
Preliminary Expression Analysis of the OSCA Gene Family in Maize and Their Involvement in Temperature Stress
 ,
,
Abstract
:1. Introduction
2. Results
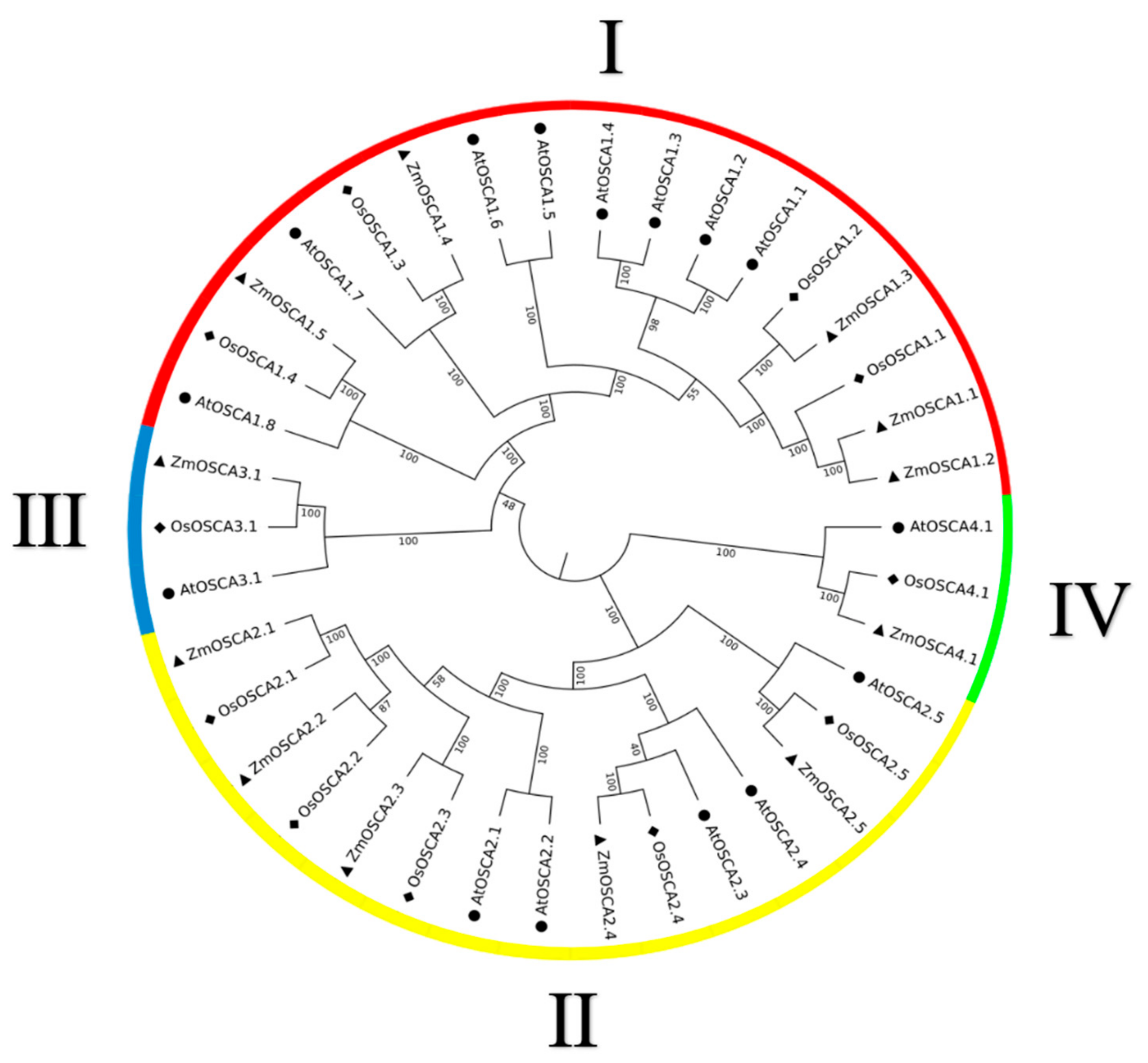
2.1. Phylogenetic Relationship and Gene Structure of Maize OSCAs
2.2. Analysis of Structural Domains of the ZmOSCA Family
2.3. Predicted Secondary Structure Analysis of ZmOSCA Gene Family Proteins
2.4. Characterization of the ZmOSCA Gene Family for Tissue-Based Expression
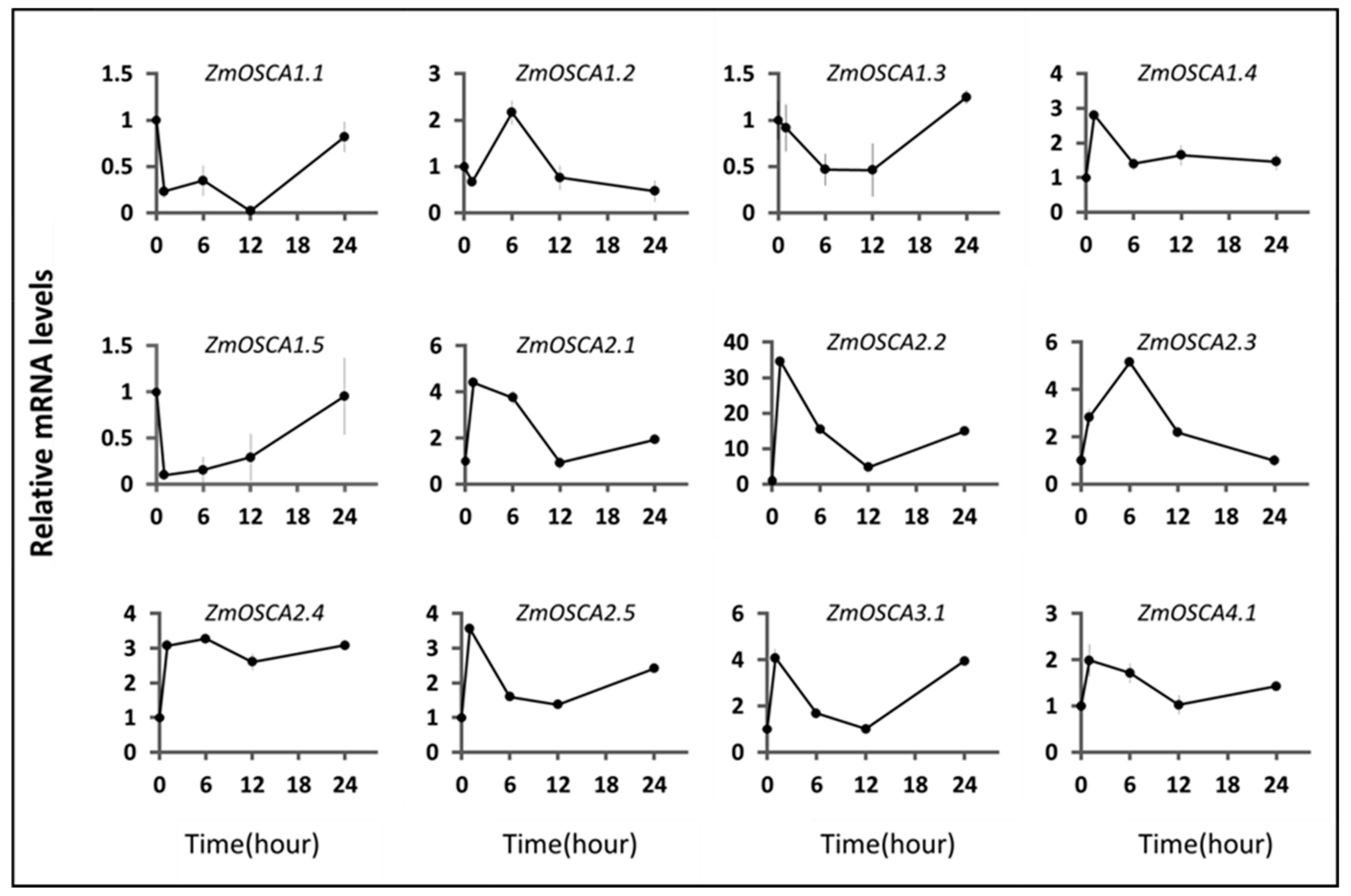
2.5. Expression Profiles of ZmOSCAs under Osmotic-Related Abiotic Stresses
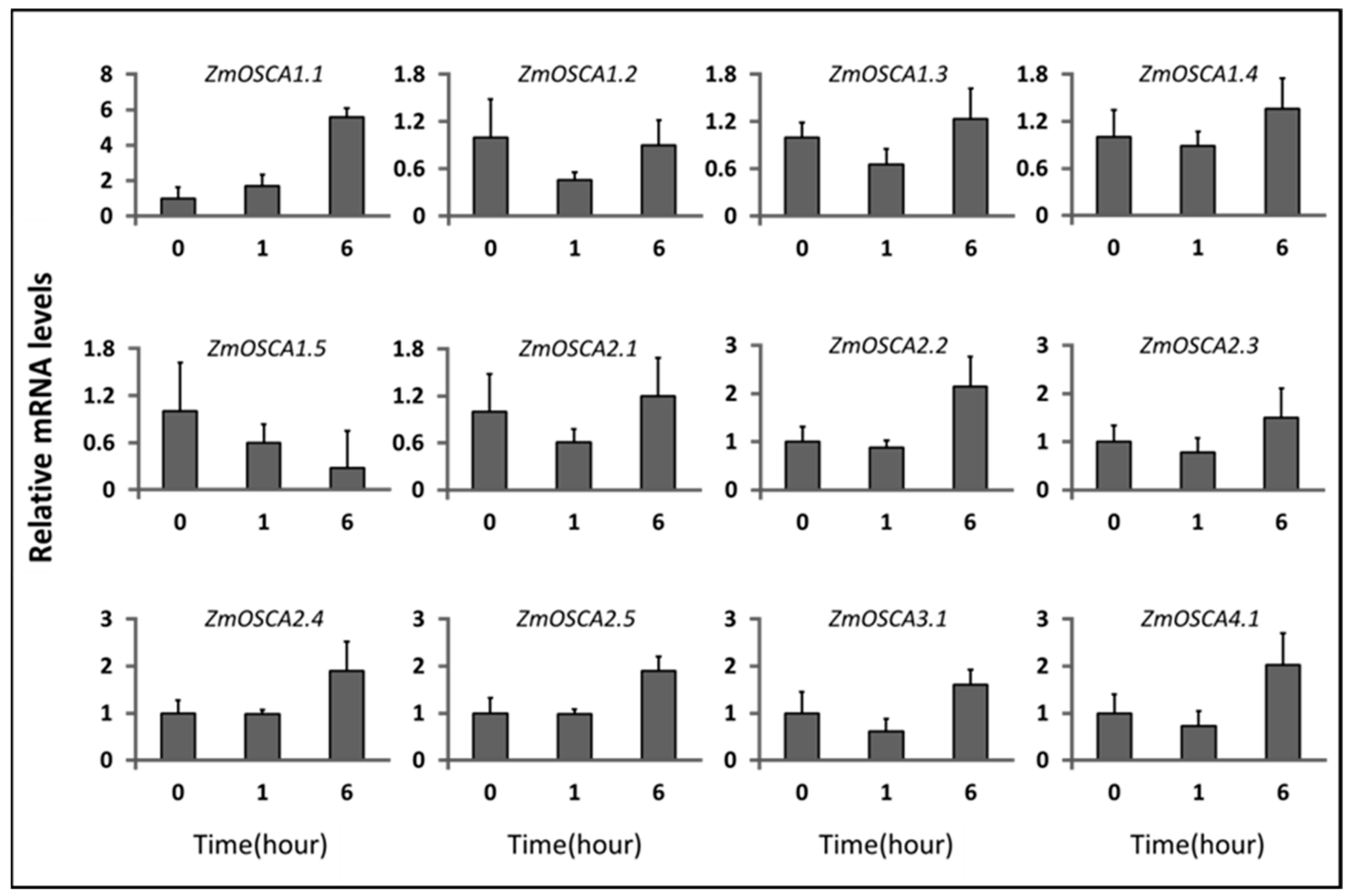
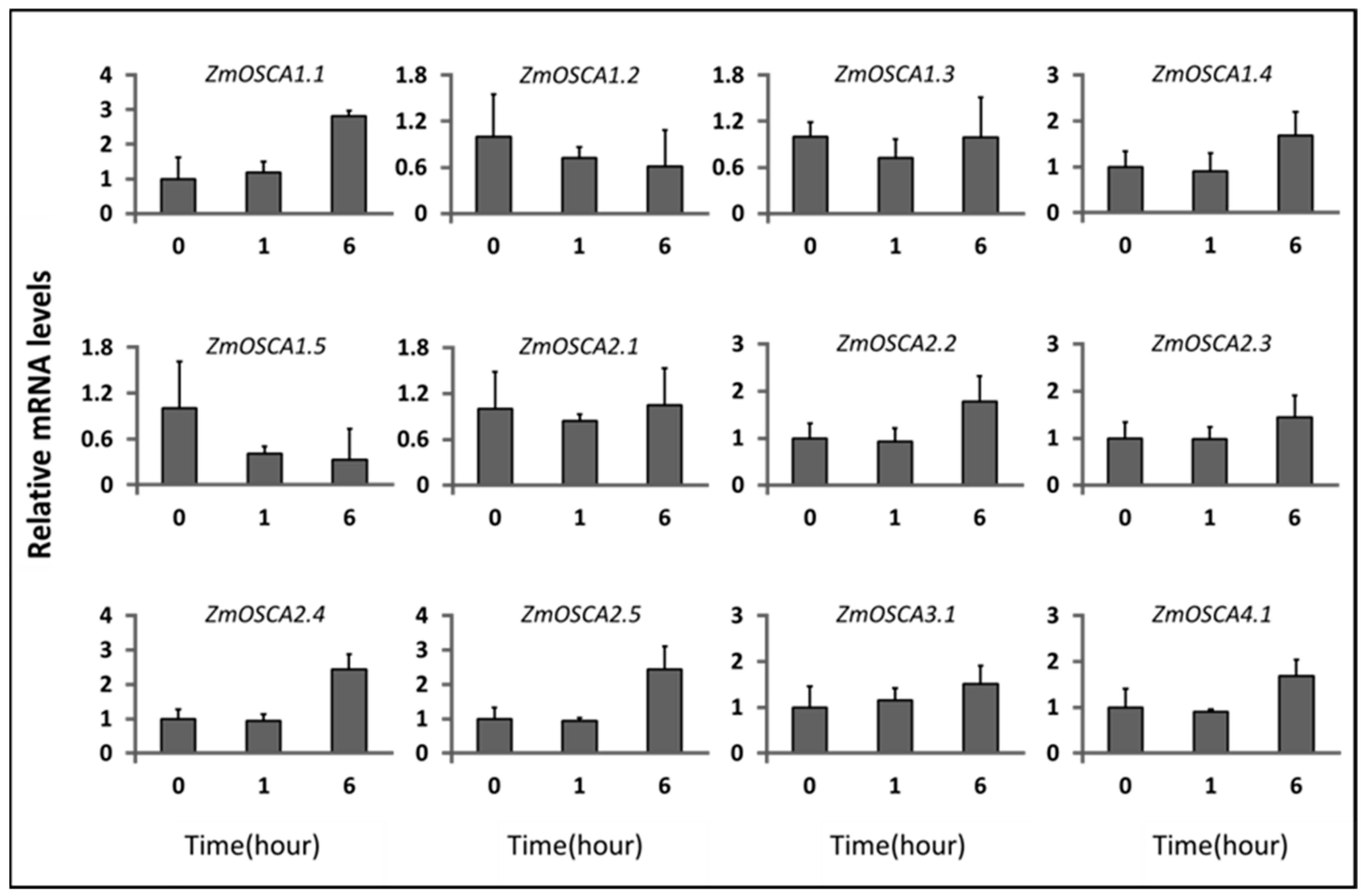
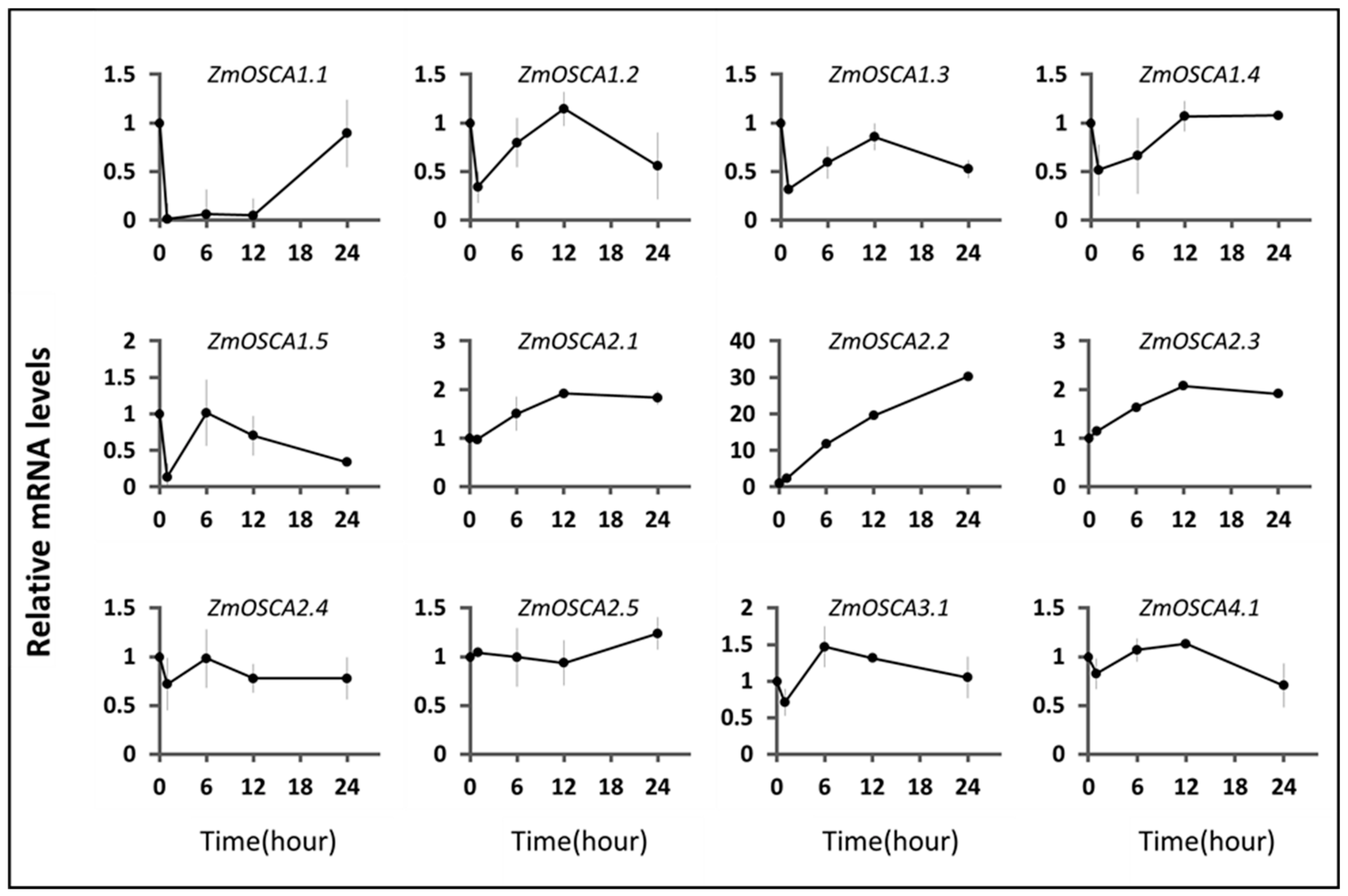
2.6. Expression Profile of ZmOSCAs under Environment-Related Abiotic Stresses
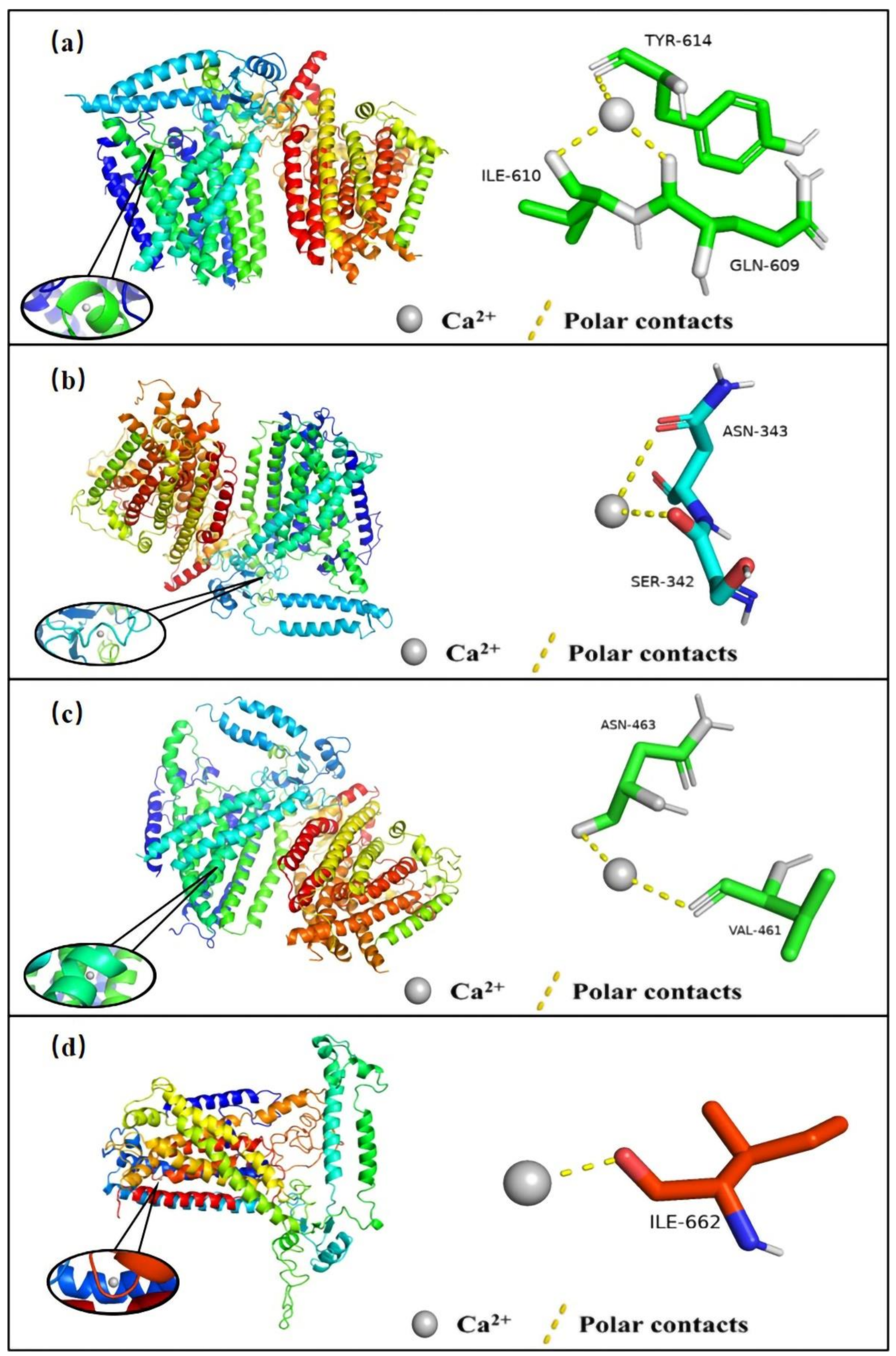
2.7. Prediction of Binding Sites for Ca2+ in OSCA Proteins
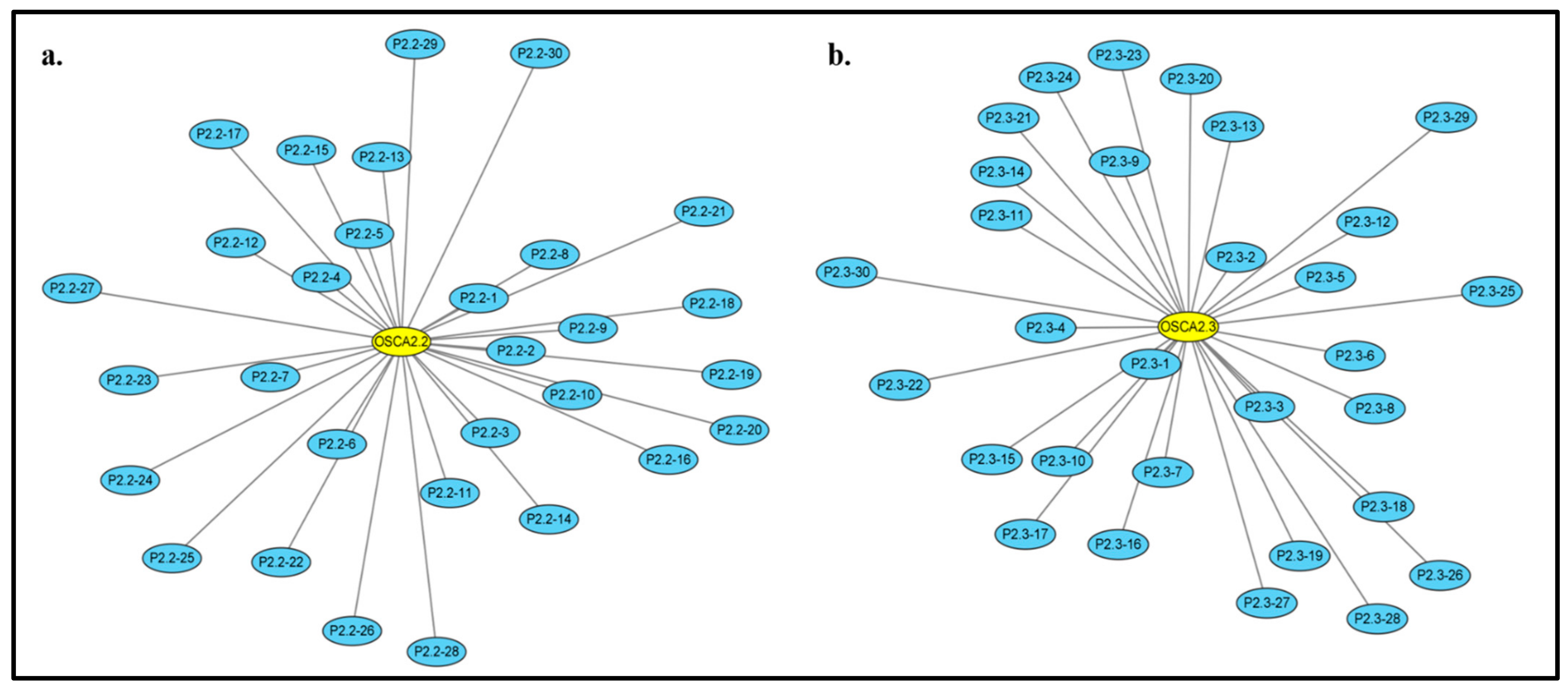
2.8. ZmOSCA2.2 and ZmOSCA2.3 Protein Prediction Analysis Results
3. Discussion
4. Materials and Methods
4.1. Bioinformatics Analysis
4.1.1. Gene Screening and Identification Analysis
4.1.2. Construction of Evolutionary Tree and Analysis of Gene Structure
4.1.3. Analysis of Protein Secondary Structure Prediction in Maize OSCA Family
