
Application of Exogenous Silicon for Alleviating Photosynthetic Inhibition in Tomato Seedlings under Low−Calcium Stress

Abstract
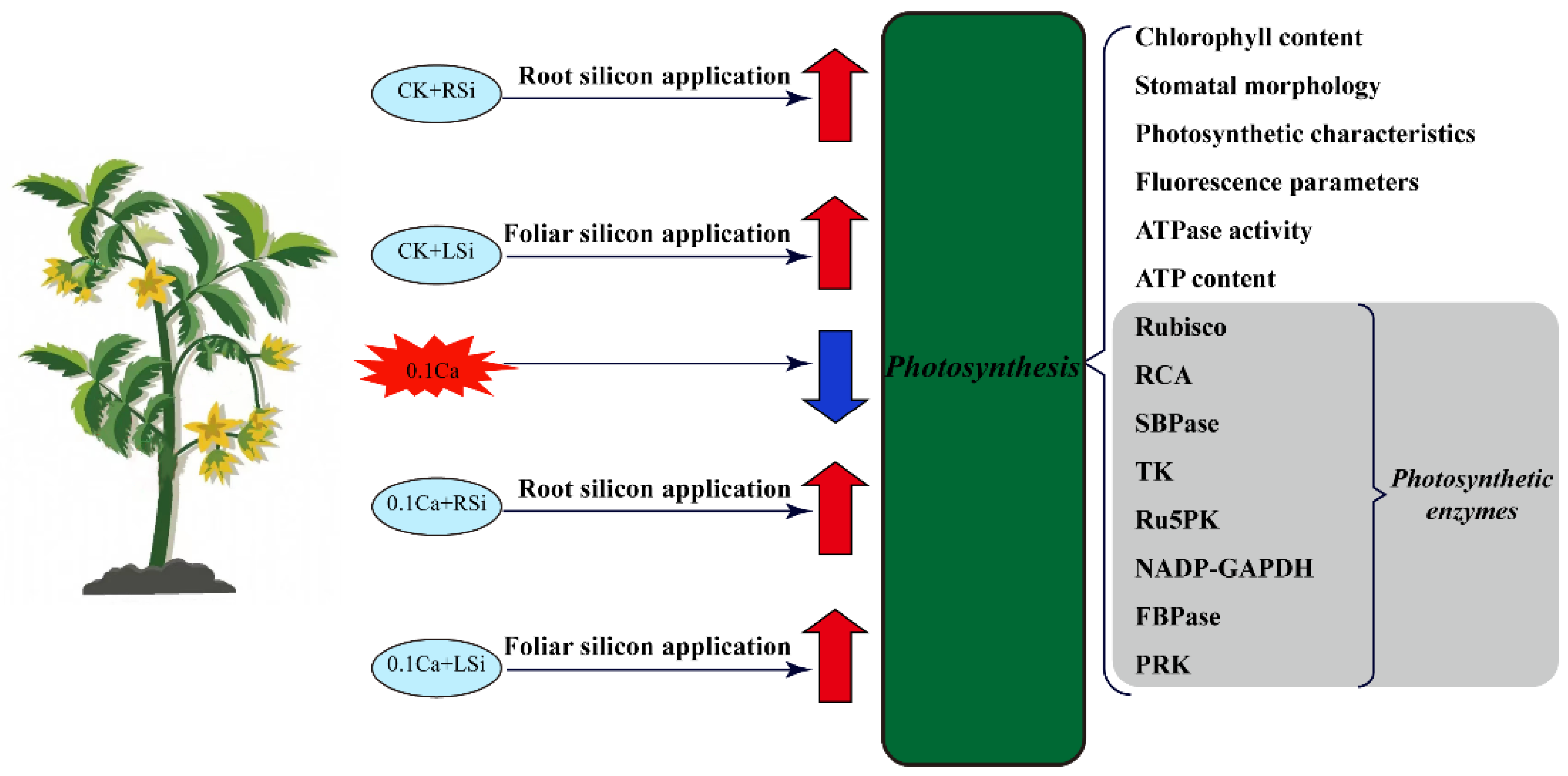
:1. Introduction
2. Results
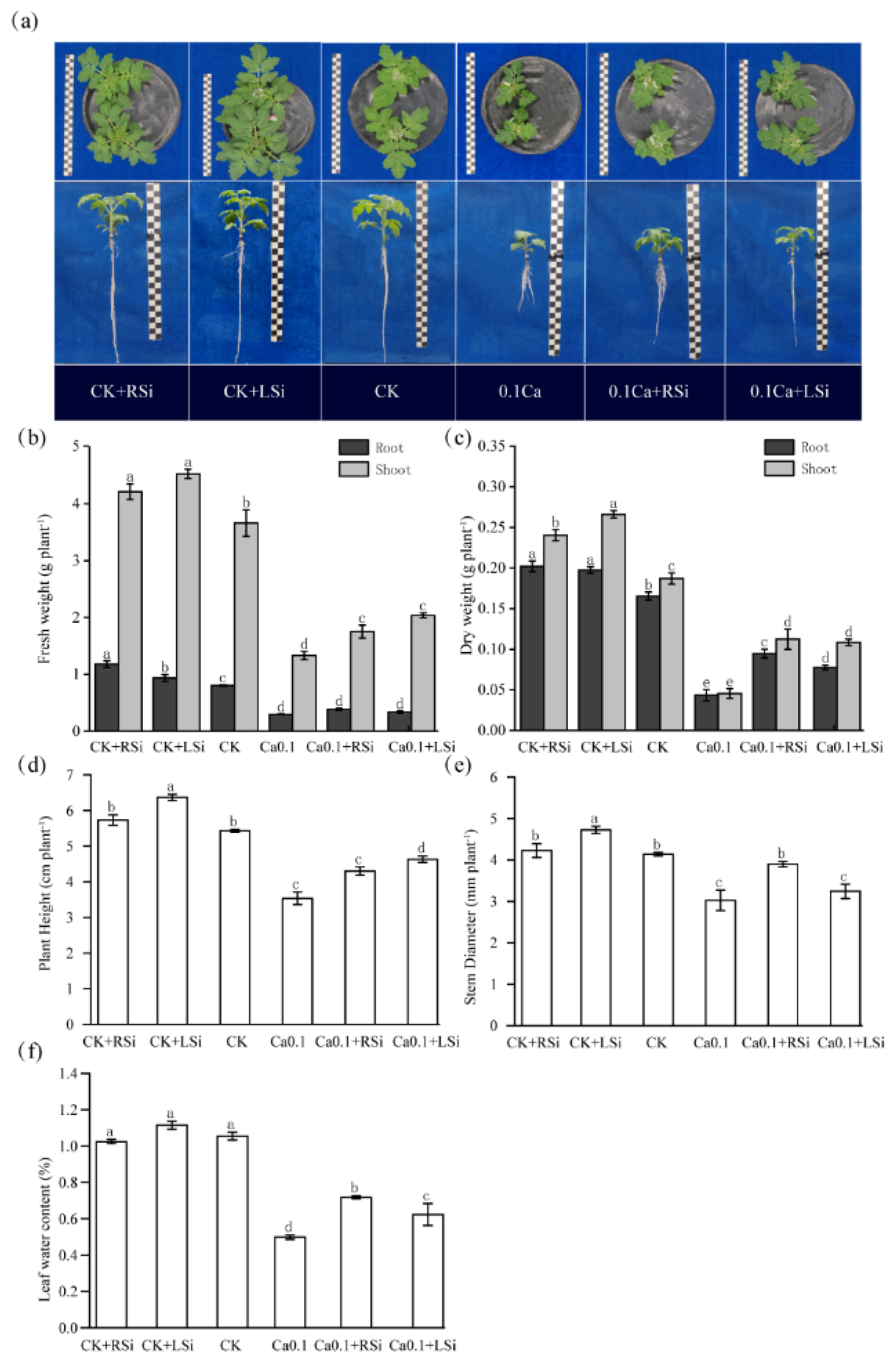
2.1. Effect of Exogenous Si on Growth Parameters in Tomato under Low−Ca Stress
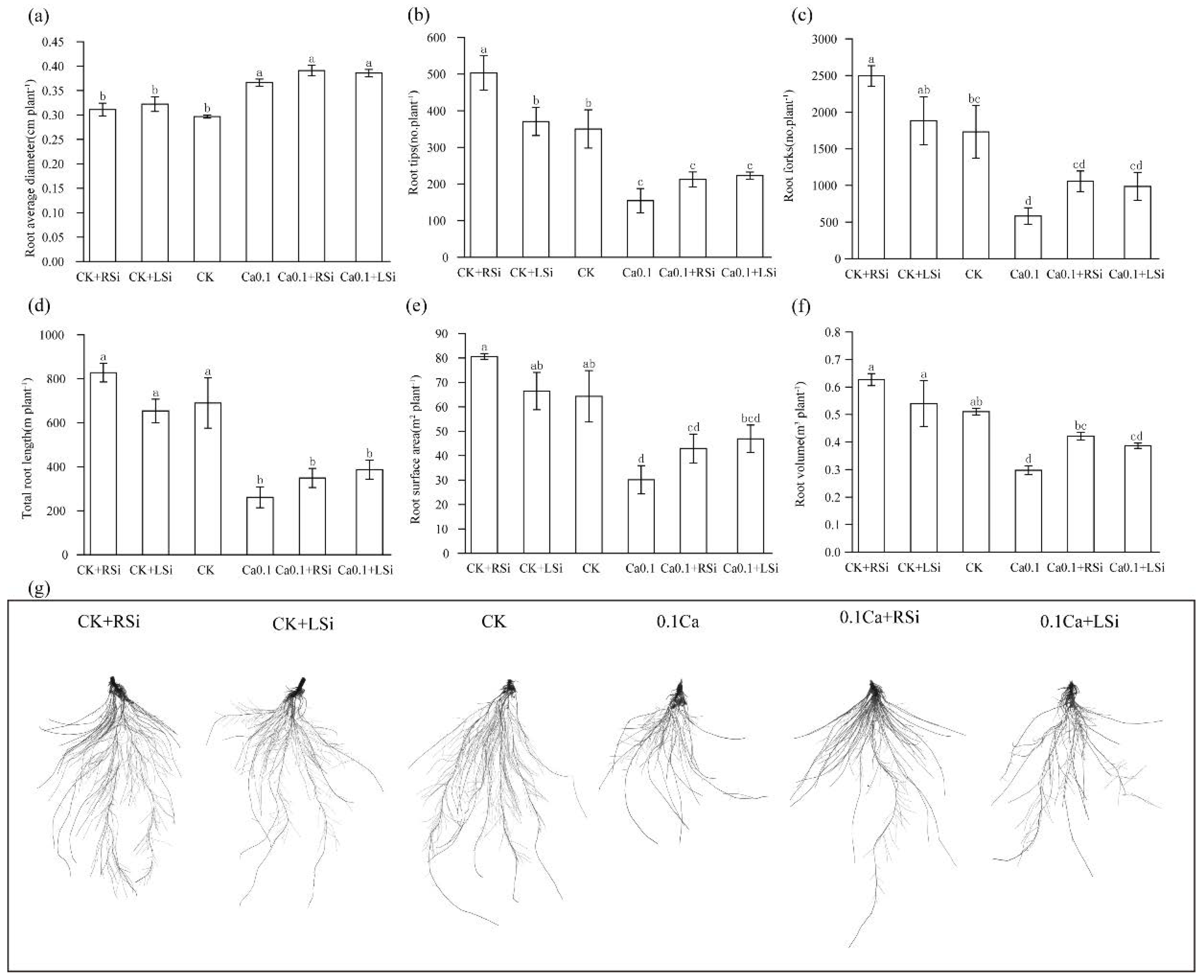
2.2. Effect of Exogenous Si on Tomato Root System under Low−Ca Stress
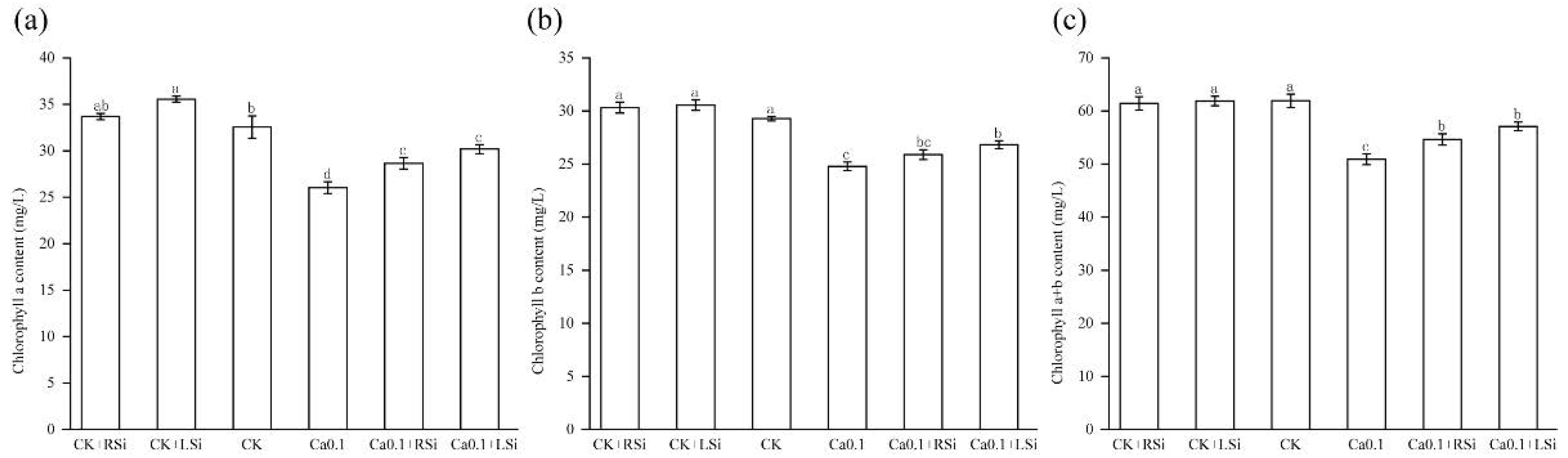
2.3. Effect of Exogenous Si on Chlorophyll Content of Tomato Leaves under Low−Ca Stress
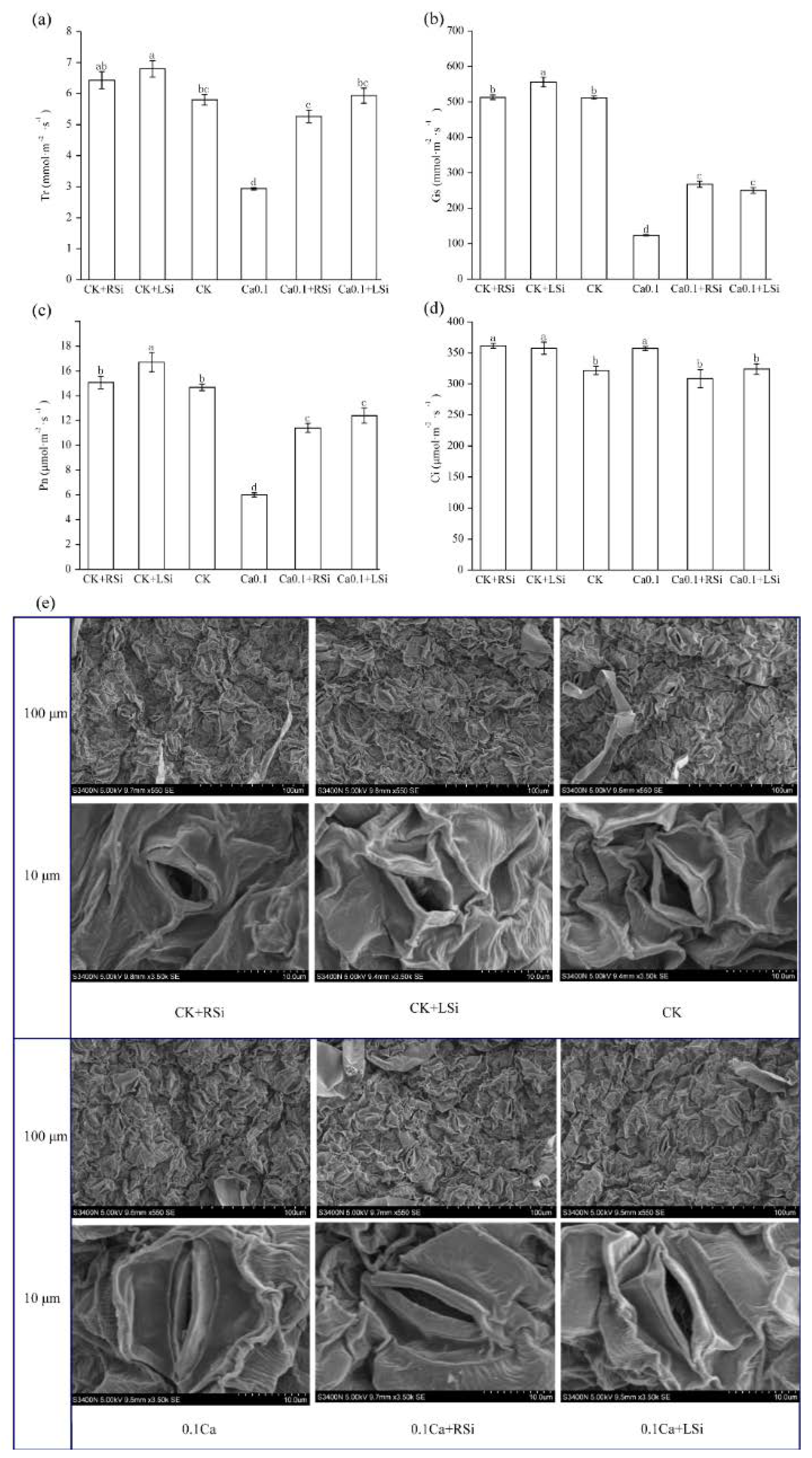
2.4. Effect of Exogenous Si on Stomatal Morphology and Photosynthetic Characteristics of Tomato Leaves under Low−Ca Stress
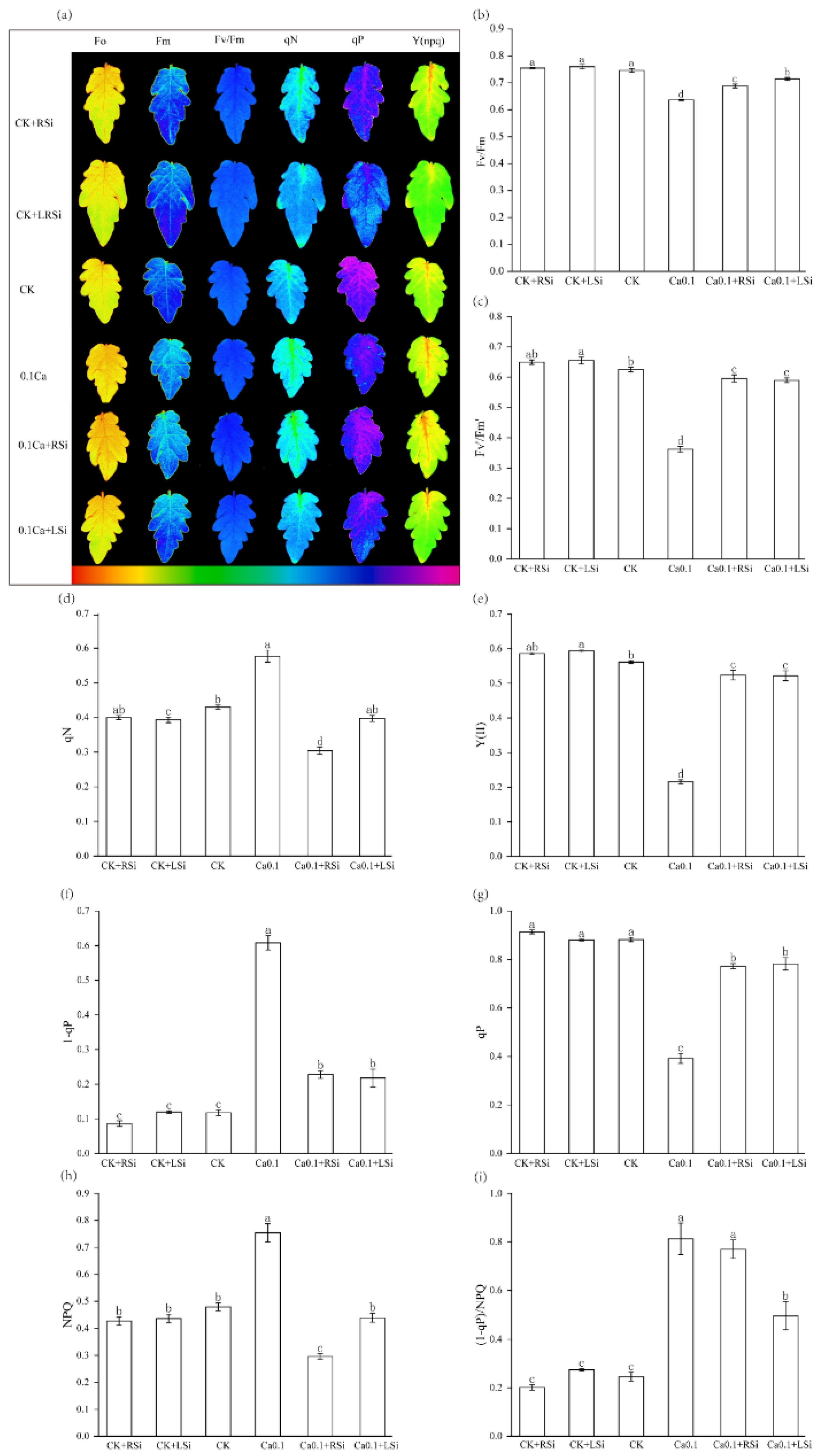
2.5. Effect of Exogenous Si on Fluorescence Parameters of Tomato Leaves under Low−Ca Stress
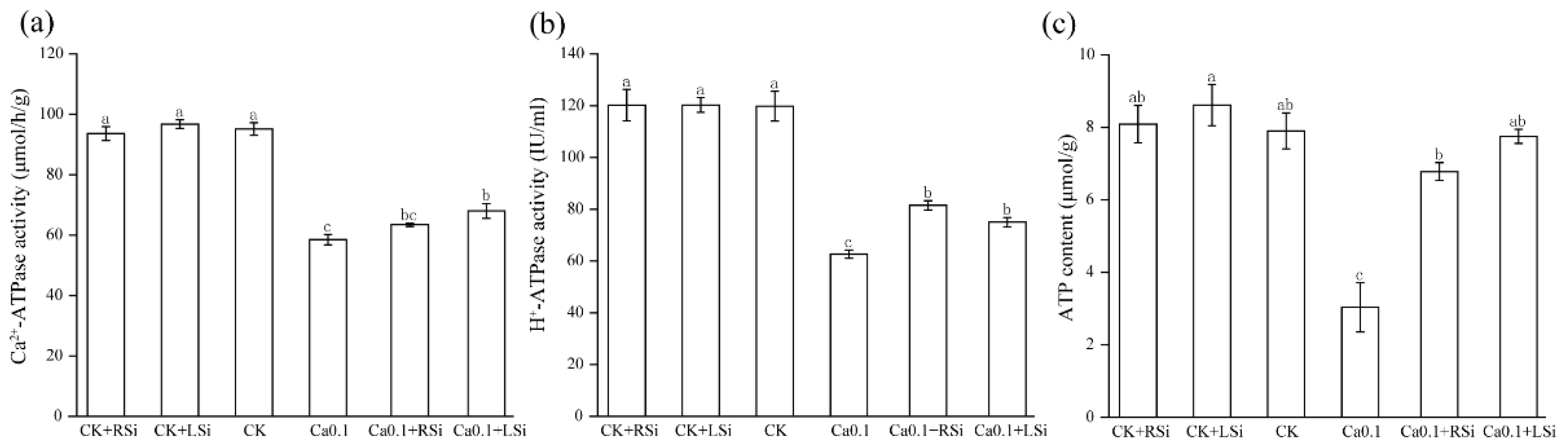
2.6. Effect of Exogenous Si on ATPase Activity and ATP Content of Tomato Chloroplasts under Low−Ca Stress
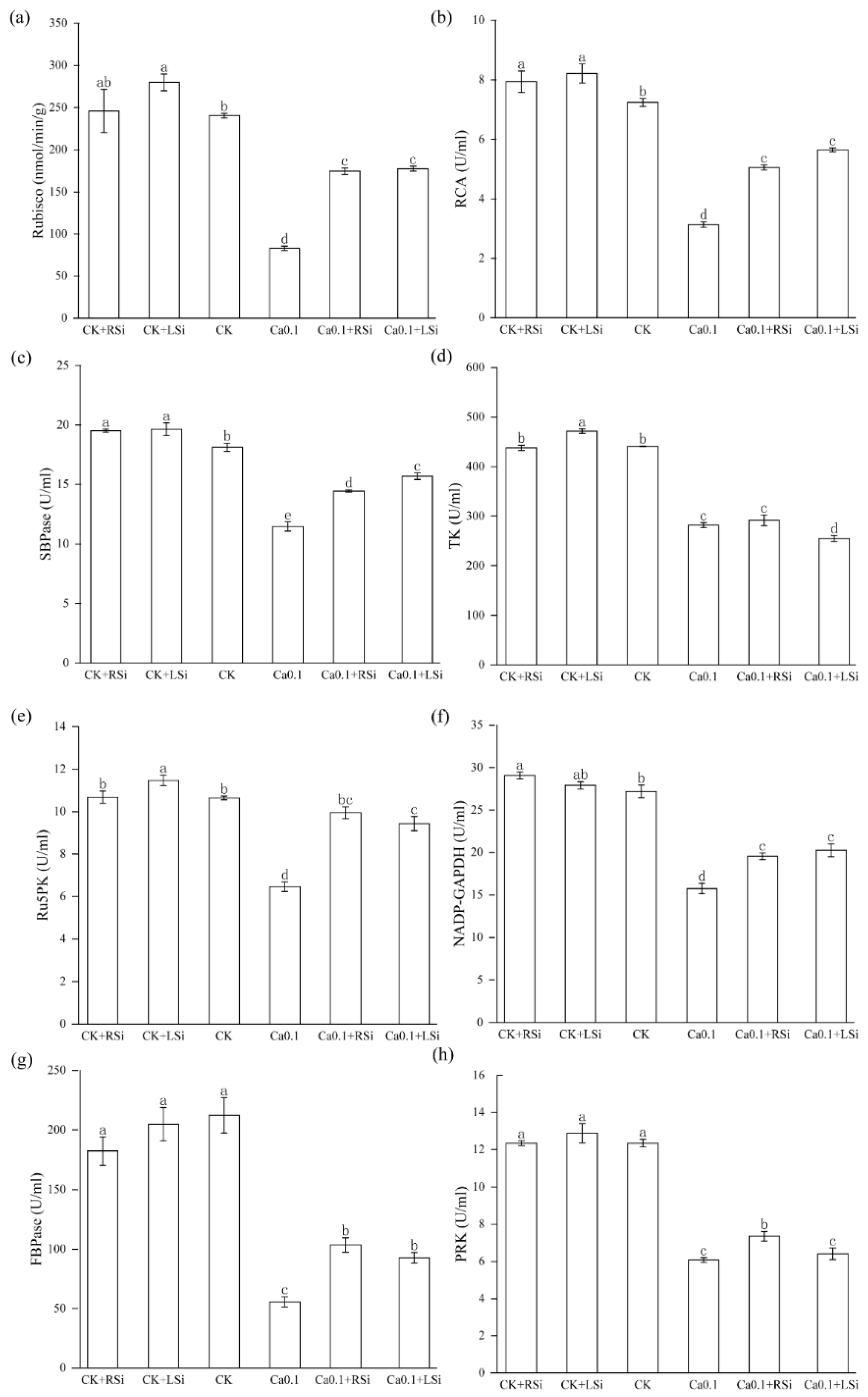
2.7. Effect of Exogenous Si on Photosynthesis−Related Enzymatic Activities in Tomato under Low−Ca Stress
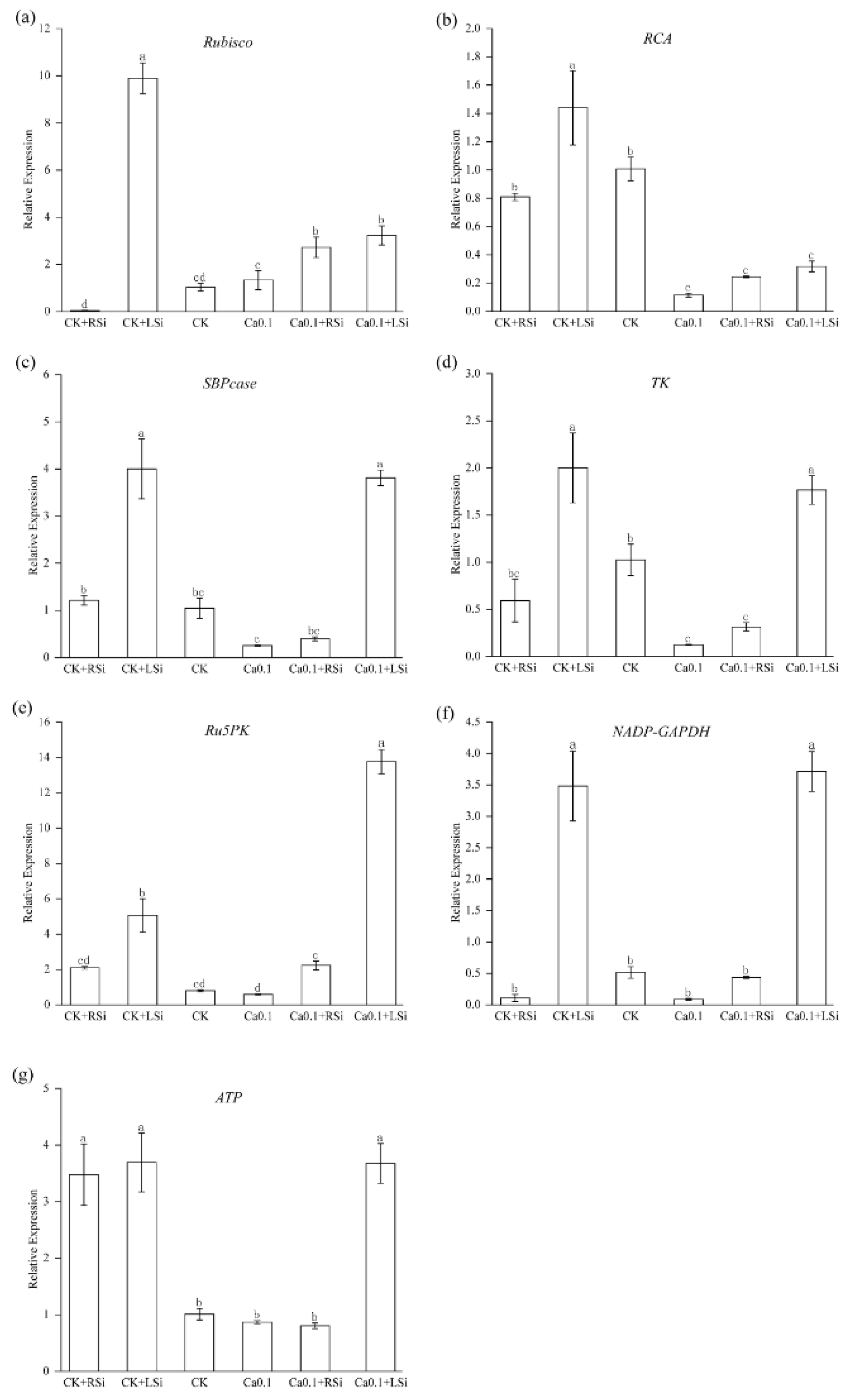
2.8. Effect of Exogenous Si on the Expression of Genes Related to Photosynthetic Enzymatic Activity in Tomato under Low−Ca Stress
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growing Conditions
4.2. Treatment and Experimental Design
4.3. Growth Indices
4.4. Leaf Relative Water Content
4.5. Root System Scanning
4.6. Chlorophyll Content
4.7. Determination of Photosynthetic Parameters
4.8. Chlorophyll Fluorescence Parameters
4.9. Leaf Stomatal Morphology Observation
4.10. Determination of Chloroplast ATPase Activity and Content
4.11. Determination of Photosynthetic Enzymatic Activity
4.12. Determination of Relative Gene Expression
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sommer, M.; Kaczorek, D.; Kuzyakov, Y.; Breuer, J. Silicon Pools and Fluxes in Soils and Landscapes—A Review. J. Plant Nutr. Soil Sci. 2006, 169, 310–329. [Google Scholar] [CrossRef]
- Collin, B.; Doelsch, E.; Keller, C.; Cazevieille, P.; Tella, M.; Chaurand, P.; Panfili, F.; Hazemann, J.L.; Meunier, J.D. Evidence of Sulfur-bound Reduced Copper in Bamboo Exposed to High Silicon and Copper Concentrations. Environ. Pollut. 2014, 187, 22–30. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.K.; Singh, S.; Singh, V.P.; Prasad, S.M.; Dubey, N.K.; Chauhan, D.K. Silicon Nanoparticles more Effectively Alleviated UV-B Stress than Silicon in Wheat (Triticum aestivum) seedlings. Plant Physiol. Biochem. 2017, 110, 70–81. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.R.; Ashfaque, F.; Chhillar, H.; Irfan, M.; Khan, N.A. The Intricacy of Silicon, Plant Growth Regulators and Other Signaling Molecules for Abiotic Stress Tolerance: An Entrancing Crosstalk between Stress Alleviators. Plant Physiol. Biochem. 2021, 162, 36–47. [Google Scholar] [CrossRef] [PubMed]
- Prychid, C.J.; Rudall, P.J.; Gregory, M. Systematics and Biology of Silica Bodies in Monocotyledons. Bot. Rev. 2003, 69, 377–440. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. Silicon Uptake and Accumulation in Higher Plants. Trends Plant Sci. 2006, 11, 392–397. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N. A Cooperative System of Silicon Transport in Plants. Trends Plant Sci. 2015, 20, 435–442. [Google Scholar] [CrossRef]
