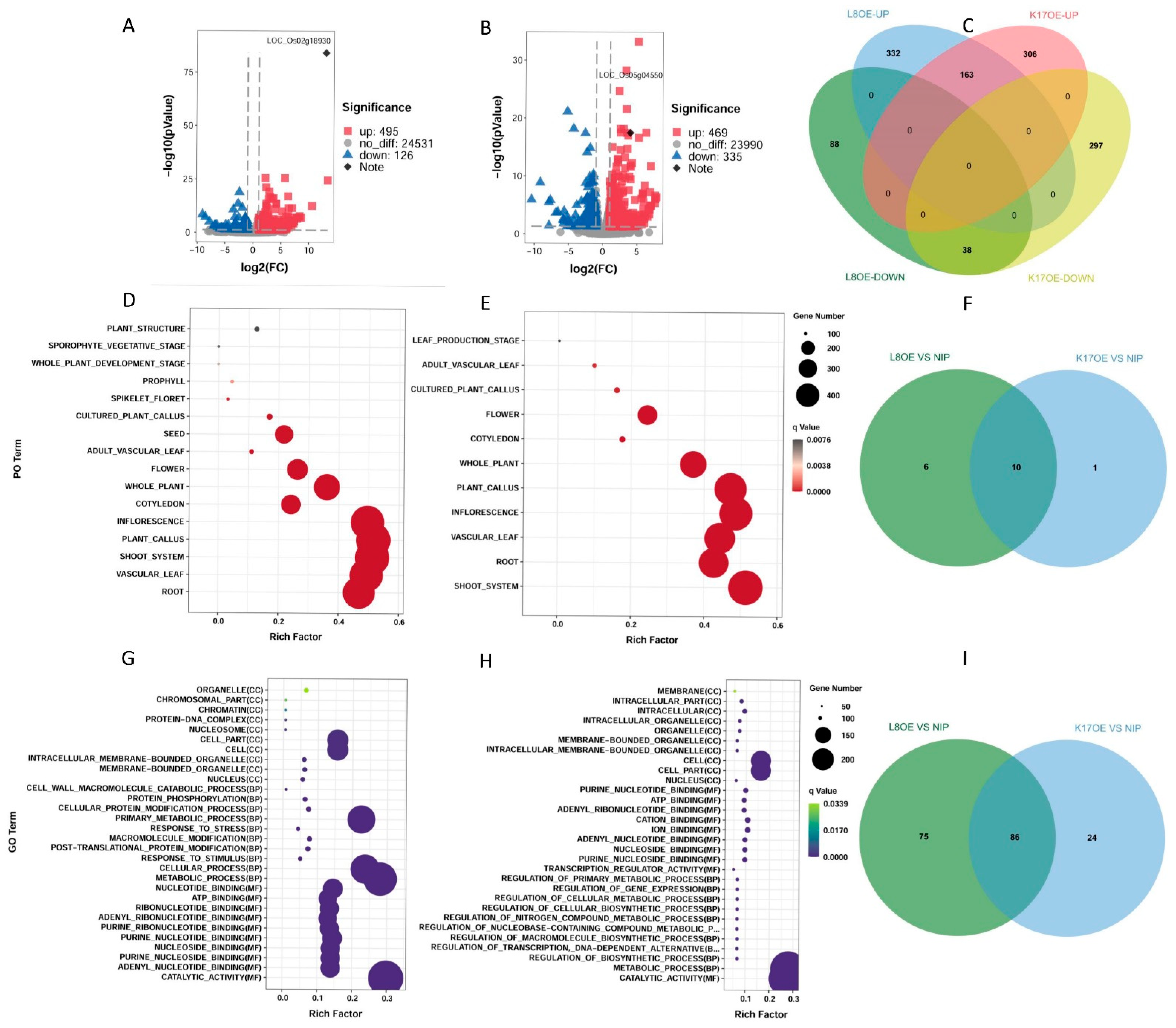
We performed RNA sequencing (RNA-seq) (
Figure 4) on the parent (NIP) and overexpressed lines (
L8OE-5 and
K17OE-3) to explore the effects of OsCBL8–OsCIPK17 on the growth of rice seedlings. We performed DEG calculations to confirm that the expression levels of
OsCBL8 and
OsCIPK17 were significantly upregulated (
Figure 4A,B). There was no intersection between the upregulated and downregulated gene sets between OsCBL8 and OsCIPK17 (
Figure 4C). This suggests that
OsCBL8 and
OsCIPK17 exhibit a highly consistent synergy in gene expression regulation. Since the phenotypes we obtained were low seed germination rate and seedling growth inhibition, we enriched DEGs with PO (
Figure 4D,E). The results showed that the DEGs were enriched in roots, stems, leaves, flowers, and calli. This suggests that OsCBL8–OsCIPK17 may affect the growth and development of these organs, consistent with the observed phenotypic changes (
Figure 4A,C,D). The DEGs were only enriched in the seeds after the overexpression of
OsCBL8. Further research is required to unravel the regulatory role of
OsCBL8 in seed germination. Similar functions of CBL–CIPK have been previously reported.