4.1.4. Predictive Analysis of Protein Interactions
4.1.5. Prediction of Recognition Sites for Calcium Ions
4.2. Plant Materials and Growth Conditions
4.3. RNA Extraction and Quantitative qRT-PCR Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gong, F.P.; Yang, L.; Tai, F.J.; Hu, X.L.; Wang, W. “Omics” of Maize Stress Response for Sustainable Food Production: Opportunities and Challenges. Omics A J. Integr. Biol. 2014, 18, 714–732. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Y.; Hu, X.K.; Xu, W.; Fang, J.Y.; Shi, P.J. Increased probability and severity of compound dry and hot growing seasons over world’s major croplands. Sci. Total Environ. 2022, 824, 153885. [Google Scholar] [CrossRef] [PubMed]
- Krasensky, J.; Jonak, C. Drought, salt, and temperature stress-induced metabolic rearrangements and regulatory networks. J. Exp. Bot. 2012, 63, 1593–1608. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Zelicourt, A.; Colcombet, J.; Hirt, H. The Role of MAPK Modules and ABA during Abiotic Stress Signaling. Trends Plant Sci. 2016, 21, 677–685. [Google Scholar] [CrossRef]
- Zhang, L.L.; Zhang, Z.; Chen, Y.; Wei, X.; Song, X. Exposure, vulnerability, and adaptation of major maize-growing areas to extreme temperature. Nat. Hazards 2018, 91, 1257–1272. [Google Scholar] [CrossRef]