- Huang, C.-H.; Roberts, P.D.; Datnoff, L.E. Silicon Suppresses Fusarium Crown and Root Rot of Tomato: Silicon Suppresses Fusarium Crown and Root Rot of Tomato. J. Phytopathol. 2011, 159, 546–554. [Google Scholar] [CrossRef]
- Gong, H.; Chen, K. The Regulatory Role of Silicon on Water Relations, Photosynthetic Gas Exchange, and Carboxylation Activities of Wheat Leaves in Field Drought Conditions. Acta Physiol. Plant. 2012, 34, 1589–1594. [Google Scholar] [CrossRef]
- Exley, C. A Possible Mechanism of Biological Silicification in Plants. Front. Plant Sci. 2015, 6, 853. [Google Scholar] [CrossRef]
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of Photosynthesis during Abiotic Stress-Induced Photoinhibition. Mol. Plant 2015, 8, 1304–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debona, D.; Rodrigues, F.A.; Datnoff, L.E. Silicon’s Role in Abiotic and Biotic Plant Stresses. Annu. Rev. Phytopathol. 2017, 55, 85–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.-H.; Khan, A.L.; Waqas, M.; Lee, I.-J. Silicon Regulates Antioxidant Activities of Crop Plants under Abiotic-Induced Oxidative Stress: A Review. Front. Plant Sci. 2017, 8, 510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etesami, H.; Jeong, B.R. Silicon (Si): Review and Future Prospects on the Action Mechanisms in Alleviating Biotic and Abiotic Stresses in Plants. Ecotoxicol. Environ. Saf. 2018, 147, 881–896. [Google Scholar] [CrossRef]
- Li, Z.; Song, Z.; Yang, X.; Song, A.; Yu, C.; Wang, T.; Xia, S.; Liang, Y. Impacts of Silicon on Biogeochemical Cycles of Carbon and Nutrients in Croplands. J. Integr. Agric. 2018, 17, 2182–2195. [Google Scholar] [CrossRef]
- Yan, G.; Nikolic, M.; Ye, M.; Xiao, Z.; Liang, Y. Silicon Acquisition and Accumulation in Plant and Its Significance for Agriculture. J. Integr. Agric. 2018, 17, 2138–2150. [Google Scholar] [CrossRef]
- Hassan, H.; Alatawi, A.; Abdulmajeed, A.; Emam, M.; Khattab, H. Roles of Si and SiNPs in Improving Thermotolerance of Wheat Photosynthetic Machinery via Upregulation of PsbH, PsbB and PsbD Genes Encoding PSII Core Proteins. Horticulturae 2021, 7, 16. [Google Scholar] [CrossRef]
- Song, A.; Li, P.; Fan, F.; Li, Z.; Liang, Y. The Effect of Silicon on Photosynthesis and Expression of Its Relevant Genes in Rice (Oryza sativa L.) under High-Zinc Stress. PLoS ONE 2014, 9, e113782. [Google Scholar] [CrossRef]
- Hussain, S.; Shuxian, L.; Mumtaz, M.; Shafiq, I.; Iqbal, N.; Brestic, M.; Shoaib, M.; Sisi, Q.; Li, W.; Mei, X.; et al. Foliar Application of Silicon Improves Stem Strength Under Low Light Stress by Regulating Lignin Biosynthesis Genes in Soybean (Glycine max (L.) Merr.). J. Hazard. Mater. 2021, 401, 123256. [Google Scholar] [CrossRef]
- Chaiwong, N.; Prom-u-thai, C.; Bouain, N.; Lacombe, B.; Rouached, H. Individual versus Combinatorial Effects of Silicon, Phosphate, and Iron Deficiency on the Growth of Lowland and Upland Rice Varieties. Int. J. Mol. Sci. 2018, 19, 899. [Google Scholar] [CrossRef]
- Wang, H.; Li, C.; Liang, Y. Chapter 21 Agricultural Utilization of Silicon in China. Stud. Plant Sci. 2001, 8, 343–358. [Google Scholar] [CrossRef]
- de Souza Alonso, T.A.; Ferreira Barreto, R.; de Mello Prado, R.; Pereira de Souza, J.; Falleiros Carvalho, R. Silicon Spraying Alleviates Calcium Deficiency in Tomato Plants, But Ca-EDTA Is Toxic. J. Plant Nutr. Soil Sci. 2020, 183, 659–664. [Google Scholar] [CrossRef]