OsCIPK3 affects flower development [
34], and
OsCIPK9 regulates root growth and development [
33]. Multiple CBL–CIPK modules play a role in pollen tube growth [
31]. GO enrichment analysis showed that DEGs mainly play a role in the synthesis and transport of substances, regulation of enzyme activity, metabolism, response to stimulation, and reproductive development (
Figure 4D,E). This suggests that OsCBL8–OsCIPK17 can regulate the above processes, as reported in previous studies. Various CIPKs regulate the growth and development of flowers and seeds [
31,
33,
36]. For example,
GhCBL2 and
GhCIPK6 are involved in plant sugar homeostasis via interactions with the tonoplast sugar transporter TST21 [
34]. In addition, we used the GSEA method, which is sensitive to non-DEGs, for enrichment analysis of KEGG. This method was also used for PO enrichment to ensure that the PO enrichment results were reliable. Overexpression of
OsCBL8 and
OsCIPK17 showed enrichment for five terms: ABC transporters, ether lipid metabolism, glycerophospholipid metabolism, phagosome proteasome, and SNARE interactions in vesicular transport. ATP-binding cassette (ABC) transporters form one of the most prominent protein families and are widespread in bacteria, archaea, and eukaryotes. They couple ATP hydrolysis to the active transport of various substrates, such as ions, sugars, lipids, sterols, peptides, and proteins [
37]. Some lipids can also promote enzymatic activity [
38]. Glycerophospholipids are the most abundant phospholipids in the body. As components of biofilms, glycerophospholipids are involved in protein recognition and signal transduction [
39]. Phagocytosis is a process involving the uptake of relatively large particles by cells and is a central mechanism in tissue remodeling and defense against infectious agents [