- Feng, S.F.; Hao, Z.C.; Zhang, X.; Hao, F.H. Probabilistic evaluation of the impact of compound dry-hot events on global maize yields. Sci. Total Environ. 2019, 689, 1228–1234. [Google Scholar] [CrossRef]
- Li, E.; Zhao, J.; Pullens, J.W.M.; Yang, X.G. The compound effects of drought and high temperature stresses will be the main constraints on maize yield in Northeast China. Sci. Total Environ. 2022, 812, 152461. [Google Scholar] [CrossRef]
- Lobell, D.B.; Gourdji, S.M. The Influence of Climate Change on Global Crop Productivity. Plant Physiol. 2012, 160, 1686–1697. [Google Scholar] [CrossRef] [Green Version]
- Mao, J.H.; Yu, Y.T.; Yang, J.; Li, G.K.; Li, C.Y.; Qi, X.T.; Wen, T.X.; Hu, J.G. Comparative transcriptome analysis of sweet corn seedlings under low-temperature stress. Crop J. 2017, 5, 396–406. [Google Scholar] [CrossRef]
- Zhu, J.K. Abiotic Stress Signaling and Responses in Plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef]
- Steinhorst, L.; Kudla, J. Calcium—A central regulator of pollen germination and tube growth. Biochim. Biophys. Acta 2013, 1833, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waters, M.T.; Langdale, J.A. The making of a chloroplast. EMBO J. 2009, 28, 2861–2873. [Google Scholar] [CrossRef] [PubMed]