- Toresano-Sánchez, F.; Valverde-García, A.; Camacho-Ferre, F. Effect of The Application of Silicon Hydroxide on Yield and Quality of Cherry Tomato. J. Plant Nutr. 2012, 35, 567–590. [Google Scholar] [CrossRef]
- Hepler, P.K. Calcium: A Central Regulator of Plant Growth and Development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Aras, S.; Keles, H.; Bozkurt, E. Physiological and Histological Responses of Peach Plants Grafted onto Different Rootstocks Under Calcium Deficiency Conditions. Sci. Hortic. 2021, 281, 109967. [Google Scholar] [CrossRef]
- Liang, W.; Wang, M.; Ai, X. The Role of Calcium in Regulating Photosynthesis and Related Physiological Indexes of Cucumber Seedlings Under Low Light Intensity and Suboptimal Temperature Stress. Sci. Hortic. 2009, 123, 34–38. [Google Scholar] [CrossRef]
- Zhang, K.; Yue, D.; Wei, W.; Hu, Y.; Feng, J.; Zou, Z. Characterization and Functional Analysis of Calmodulin and Calmodulin-Like Genes in Fragaria vesca. Front. Plant Sci. 2016, 7, 1820. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Zeng, F.; Song, W.; Zhang, M.; Deng, R. Effects of Calcium and Lanthanum on ABA Biosynthesis in Cucumber Leaves. Russ. J. Plant Physiol. 2002, 49, 696–699. [Google Scholar] [CrossRef]
- Saito, S.; Uozumi, N. Calcium-Regulated Phosphorylation Systems Controlling Uptake and Balance of Plant Nutrients. Front. Plant Sci. 2020, 11, 44. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef]
- Hernández-Pérez, O.I.; Valdez-Aguilar, L.A.; Alia-Tejacal, I.; Cartmill, A.D.; Cartmill, D.L. Tomato Fruit Yield, Quality, and Nutrient Status in Response to Potassium: Calcium Balance and Electrical Conductivity in the Nutrient Solution. J. Soil Sci. Plant Nutr. 2020, 20, 484–492. [Google Scholar] [CrossRef]
- Weinl, S.; Held, K.; Schlücking, K.; Steinhorst, L.; Kuhlgert, S.; Hippler, M.; Kudla, J. A Plastid Protein Crucial for Ca2+-Regulated Stomatal responses. New Phytol. 2008, 179, 675–686. [Google Scholar] [CrossRef]
- Popelkova, H.; Boswell, N.; Yocum, C. Probing the Topography of the Photosystem II Oxygen Evolving Complex: Psbo is Required for Efficient Calcium Protection of the Manganese Cluster Against Dark-Inhibition by an Artificial Reductant. Photosynth. Res. 2011, 110, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Rocha, A.; Vothknecht, U. Identification of CP12 as a Novel Calcium-Binding Protein in Chloroplasts. Plants 2013, 2, 530–540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Białasek, M.; Górecka, M.; Mittler, R.; Karpiński, S. Evidence for the Involvement of Electrical, Calcium and ROS Signaling in the Systemic Regulation of Non-Photochemical Quenching and Photosynthesis. Plant Cell Physiol. 2017, 58, 207–215. [Google Scholar] [CrossRef] [PubMed]
- Kalaji, H.M.; Rastogi, A.; Živčák, M.; Brestic, M.; Daszkowska-Golec, A.; Sitko, K.; Alsharafa, K.Y.; Lotfi, R.; Stypiński, P.; Samborska, I.A.; et al. Prompt Chlorophyll Fluorescence as A Tool for Crop Phenotyping: An Example of Barley Landraces Exposed to Various Abiotic Stress Factors. Photosynthetica 2018, 56, 953–961. [Google Scholar] [CrossRef] [Green Version]
- Prado, R.D.M.; Santos, V.H.G.; Gondim, A.; Alves, A.U.; Cecílio Filho, A.B.; Correia, M.A.R. Crescimento E Marcha de Absorção de Nutrientes em Tomateiro Cultivar RaÍSa Cultivado em Sistema Hidropônico. Semin.-Cienc. Agrar. 2011, 32, 19. [Google Scholar] [CrossRef] [Green Version]
- Bergougnoux, V. The History of Tomato: From Domestication to Biopharming. Biotechnol. Adv. 2014, 32, 170–189. [Google Scholar] [CrossRef]