40]. The proteasome is a protein-destroying apparatus involved in many essential cellular functions, such as the regulation of the cell cycle, cell differentiation, signal transduction pathways, antigen processing for appropriate immune responses, stress signaling, and apoptosis [
41]. SNAREs mediate membrane fusion, a fundamental process in cellular biophysics that involves viral infection, endocytosis, and exocytosis [
42]. The results of the KEGG enrichment analysis showed a high degree of consistency with GO enrichment. The enrichment analysis results indicated that OsCBL8–OsCIPK17 could affect the growth of rice seedlings and their responses to external stimuli.
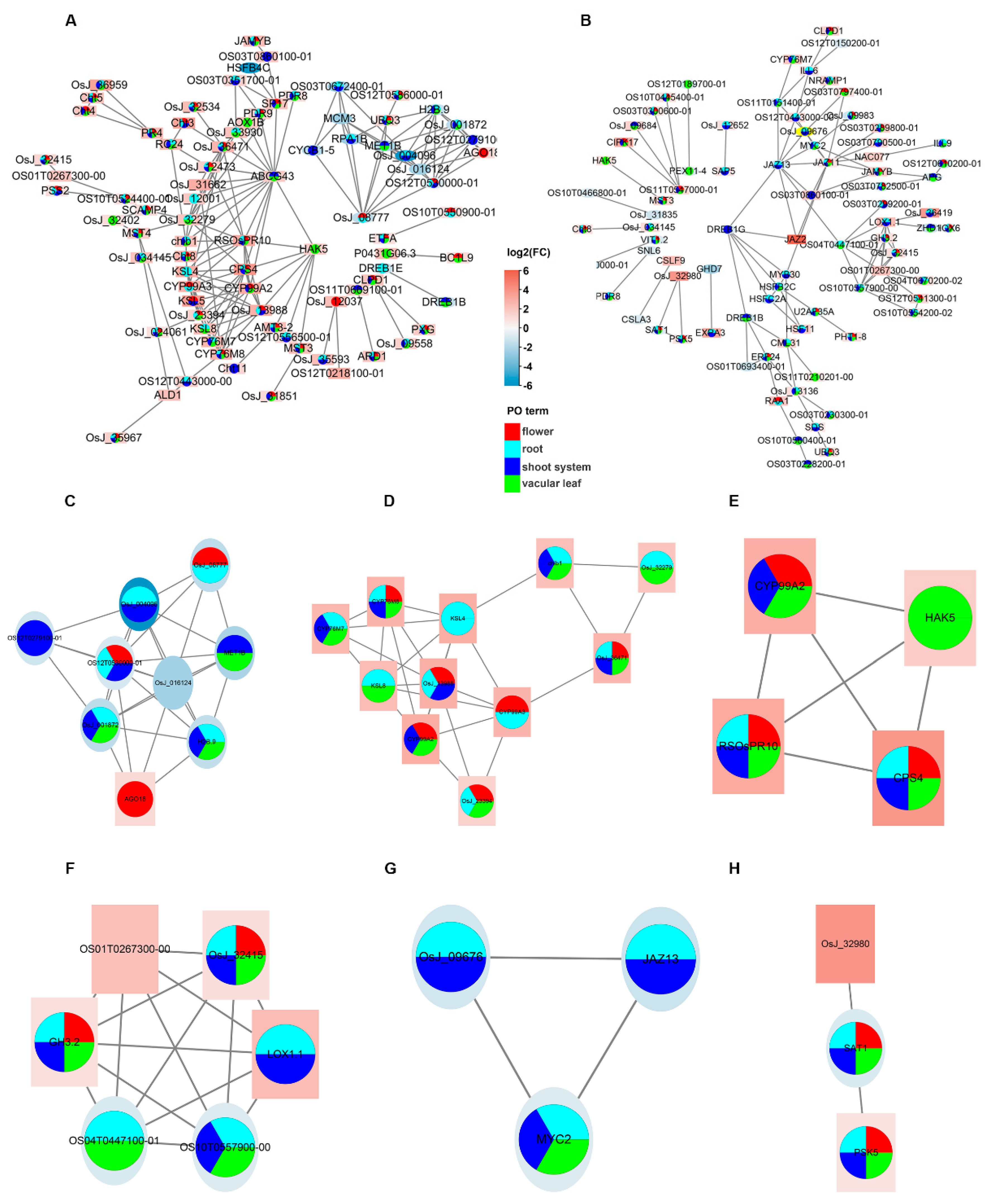
PPI networks were used to analyze the association of DEGs at the protein level. At the same time, we mapped part of the PO enrichment results to the interaction network to predict the impact on the rice phenotype. Many PO terms were mapped onto the PPI network. This shows that OsCBL8–OsCIPK17 participates in rice growth and development. There are many reports that CBL–CIPK is involved in the regulation of plant growth and development [
31,
33,
36]. To understand how OsCBL8–OsCIPK17 regulates growth and development, we explored the sub-modules of the PPI network. Among them, sub-module 1, obtained after overexpression of
OsCBL8, plays a major role in the regulation of RNA expression. The JA-responsive transcription factor JAMYB directly binds to the promoter node AGO18 in sub-module 1 to activate AGO18 transcription. AGO18 is a core RNA-silencing component that promotes rice antiviral defense by isolating miR168 and miR528, which inhibit key antiviral defense proteins [
43]. This confirms that
OsCBL8 regulates AGO18 to enhance seedling disease resistance. This was confirmed by enrichment analysis. A similar phenomenon has been reported in tomatoes, whereby the CBL10–CIPK6 complex is involved in plant immune responses [