- Hubbard, K.E.; Siegel, R.S.; Valerio, G.; Brandt, B.; Schroeder, J.I. Abscisic acid and CO2 signalling via calcium sensitivity priming in guard cells, new CDPK mutant phenotypes and a method for improved resolution of stomatal stimulus-response analyses. Ann. Bot. 2012, 109, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Kudla, J.; Rodriguez-Concepcion, M.; Yalovsky, S.; Gruissem, W. Calmodulins and calcineurin B-like proteins: Calcium sensors for specific signal response coupling in plants. Plant Cell 2002, 14, S389–S400. [Google Scholar] [CrossRef] [Green Version]
- Hepler, P.K. Calcium: A central regulator of plant growth and development. Plant Cell 2005, 17, 2142–2155. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.M.; Xue, Y.; Kong, D.D.; Ye, R.; Li, C.J.; Zhang, J.Y.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–378. [Google Scholar] [CrossRef]
- Cao, L.R.; Zhang, P.Y.; Lu, X.M.; Wang, G.R.; Wang, Z.H.; Zhang, Q.J.; Zhang, X.; Wei, X.; Mei, F.J.; Wei, L.; et al. Systematic Analysis of the Maize OSCA Genes Revealing ZmOSCA Family Members Involved in Osmotic Stress and ZmOSCA2.4 Confers Enhanced Drought Tolerance in Transgenic Arabidopsis. Int. J. Mol. Sci. 2020, 21, 351. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.X.; Chen, L. Recent Progress on The Studies of OSCA/TMEM63 Family Ion Channels. Prog. Biochem. Biophys. 2020, 47, 123–130. [Google Scholar]
- Hou, C.C.; Tian, W.; Kleist, T.; He, K.; Garcia, V.; Bai, F.L.; Hao, Y.L.; Luan, S.; Li, L.G. DUF221 proteins are a family of osmosensitive calcium-permeable cation channels conserved across eukaryotes. Cell Res. 2014, 24, 632–635. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.S.; Yuan, F.; Wen, Z.H.; Li, Y.H.; Wang, F.; Zhu, T.; Zhuo, W.Q.; Jin, X.; Wang, Y.D.; Zhao, H.P.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. Bmc Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef] [Green Version]
- Lv, G.D. Cloning, Marker Development and Functional Analysis of the Wheat TaOSCA1.4 Gene. Master’s Thesis, Shandong Agricultural University, Taian, China, 2015. [Google Scholar]
- Lee, H.W.; Park, J.H.; Park, M.Y.; Kim, J. GIP1 may act as a coactivator that enhances transcriptional activity of LBD18 in Arabidopsis. J. Plant Physiol. 2014, 171, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, T.; Tanaka, A.; Kawamoto, J.; Kurihara, T. Purification and characterization of 1-acyl-sn-glycerol-3-phosphate acyltransferase with a substrate preference for polyunsaturated fatty acyl donors from the eicosapentaenoic acid-producing bacterium Shewanella livingstonensis Ac10. J. Biochem. 