- Mestre, T.C.; Garcia-Sanchez, F.; Rubio, F.; Martinez, V.; Rivero, R.M. Glutathione Homeostasis as an Important and Novel Factor Controlling Blossom-End Rot Development in Calcium-Deficient Tomato Fruits. J. Plant Physiol. 2012, 169, 1719–1727. [Google Scholar] [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D. Reactive Oxygen Species Produced by NADPH Oxidase Regulate Plant Cell Growth. Nature 2003, 422, 442–446. [Google Scholar] [CrossRef]
- Schmitz-Eiberger, M.; Haefs, R.; Noga, G. Reduction of Calcium Deficiency Symptoms by Exogenous Application of Calcium Chloride Solutions. Acta Hortic. Sin. 2002, 594, 535–540. [Google Scholar] [CrossRef]
- Saure, M.C. Why Calcium Deficiency is not the Cause of Blossom-End Rot in Tomato and Pepper Fruit—A Reappraisal. Scientia Hortic. 2014, 174, 151–154. [Google Scholar] [CrossRef]
- Islam, M.Z.; Mele, M.A.; Baek, J.P.; Kang, H.-M. Cherry Tomato Qualities Affected by Foliar Spraying with Boron and Calcium. Hortic. Environ. Biotechnol. 2016, 57, 46–52. [Google Scholar] [CrossRef]
- White, P.J. Calcium in Plants. Ann. Bot. 2003, 92, 487–511. [Google Scholar] [CrossRef]
- Kleemann, M. Development of Calcium Deficiency Symptoms in Chervil (Antriscus cerefolium (L.) Hoffm.) And Curled Parsley (Petroselinum crispum (Mill.) Nym. Convar Crispum). Zesz. Probl. Postępów Nauk. Rol. 1999, 468, 335–348. [Google Scholar]
- Olle, M.; Bender, I. Causes and Control of Calcium Deficiency Disorders in Vegetables: A Review. J. Hortic. Sci. Biotechnol. 2009, 84, 577–584. [Google Scholar] [CrossRef]
- Detmann, K.C.; Araújo, W.L.; Martins, S.C.V.; Sanglard, L.M.V.P.; Reis, J.V.; Detmann, E.; Rodrigues, F.Á.; Nunes-Nesi, A.; Fernie, A.R.; DaMatta, F.M. Silicon Nutrition Increases Grain Yield, Which, in Turn, Exerts a Feed-Forward Stimulation of Photosynthetic Rates Via Enhanced Mesophyll Conductance and Alters Primary Metabolism in Rice. New Phytol. 2012, 196, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Van Bockhaven, J.; Steppe, K.; Bauweraerts, I.; Kikuchi, S.; Asano, T.; Höfte, M.; De Vleesschauwer, D. Primary Metabolism Plays a Central Role in Moulding Silicon-Inducible Brown Spot Resistance in Rice: Primary Metabolism Moulds Si-Induced Resistance. Mol. Plant Pathol. 2015, 16, 811–824. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial Effects of Silicon in Alleviating Salinity Stress of Tomato Seedlings Grown Under Sand Culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Pavlovic, J.; Samardzic, J.; Kostic, L.; Laursen, K.H.; Natic, M.; Timotijevic, G.; Schjoerring, J.K.; Nikolic, M. Silicon Enhances Leaf Remobilization of Iron in Cucumber Under Limited Iron Conditions. Ann. Bot. 2016, 118, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Hattori, T.; Inanaga, S.; Araki, H.; An, P.; Morita, S.; Luxova, M.; Lux, A. Application of Silicon Enhanced Drought Tolerance in Sorghum Bicolor. Physiol. Plant. 2005, 123, 459–466. [Google Scholar] [CrossRef]
- Epstein, E. Silicon: Its Manifold Roles in Plants. Ann. Appl. Biol. 2009, 155, 155–160. [Google Scholar] [CrossRef]
- Jang, S.-W.; Kim, Y.; Khan, A.L.; Na, C.-I.; Lee, I.-J. Exogenous Short-Term Silicon Application Regulates Macro-Nutrients, Endogenous Phytohormones, and Protein Expression in Oryza Sativa, L. BMC Plant Biol. 2018, 18, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalaji, H.M.; Dąbrowski, P.; Cetner, M.D.; Samborska, I.A.; Łukasik, I.; Brestic, M.; Zivcak, M.; Tomasz, H.; Mojski, J.; Kociel, H.; et al. A Comparison Between Different Chlorophyll Content Meters Under Nutrient Deficiency Conditions. J. Plant Nutr. 2017, 40, 1024–1034. [Google Scholar] [CrossRef]