14]. Sub-modules 2 and 3 revealed the involvement of chitin and Na
+ in the seedling defense response, respectively. The sub-modules obtained after the overexpression of
OsCIPK17 showed a highly consistent function with overexpression of
OsCBL8. That is, it can regulate the JA-mediated defense response. This implies that OsCBL8–OsCIPK17 can endow the seedlings with disease resistance. In addition, the sub-modules obtained after overexpression of
OsCIPK17 were also involved in auxin metabolism and response to temperature. Similar phenomena have been reported previously. For example, the primary function of
CaCIPK6 is in auxin transport [
44]. A high expression of CIPK gene will lead to thermo-sensitive genic self-incompatible in maize [
45].
PbCBL2 is strongly induced under high temperature stress in pear. In summary, PPI network analysis demonstrated that OsCBL8–OsCIPK17 likely confers disease resistance to seedlings and regulates their growth and development. This was confirmed by the subsequent hub gene analysis.
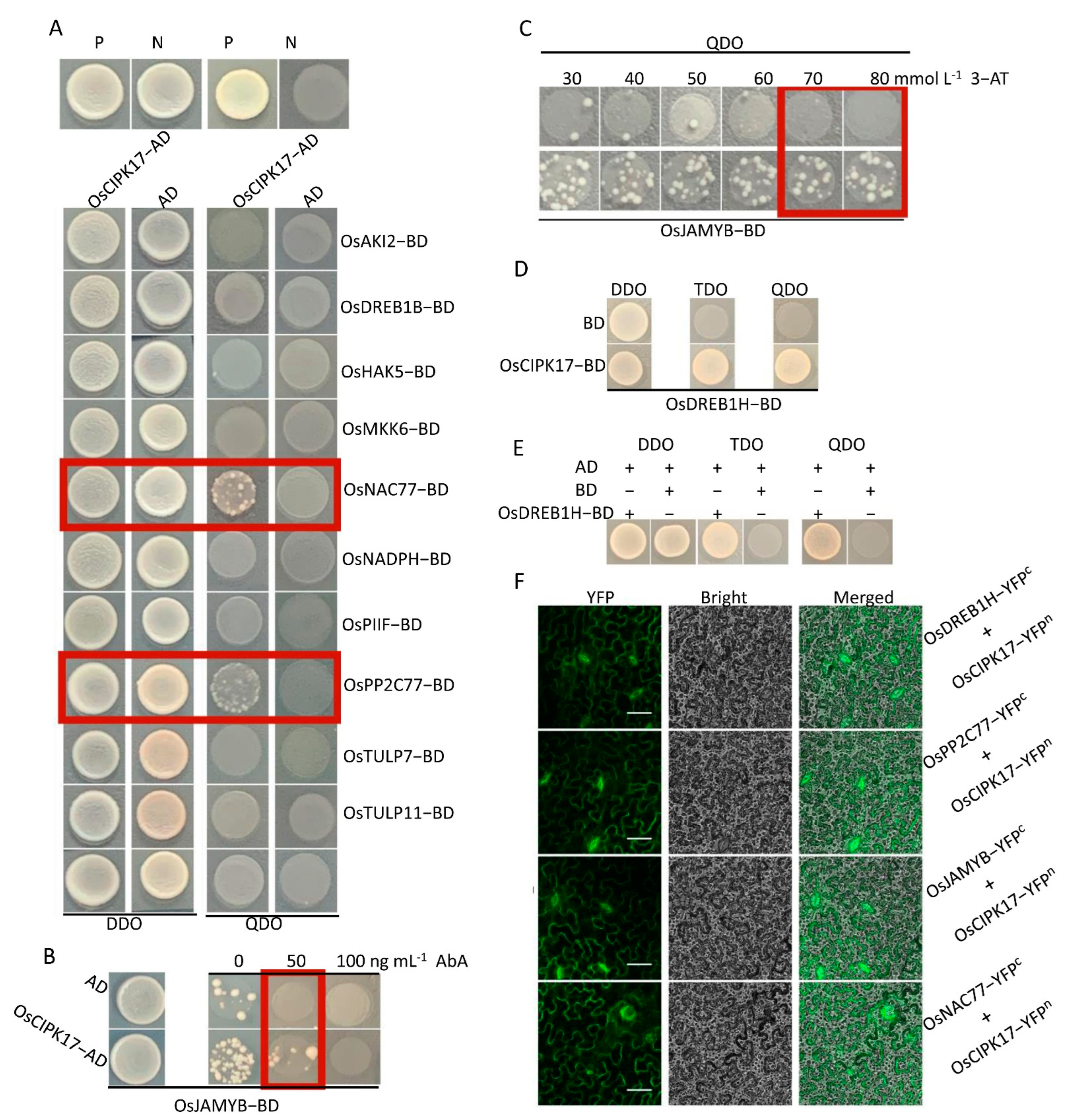
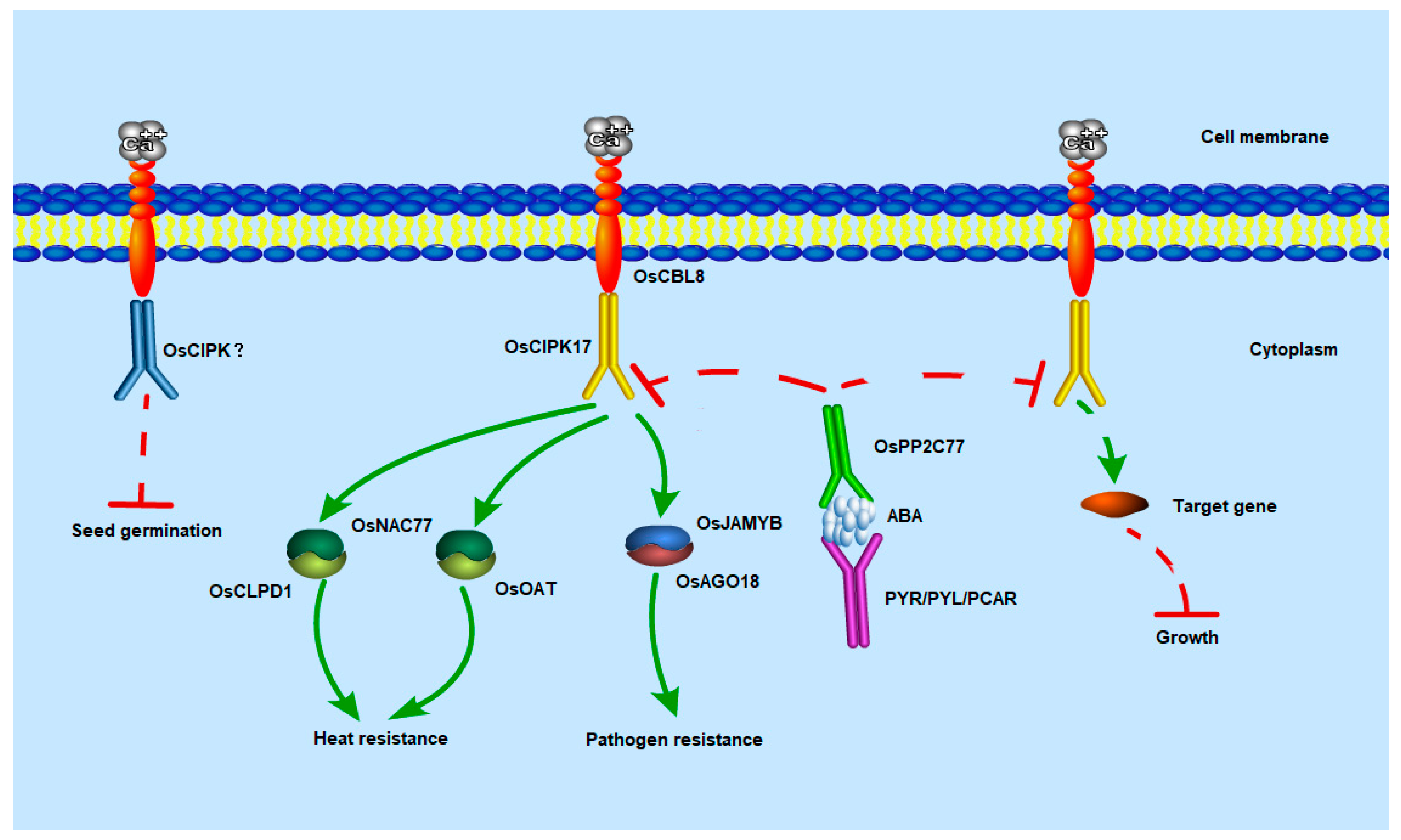
We predicted and verified the target proteins that may have a regulatory relationship with OsCIPK17. The results showed that OsCIPK17 interacts with OsPP2C77, OsJAMYB, and OsNAC77. OsPP2C77 encodes an ABA-mediated phosphatase that interacts with CIPK [
11]. ABA initially binds to its corresponding receptor protein (PYR/PYL/RCAR). Subsequently, the receptor protein binds to PP2C and mediates the PP2C dephosphorylation of CIPK, thereby regulating downstream target proteins [
12]. Similar phenomena have been reported previously. For example, ABI1, ABI2, and AIP1, members of the PP2C family, can interact with CIPK to regulate downstream proteins [
11,
12].