2018, 164, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, P.; Diaz, A.R.; Re, M.F.; Porrini, L.; de Mendoza, D.; Albanesi, D.; Mansilla, M.C. Identification of Novel Thermosensors in Gram-Positive Pathogens. Front. Mol. Biosci. 2020, 7, 592747. [Google Scholar] [CrossRef] [PubMed]
- Sang-Hoon, L.; Jun, C.; KimKi, Y.; Chung, J.; Soo, P.H.; Dong-Gi, L.; Ki-Won, L. Identification of temperature stress-induced differentially expressed genes of tall fescue leaves. Res. J. Biotechnol. 2015, 10, 99–101. [Google Scholar]
- Hou, C.C. Isolation, Identification and Preliminary Functional Analysis of Calcium Permeable Cation Channels Gated by Hyperosmotic Stress. Ph.D. Thesis, Capital Normal University, Beijing, China, 2014. [Google Scholar]
- Xiong, L.; Schumaker, K.S.; Zhu, J.-K. Cell Signaling during Cold, Drought, and Salt Stress. Plant Cell 2002, 14, S165–S183. [Google Scholar] [CrossRef] [Green Version]
- Kumari, P.H.; Kumar, S.A.; Ramesh, K.; Reddy, P.S.; Nagaraju, M.; Prakash, A.B.; Shah, T.; Henderson, A.; Srivastava, R.K.; Rajasheker, G.; et al. Genome-Wide Identification and Analysis of Arabidopsis Sodium Proton Antiporter (NHX) and Human Sodium Proton Exchanger (NHE) Homologs in Sorghum bicolor. Genes 2018, 9, 236. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.B.; Ding, Y.L.; Shi, Y.T.; Ma, L.; Wang, Y.; Song, C.P.; Wilkins, K.A.; Davies, J.M.; Knight, H.; Knight, M.R.; et al. The calcium transporter ANNEXIN1 mediates cold-induced calcium signaling and freezing tolerance in plants. EMBO J. 2021, 40, e104559. [Google Scholar] [CrossRef]
- Tang, Y.M.; Zhao, C.P.; Gao, S.Q.; Tian, I.Q.; Shan, F.H.; Wu, J.X. Advances in the study of drought-related genes in plants. J. Triticeae Crops 2009, 29, 166–173. [Google Scholar]
- Schramm, F.; Larkindale, J.; Kiehlmann, E.; Ganguli, A.; Englich, G.; Vierling, E.; von Koskull-Doering, P. A cascade of transcription factor DREB2A and heat stress transcription factor HsfA3 regulates the heat stress response of Arabidopsis. Plant J. 2008, 53, 264–274. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.T.; Yu, Y.; Gao, X.R.; Pi, E.X. Progress in the study of MYB-like transcription factors regulating the mechanism of plant stress tolerance. J. Hangzhou Norm. Univ. 2021, 20, 621–627. [Google Scholar]