- Harrison, E.L.; Arce Cubas, L.; Gray, J.E.; Hepworth, C. The Influence of Stomatal Morphology and Distribution on Photosynthetic Gas Exchange. Plant J. 2020, 101, 768–779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Yang, S.; Wan, S.; Li, X. The Significance of Calcium in Photosynthesis. Int. J. Mol. Sci. 2019, 20, 1353. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Yu, L.; Li, B.; Du, N.; Guo, S. The Effect of Exogenous Calcium on Cucumber Fruit Quality, Photosynthesis, Chlorophyll Fluorescence, and Fast Chlorophyll Fluorescence During the Fruiting Period Under Hypoxic Stress. BMC Plant Biol. 2018, 18, 180. [Google Scholar] [CrossRef]
- Zhang, Y.; Shi, Y.; Gong, H.; Zhao, H.; Li, H.; Hu, Y.; Wang, Y. Beneficial Effects of Silicon on Photosynthesis of Tomato Seedlings under Water Stress. J. Integr. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef] [Green Version]
- Sha, Y.; Fang, W.; Feng, G.; Jing-Jing, M.; Xin-Guo, L.; Shu-Ting, D.; Shu-Bo, W. Exogenous Calcium Alleviates Photoinhibition of PSII by Improving the Xanthophyll Cycle in Peanut (Arachis hypogaea) Leaves during Heat Stress under High Irradiance. PLoS ONE 2013, 8, e71214. [Google Scholar] [CrossRef] [Green Version]
- Tyryshkin, A.M.; Watt, R.K.; Baranov, S.V.; Dasgupta, J.; Hendrich, M.P.; Dismukes, G.C. Supporting Information for Spectroscopic Evidence for Ca2+ Involvement in the Assembly of the Mn4Ca Cluster in the Photosynthetic Water Oxidizing Complex. Biochemistry 2006, 45, 12876–12889. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon Alleviates Drought Stress of Rice Plants by Improving Plant Water Status, Photosynthesis and Mineral Nutrient Absorption. Biol. Trace Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Guo, J.; Feng, R.; Jia, J.; Han, W.; Gong, H. The Regulatory Role of Silicon on Carbohydrate Metabolism in Cucumis sativus, L. under Salt Stress. Plant Soil 2016, 406, 231–249. [Google Scholar] [CrossRef]
- Rastogi, A.; Yadav, S.; Hussain, S.; Kataria, S.; Hajihashemi, S.; Kumari, P.; Xinghong, Y.; Brestic, M. Does Silicon Really Matter for the Photosynthetic Machinery in Plants…? Plant Physiol. Biochem. 2021, 169, 40–48. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zeng, H.; Xu, F.; Yan, F.; Xu, W. H+-ATPases in Plant Growth and Stress Responses. Annu. Rev. Plant Biol. 2022, 73, 495–521. [Google Scholar] [CrossRef] [PubMed]
- Sukhov, V.; Surova, L.; Morozova, E.; Sherstneva, O.; Vodeneev, V. Changes in H+-ATP Synthase Activity, Proton Electrochemical Gradient, and pH in Pea Chloroplast Can Be Connected with Variation Potential. Front. Plant Sci. 2016, 07, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Xu, F.; Yuan, W.; Zhang, D.; Liu, J.; Sun, L.; Cui, L.; Zhang, J.; Xu, W. Rice G Protein Γ Subunit Qpe9-1 Modulates Root Elongation for Phosphorus Uptake by Involving 14-3-3 Protein Osgf14b and Plasma Membrane H+-Atpase. Plant J. 2021, 107, 1603–1615. [Google Scholar] [CrossRef]
- Kreimer, G.; Melkonian, M.; Holtum, J.A.M.; Latzko, E. Stromal Free Calcium Concentration and Light-Mediated Activation of Chloroplast Fructose-1,6-Bisphosphatase. Plant Physiol. 1988, 86, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Raines, C.A. The Calvin Cycle Revisited. Photosynth. Res. 2003, 75, 1–10. [Google Scholar] [CrossRef]