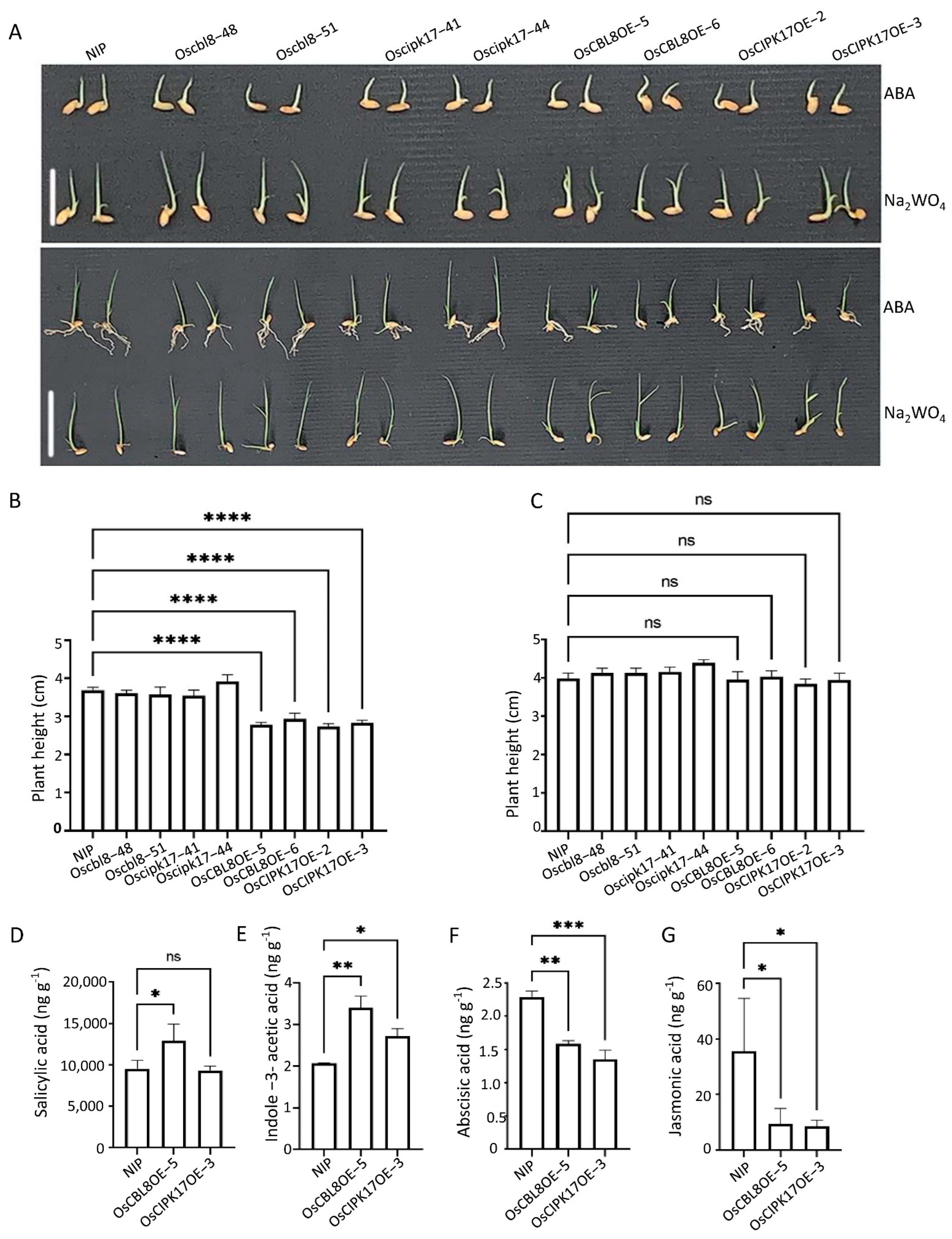
This interaction has only been reported in the model plant
Arabidopsis thaliana. When the seedlings were treated with a low concentration of ABA, the overexpression line maintained the inhibition of growth compared with that in NIP. However, this inhibitory state was restored after treatment with the ABA inhibitor Na
2WO
4. This indicates that ABA is involved in the regulation of seedling growth by OsCBL8–OsCIPK17. There have been many reports on the involvement of ABA in the CBL–CIPK signaling pathway. In apples, CIPK22 interacts with the transcription factor AREB2 to increase sensitivity to ABA through ABA-dependent signal transduction [
46]. However, in our study, the growth of seedlings after ABA or Na
2WO
4 treatment was inhibited compared with that under normal conditions. This is somewhat similar to the effect of auxin; that is, excess or inadequate levels of auxin will inhibit growth and development. The association of auxin with the CBL–CIPK module has been reported. In tobacco,
CaCIPK6 participates in auxin transport [
44]. In addition, in PPI network analysis, node GH3.2 affects auxin metabolism. These results suggest that OsPP2C77, in response to ABA, is involved in OsCBL8–OsCIPK17-mediated regulation of seedling growth and development, which may be achieved through the auxin pathway. OsNAC77 is a plant-specific transcription factor of the NAC family that binds to the DNA-specific sequences of the CLPD1 and OAT promoters in vitro [
47]. CLPD1 encodes a molecular chaperone that plays a role in heat stress responses and helps plants resist dehydration and salt stress [
48]. In addition, it has been reported that overexpression of
CLPD1 in plants leads to increased tolerance to drought and salt stress [
48].
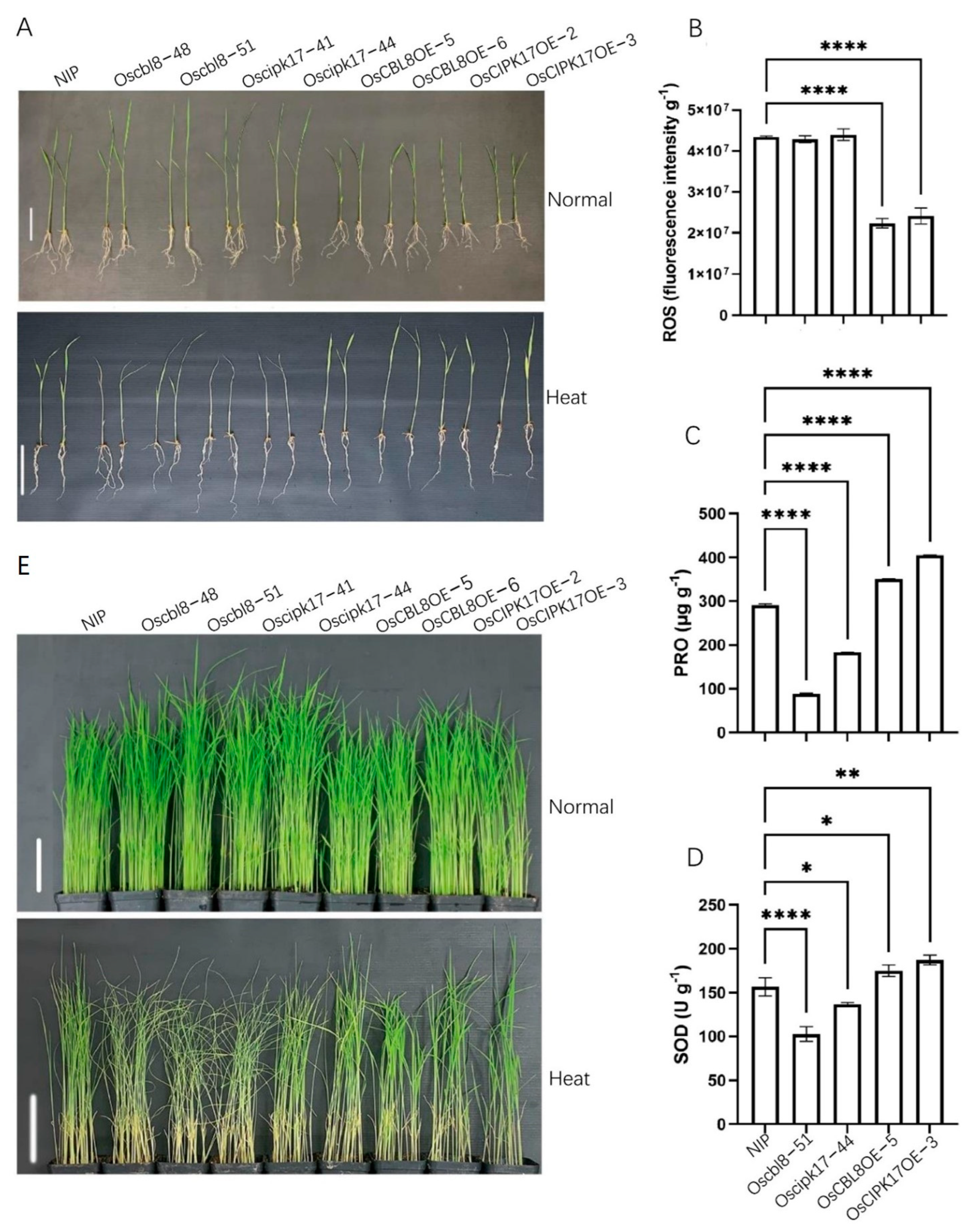
OAT encodes an ornithine aminotransferase that enhances tolerance to drought and oxidative stress, primarily by enhancing ROS clearance and PRO pre-accumulation [
49]. This suggests that OsCBL8 mediates OsCIPK17 phosphorylation of OsNAC77 to regulate seedling resistance to dehydration and salt stress. However, when rice plants are exposed to extreme temperatures and drought, they often become dehydrated. Therefore, OsCBL8–OsCIPK17–OsNAC77 likely regulates seedling responses to temperature and drought. This finding was also confirmed in the present study. Previous studies have also shown that
OsCBL8 responds to a variety of abiotic stresses, including drought [
50]. Fifteen of the thirty-three
CIPK genes (
CIPK1,
CIPK2,
CIPK5,
CIPK9,
CIPK11,
CIPK12,
CIPK15,
CIPK17,
CIPK20,
CIPK21,
CIPK22,
CIPK23,
CIPK24,
CIPK29 and
CIPK30) in rice are responsive to drought [
7]. Further research found that overexpression of
OsCIPK12 and
OsCIPK23 can enhance drought tolerance of rice [
7,
51]. This study also confirmed that overexpression of
CIPK17 could increase drought tolerance of rice (
Figure 8). Members of the NAC family have been widely reported to be involved in plant responses to temperature [
52]. In addition, studies have shown that ABA can also participate in the regulation of NAC in plants. For example, the expression of SNAC3 is induced by drought, high salt, high temperature, ABA, and oxidative stress. It mainly improves the tolerance of transgenic plants to drought and high temperatures by scavenging ROS accumulated under stress in rice [
52]. In addition, some studies have found that NAC family members regulate auxin [
53]. These results indicate that OsNAC77 is likely involved in the ABA-mediated regulatory role of OSCBL8–OSCIPK17 in seedling growth and development. OsJAMYB encodes a JA-mediated transcription factor that directly binds to the AGO18 promoter to activate AGO18 transcription, thereby endowing rice with blast resistance [
43]. In addition, overexpression of
JAMYB during seed germination, seedling growth, and root elongation can increase the salt tolerance of transgenic lines [
54]. These results indicate that OsCBL8–OsCIPK17 likely mediates OsJAMYB to regulate disease resistance and confers tolerance to salt stress in rice. Generally, OsCBL8–OsCIPK17 regulates seedling growth and development through ABA-mediated pathways and endows seedlings with multiple resistances (
Figure 9).