- Delobel, A.; Graciet, E.; Andreescu, S.; Gontero, B.; Halgand, F.; Laprévote, O. Mass Spectrometric Analysis of the Interactions Between CP12, A Chloroplast Protein, and Metal Ions: A Possible Regulatory Role Within a PRK/GAPDH/CP12 Complex. Rapid Commun. Mass Spectrom. 2005, 19, 3379–3388. [Google Scholar] [CrossRef] [Green Version]
- Sanglard, L.M.V.P.; Martins, S.C.V.; Detmann, K.C.; Silva, P.E.M.; Lavinsky, A.O.; Silva, M.M.; Detmann, E.; Araújo, W.L.; DaMatta, F.M. Silicon Nutrition Alleviates the Negative Impacts of Arsenic on the Photosynthetic Apparatus of Rice Leaves: An Analysis of the Key Limitations of Photosynthesis. Physiol. Plant. 2014, 152, 355–366. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, T.; Calomme, M.; Borowiak, K. The Effect of Choline-Stabilized Orthosilicic Acid on Microelements and Silicon Concentration, Photosynthesis Activity and Yield of Tomato Grown Under Mn Stress. Plant Physiol. Biochem. 2015, 96, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, B.; Jiang, D.; Chen, G. Silicon Improves Photosynthetic Performance by Optimizing Thylakoid Membrane Protein Components in Rice Under Drought Stress. Environ. Exp. Bot. 2019, 158, 117–124. [Google Scholar] [CrossRef]
- Verma, K.K.; Xiu-Peng, S.; Yuan, Z.; Dong-Mei, L.; Dao-Jun, G.; Vishnu, D.R.; Gan-Lin, C.; Anatoly, B.; Tatiana, M.M.; Yang-Rui, L. Characteristics of Leaf Stomata and Their Relationship with Photosynthesis in Saccharum officinarum Under Drought and Silicon Application. ACS Omega 2020, 5, 24145–24153. [Google Scholar] [CrossRef] [PubMed]
- Kafi, M.; Nabati, J.; Ahmadi-Lahijani, M.J.; Oskoueian, A. Silicon Compounds and Potassium Sulfate Improve Salinity Tolerance of Potato Plants through Instigating the Defense Mechanisms, Cell Membrane Stability, and Accumulation of Osmolytes. Commun. Soil Sci. Plant Anal. 2021, 52, 843–858. [Google Scholar] [CrossRef]
- Souri, Z.; Khanna, K.; Karimi, N.; Ahmad, P. Silicon and Plants: Current Knowledge and Future Prospects. J. Plant Growth Regul. 2021, 40, 906–925. [Google Scholar] [CrossRef]
- Bokor, B. Expression of Genes for Si Uptake, Accumulation, and Correlation of Si with Other Elements in Ionome of Maize Kernel. Front. Plant Sci. 2017, 8, 1063. [Google Scholar] [CrossRef] [Green Version]
- Fatemi, H.; Esmaiel Pour, B.; Rizwan, M. Foliar application of silicon nanoparticles affected the growth, vitamin C, flavonoid, and antioxidant enzyme activities of coriander (Coriandrum sativum L.) plants grown in lead (Pb)-spiked soil. Environ. Sci. Pollut. Res. 2021, 28, 1417–1425. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, L.; Higgs, D. Effect of Silicon on Plant Growth and Mineral Nutrition of Maize Grown Under Water-Stress Conditions. J. Plant Nutr. 2006, 29, 1469–1480. [Google Scholar] [CrossRef]
- Hernandez-Apaolaza, L. Can silicon partially alleviate micronutrient deficiency in plants? A review. Planta 2014, 240, 447–458. [Google Scholar] [CrossRef]
- Kim, Y.H. Silicon Application to Rice Root Zone Influenced the Phytohormonal and Antioxidant Responses Under Salinity Stress. J. Plant Growth Regul. 2014, 33, 137–149. [Google Scholar] [CrossRef]
- Yue, W.; Linli, H.; Weibiao, L.; Dawuda, M.M.; Jian, L.; Jianming, X.; Zhi, F.; Calderón-Urrea, A.; Yu, J. Foliar Application of 5-Aminolevulinic Acid (ALA) Alleviates Nacl Stress in Cucumber (Cucumis sativus, L.) Seedlings Through the Enhancement of Ascorbate-Glutathione Cycle. Sci. Hortic. 2019, 257, 108761. [Google Scholar] [CrossRef]
- Korkmaz, A.; Korkmaz, Y.; Demirkıran, A.R. Enhancing Chilling Stress Tolerance of Pepper Seedlings by Exogenous Application of 5-Aminolevulinic Acid. Environ. Exp. Bot. 2010, 67, 495–501. [Google Scholar] [CrossRef]
- Kramer, D.M.; Johnson, G.; Kiirats, O.; Edwards, G.E. New Fluorescence Parameters for the Determination of Q A Redox State and Excitation Energy Fluxes. Photosynth. Res. 2004, 79, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Yu, H.; Li, Q.; Chai, L.; Jiang, W. Improving Plant Growth and Alleviating Photosynthetic Inhibition and Oxidative Stress from Low-Light Stress with Exogenous GR24 in Tomato (Solanum lycopersicum L.) Seedlings. Front. Plant Sci. 2019, 10, 490. [Google Scholar] [CrossRef] [PubMed]
- Min, Z.; Li, R.; Chen, L.; Zhang, Y.; Li, Z.; Liu, M.; Ju, Y.; Fang, Y. Alleviation of Drought Stress in Grapevine by Foliar-Applied Strigolactones. Plant Physiol. Biochem. 2019, 135, 99–110. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source Element | Molarity (0.1 mM Ca) | Molarity (4 mM Ca) |
|---|---|---|
| NH4NO3 | 4.9 mM | 1 mM |
| Ca(NO3)2·4H2O | 0.1 mM | 4 mM |
| KNO3 | 5 mM | 5 mM |
| MgSO4·7H2O | 2 mM | 2 mM |
| KH2PO4 | 1 mM | 1 mM |
| H3BO3 | 0.045 mM | 0.045 mM |
| MnCl4·4H2O | 0.01 mM | 0.01 mM |
| ZnSO4·7H2O | 0.8 μM | 0.8 μM |
| H2MoO4 | 0.4 μM | 0.4 μM |
| CuSO4·5H2O | 0.3 μM | 0.3 μM |
| FeSO4·7H2O | 0.02 mM | 0.02 mM |
| EDTA−Na2 | 0.02 mM | 0.02 mM |
| Ca (mM) | Plant Height (cm) | Stem Diameter (mm) | Shoot | Root | ||
|---|---|---|---|---|---|---|
| Dry Weight (g) | Fresh Weight (g) | Dry Weight (g) | Fresh Weight (g) | |||
| 0 | 3.167 ± 0.139 g | 2.497 ± 0.669 f | 0.149 ± 0.014 h | 1.633 ± 0.049 g | 0.0248 ± 0.0008 g | 0.4233 ± 0.02 f |
| 0.05 | 3.333 ± 0.053 g | 2.717 ± 0.074 ef | 0.176 ± 0.006 gh | 1.773 ± 0.033 g | 0.0278 ± 0.0005 f | 0.5833 ± 0.0145 e |
| 0.1 | 3.7667 ± 0.088 f | 3.057 ± 0.065 de | 0.195 ± 0.004 fg | 2.077 ± 0.026 f | 0.0333 ± 0.0005 e | 0.68 ± 0.0173 d |
| 0.25 | 4.233 ± 0.145 e | 3.4 ± 0.121 cd | 0.217 ± 0.003 f | 2.853 ± 0.041 e | 0.0393 ± 0.0005 d | 0.7533 ± 0.0145 c |
| 0.5 | 4.667 ± 0.088 d | 3.65 ± 0.05 bc | 0.247 ± 0.009 e | 2.94 ± 0.04 e | 0.0411 ± 0.0007 d | 0.7867 ± 0.020 c |
| 1 | 4.967 ± 0.088 d | 3.817 ± 0.91 b | 0.289 ± 0.013 d | 3.723 ± 0.069 d | 0.0441 ± 0.0012 c | 0.8467 ± 0.0176 b |
| 2.5 | 5.3 ± 0.115 c | 3.897 ± 0.41 ab | 0.338 ± 0.015 c | 4.283 ± 0.095 c | 0.047 ± 0.0009 b | 0.87 ± 0.0116 ab |
| 4 | 6.533 ± 0.145 a | 4.047 ± 0.384 ab | 0.488 ± 0.008 a | 5.313 ± 0.097 a | 0.0529 ± 0.0014 a | 0.91 ± 0.006 a |
| 5 | 5.633 ± 0.088 b | 4.243 ± 0.325 a | 0.451 ± 0.009 b | 4.653 ± 0.000 b | 0.0474 ± 0.0004 b | 0.8667 ± 0.012 ab |
| Si | Plant Height | Stem Diameter | Shoot | Root | ||
|---|---|---|---|---|---|---|
| (mM) | (cm) | (mm) | Dry Weight (g) | Fresh Weight (g) | Dry Weight (g) | Fresh Weight (g) |
| Root application of Si | ||||||
| 0 | 3.63 ± 0.115 d | 3.83 ± 0.127 e | 0.16 ± 0.006 e | 1.98 ± 0.039 d | 0.017 ± 0.001 e | 0.53 ± 0.026 d |
| 0.25 | 3.83 ± 0.145 d | 3.88 ± 0.061 e | 0.18 ± 0.003 e | 2.06 ± 0.041 d | 0.0195 ± 0.000 e | 0.64 ± 0.025 c |
| 0.5 | 4.5 ± 0.153 c | 4.38 ± 0.096 c | 0.25 ± 0.022 d | 2.23 ± 0.036 d | 0.0245 ± 0.001 d | 0.79 ± 0.017 b |