4. Materials and Methods
Identification of gene family and construction of phylogenetic trees and Synteny analyses.
Data for identifying the gene family, construction of phylogenetic trees and synteny analyses of CBL and CIPK, with explicit annotations, were obtained from NCBI and UniProt. Similar data from the two websites were selected for subsequent analyses and experiments. MUSCLE v.3.8.31 was used to generate multiple protein sequence alignments. The aligned sequences were trimmed using TrimAL v.1.2. Phylogenetic trees were reconstructed using maximum-likelihood (ML) in IQ-TREE v.2.0 with 1000 bootstrap replicates. The optimal models of CBL and CIPK were selected automatically in IQ-TREE v.2.0 according to the Bayesian Information Criterion, set to JTT + G4 and JTT + R5, respectively. For the Bayesian inference tree, the configuration file was first created with PhyloSuite v.1.2.2 and then imported into the online website CIPRES; MrBayes v.3.2.7a was used to construct the tree. The phylogenetic trees were edited and annotated using FigTree v.1.4.3 and iTOL v.3. TimeTree was used to construct phylogenetic trees with different divergence times. A phylogenetic tree of the CBL gene family in rice was constructed using PhyML v.3.0, with default settings and 1000 bootstrap replicates.
The syntenic gene pairs were extracted (e-value < 1 × 10−3) within and between species using MCScanX, which selected CBL and CIPK gene family members, and were plotted using TBtools v.1.077. For each pair of duplicated genes, the value of nonsynonymous substitution (Ka)/synonymous substitution (Ks) was calculated using the plug-in Simple Ka/Ks calculator (NG) in TBtools v.1.077, and the frequency distribution histogram was generated using SPSS v.25. The Sankey diagram of replication types in the rice CBL and CIPK gene families was plotted using OmicStudio. Genomic data were obtained from Phytozome and Ensembl Plants.
4.1. Generation and Identification of Transgenic Plants
We constructed the overexpression vectors pWM101 containing the full-length coding sequences of OsCBL2, OsCBL8, and OsCIPK17. The mutant vectors for CRISPR-Cas9 were constructed by Biotechnology Co., Ltd. (Aidijing, Wuhan, China). Resistant transgenic rice plantlets, including overexpression lines and mutants, were obtained from Biotechnology Co., Ltd. The embryogenic callus of rice was infected using the
Agrobacterium-mediated method to generate transgenic rice. Resistant transgenic rice plantlets were identified using PCR. The DNA of the T1 generation mutant lines was extracted according to the instructions provided in the DNA kit (Tiangen, Beijing, China). Primers were designed upstream and downstream of the target nucleotides to be mutated, and DNA sequencing was performed to ensure that homozygous mutant lines were obtained. The overexpression lines were identified using qPCR. The total RNA from T1 generation plants was extracted using TRIzol™ Reagent (Tiangen, Beijing, China). The RNA was converted to complementary DNA (cDNA) using a kit according to the manufacturer’s protocol. qPCR was performed using a 7500 Real-Time PCR Detection System in conjunction with the reaction Mix (Roche, Basel, Switzerland).
OsACTIN-1 (
LOC_Os05g36290) was used as the reference gene. As shown in
Supplementary Figure S20A, the mutant lines lost or gained a few nucleotides, whereas the gene expression of the overexpressed lines was significantly higher than that of Nipponbare (NIP) (
Supplementary Figure S20B). All relevant experiments used the T2 generation plants.
Supplementary Table S1 lists all primers used in this study.
4.2. Growth of Plant Materials, Gene Expression, and RNA-Seq Analysis
The growth and stress treatments of NIP were performed according to the procedure published by Lu et al. [
55] with minor modifications. Seedlings were cultivated hydroponically with Mucun B nutrient solution, in a climate chamber (QY-14; Quanyou Electronic Technology Co., Ltd., Nanjing, China) at 22 °C/20 °C under a 16 h/8 h light/dark cycle at a light intensity of 100 μmol m
−2 s
−1. The seedlings were stressed when they reached the two real leaf stage (22 days). Stress treatments included cold or heat treatment at 8 or 39 °C, respectively, and drought treatment with 20% PEG6000, 100 mmol L
−1 sodium chloride, and 100 μmol L
−1 cadmium sulfate. The shoots and roots of plants subjected to all treatments were sampled, frozen in liquid nitrogen, and stored at −80 °C for gene expression analysis. The seedlings used for measuring plant height, root length, fresh weight, and dry weight were cultured for 2 weeks. Germination rate was calculated after culturing for 1 week. Transcriptome sequencing was performed after culturing for 2 weeks. The samples were sent to Yongji Biotechnology Co., Ltd. (Guangzhou, China) and sequenced using Illumina NovaSeq 6000. Three biological replicates of each line were sequenced. The original RNA-seq data were submitted to the National Genomics Data Center (
https://ngdc.cncb.ac.cn/); access number is PRJCA009668 and this BioProject will be available on 19th May 2024. Seedlings were cultured in the same manner for experiments involving abscisic acid (ABA), Na
2WO
4 (an inhibitor of ABA synthesis), and temperature responses. The high-temperature treatment was performed at 39 °C/30 °C instead of 22 °C/20 °C.