| 0.75 | 5.53 ± 0.176 d | 4.68 ± 0.036 ab | 0.34 ± 0.01 bc | 3.30 ± 0.132 b | 0.0394 ± 0.001 c | 0.88 ± 0.017 b |
| 1 | 6.23 ± 0.145 a | 4.84 ± 0.032 a | 0.42 ± 0.009 a | 4.22 ± 0.087 a | 0.0485 ± 0.001 a | 1.036 ± 0.064 a |
| 1.5 | 4.533 ± 0.145 c | 4.50 ± 0.045 bc | 0.35 ± 0.019 b | 2.83 ± 0.1 c | 0.0419 ± 0.001 bc | 0.833 ± 0.026 b |
| 2 | 4.367 ± 0.12 c | 4.14 ± 0.045 d | 0.31 ± 0.009 c | 2.73 ± 0.151 c | 0.0432 ± 0.001 b | 0.583 ± 0.020 cd |
| Foliar spray application of Si | ||||||
| 0 | 3.533 ± 0.115 c | 3.81 ± 0.096 c | 0.13 ± 0.013 e | 1.94 ± 0.056 e | 0.014 ± 0.001 e | 0.51 ± 0.012 d |
| 0.25 | 3.86 ± 0.145 c | 3.91 ± 0.027 c | 0.17 ± 0.008 d | 1.98 ± 0.087 e | 0.019 ± 0.00 d | 0.54 ± 0.012 d |
| 0.5 | 4.36 ± 0.067 b | 4.18 ± 0.115 b | 0.19 ± 0.003 d | 2.36 ± 0.116 d | 0.029 ± 0.001 c | 0.61 ± 0.009 c |
| 0.75 | 5.96 ± 0.067 a | 4.23 ± 0.078 b | 0.28 ± 0.023 c | 3.43 ± 0.098 c | 0.040 ± 0.001 b | 0.79 ± 0.017 b |
| 1 | 6.26 ± 0.088 a | 4.74 ± 0.033 a | 0.46 ± 0.021 a | 4.54 ± 0.057 a | 0.047 ± 0.001 a | 0.99 ± 0.032 a |
| 1.5 | 4.73 ± 0.12 b | 4.22 ± 0.018 b | 0.39 ± 0.006 b | 3.98 ± 0.078 b | 0.046 ± 0.002 a | 0.80 ± 0.028 b |
| 2 | 4.6 ± 0.153 b | 4.14 ± 0.052 b | 0.31 ± 0.013 c | 2.57 ± 0.056 d | 0.045 ± 0.002 a | 0.48 ± 0.020 d |
| Gene | Gene ID | Primer Sequence (5′ to 3′) | Gene Function |
|---|---|---|---|
| Rubisco | Solyc07g066530 | F: CGCTCAAACCTTAGTCACCCTCAAG | Rubisco auxiliary enzyme (rubisco) |
| R: TGCCAGAACCATCTCACTCCTATCC | |||
| RCA | Solyc10g086580 | F: ATCGGATGACCAACAGGACATTGC | Ribulose diphosphate carboxylase/oxygenase activator enzyme (RCA) |
| R: CTTGACCTTTGCCTCCCCATACAC | |||
| SBPcase | Solyc05g052600 | F: GGGATCACAGGAAGAGAGCAAGTTG | 1, 7−diphosphate sedum heptanose−esterase (SBPcase) |
| R: TCAAGAATCCTAACGGTGCCACTTC | |||
| TK | Solyc10g018300 | F: TTGGCTTGATCCCGTATTGTGCTAC | Transketoolase (TK) |
| R: GCTCCTGCTGTCTCATTACCATCTG | |||
| Ru5PK | Solyc08g076220 | F: CCATGTACGATGAGCGTGTGAGAG | Ribulose 5−phosphate kinase (Ru5PK) |
| R: TGAGTTGGGAGCACTTCAATGACTG | |||
| NADP−GAPDH | Solyc12g094640 | F: CAAAGGCTGTGTCTCTAGTGCTACC | 3−phosphoglycerate dehydrogenase (NADP−GAPDH) |
| R: CTTGACCATGTCGTCTCCCATAACC | |||
| ATP | Solyc06g072540 | F: GGCGACGGTTTGTTGATACAAGAAG | ATP synthase subunit (ATP) |
| R: AAGAGGCTCATATACGGAACAACGC | |||
| Actin | Solyc04g011500.3 | F: TGGGTCAAAAAGACGCCTATG | |
| R: ATAATCTGGGTCATCTTTTCACGA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Z.; Liu, Z.; Yue, Z.; Wang, J.; Jin, L.; Xu, Z.; Jin, N.; Zhang, B.; Lyu, J.; Yu, J. Application of Exogenous Silicon for Alleviating Photosynthetic Inhibition in Tomato Seedlings under Low−Calcium Stress. Int. J. Mol. Sci. 2022, 23, 13526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113526
Li Z, Liu Z, Yue Z, Wang J, Jin L, Xu Z, Jin N, Zhang B, Lyu J, Yu J. Application of Exogenous Silicon for Alleviating Photosynthetic Inhibition in Tomato Seedlings under Low−Calcium Stress. International Journal of Molecular Sciences. 2022; 23(21):13526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113526
Chicago/Turabian StyleLi, Zhaozhuang, Zeci Liu, Zhibin Yue, Jie Wang, Li Jin, Zhiqi Xu, Ning Jin, Bo Zhang, Jian Lyu, and Jihua Yu. 2022. "Application of Exogenous Silicon for Alleviating Photosynthetic Inhibition in Tomato Seedlings under Low−Calcium Stress" International Journal of Molecular Sciences 23, no. 21: 13526. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113526