Gene expression was analyzed using RT-qPCR as described in the previous section. The relative expression levels were calculated according to Cao et al. [
56] and visualized as a heat map using TBtools v.1.077. In addition, heat map data were obtained from BAR.
4.3. Subcellular Localization Analysis
Tobacco seeds were surface-sterilized with 70% ethanol and 20% bleach. They were cultured for 4 weeks in a climate chamber. The coding sequences (CDS) of OsCBL2, OsCBL8, and OsCIPK17 were inserted into the enhanced green fluorescent protein (eGFP, pCAMBIA-2300-35S-N-eGFP-OCS) vector to generate fusion constructs, which were then transferred into Agrobacterium (GV3101) and co-cultured to infect tobacco. The infected tobacco was observed after dark culture for 3 days. All images were captured using a confocal laser microscope of TCS SP8 (Leica, Heidelberg, Germany).
4.4. Production of Transgenic Arabidopsis thaliana and GUS Tissue Staining
Transgenic
Arabidopsis thaliana with promoters of the
OsCBL8, maintained in this study, was produced following the methods described above [
26]. β-Glucuronidase (GUS) staining was performed in tissues of T2 transgenic
Arabidopsis thaliana containing the GUS reporter gene using a kit (Tiangen, Beijing, China).
4.5. Yeast Two-Hybrid and Bimolecular Fluorescent Complementation (BiFC) Assays
Yeast two-hybrid (Y2H) assays were performed using the Matchmaker Gold Yeast Two-Hybrid System (Clontech, Kusatsu, Japan), according to the manufacturer’s instructions. Full-length coding sequences of OsCBL were cloned into the pGADT7 vector, and those of OsCIPK were introduced into the pGBKT7 vectors. The plasmids were co-transformed into the Y2HGold strain using the lithium acetate method, according to the TRANFOR protocol. The transformed yeast cells were grown on DDO (SD-Leu/-Trp) medium for 3–5 days. Positive colonies were picked up and grown on TDO (SD-Leu/-Trp/-His) and QDO (SD-Leu/-Trp/-His/-Ade) media for another 3–5 days.
Full-length coding regions of
OsCBL and
OsCIPK were cloned into vectors pSPYCE or pSPYNE [
57]. BiFC assays were performed according to the previously described method [
58]. The tested construct pairs were expressed in the leaves of
Nicotiana benthamiana for three days. YFP fluorescence in the transformed leaves was analyzed using a confocal laser microscope (TCS SP8, Leica, Heidelberg, Germany).
4.6. Analysis of Physiological Parameters
Seed vigor was measured using 2, 3, 5-triphenyltetrazolium chloride (TTC) staining. Shelled seeds were placed in a 2% TTC staining solution and placed in a 33 °C water bath for 2 h for observation. Reactive oxygen species (ROS) content was detected using the fluorescent probe H2DCFDA according to the manufacturer’s instructions. Analysis of proline and superoxide dismutase was performed using a kit (Jiancheng, Nanjing, China) according to the manufacturer’s instructions and measured using a microplate detector (EnSpireTM 2300, PerkinElmer, Waltham, MA, USA) and ultraviolet spectrophotometer (Evolution 300 BB, ThermoFisher, Waltham, MA, USA).
4.7. Measurement of Hormone Content
Leaves of the 2-week-old seedlings of the NIP and overexpression lines were selected for hormone detection. The samples were stored in dry ice and forwarded to PANOMIX (Suzhou, China) for hormone detection using liquid chromatography–mass spectrometry. Three biological replicates were detected for each line.
4.8. Enrichment Analysis of Differential Genes
After obtaining the original transcriptome sequencing data, we used FastQC and Trimmomatic for quality monitoring and control to ensure that they could be used for subsequent analyses. Kallisto was used to obtain the gene expression matrix. The transcription level matrix obtained by Kallisto was transformed into a gene-level matrix using Trans Value Sum. Differentially expressed genes (DEGs) were detected using DESeq2. PlantGSEA was used for enrichment analysis, and the results were visualized using OmicStudio. Fisher’s exact test was used for statistical analysis and the Benjamini–Yekutieli procedure (FDR under dependency) for multi-test adjustment. Gene set enrichment analysis was performed using GSEA v.4.1.0, with default settings. Volcanic maps and other data were visualized in the same manner.
4.9. Construction of the Protein–Protein Interaction (PPI) Network
The acquired DEGs were introduced into STRING to determine interactions. Genetic interactions were introduced into Cytoscape v.3.8.2 to construct the network. The plug-in MCODE with default settings was used to obtain the sub-modules. The expression levels of related genes were introduced into OmicStudio, and correlation analyses of genes in the sub-modules were carried out. Finally, the cytoHubba plug-in with default settings was used to identify hub genes in the network. The results were displayed in the form of a heat map using TBtools v.1.077.
4.10. Statistical Analysis
Figure legends describe in detail the experiments, statistical analyses, and the number of samples. Data analysis and visualization were performed using GraphPad Prism v.9.0.0.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}