- Jung, C.; Seo, J.S.; Han, S.W.; Koo, Y.J.; Kim, C.H.; Song, S.I.; Nahm, B.H.; Do Choi, Y.; Cheong, J.-J. Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol. 2008, 146, 623–635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Luan, Y.; Meng, J.; Sun, J.; Tao, J.; Zhao, D. WRKY Transcription Factor Response to High-Temperature Stress. Plants 2021, 10, 2211. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.W. Identification and Stress Response Analysis of the OSCA Gene Family of Calcium Channels in Tomato. Master’s Thesis, Shandong Agricultural University, Taian, China, 2019. [Google Scholar]
- Thor, K.; Jiang, S.S.; Michard, E.; George, J.; Scherzer, S.; Huang, S.G.; Dindas, J.; Derbyshire, P.; Leitao, N.; DeFalco, T.A.; et al. The calcium-permeable channel OSCA1.3 regulates plant stomatal immunity. Nature 2020, 585, 569–573. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.X.; Fang, C.M.; He, Y.Q. Distribution and clustering analysis of hydrophobic proteins in the genome of rice blast fungus. Mol. Plant Breed. 2009, 7, 978–984. [Google Scholar]
- Kiefer, F.; Arnold, K.; Kunzli, M.; Bordoli, L.; Schwede, T. The SWISS-MODEL Repository and associated resources. Nucleic Acids Res. 2009, 37, D387–D392. [Google Scholar] [CrossRef]
- Li, B.; Hou, L.; Song, C.; Wang, Z.; Xue, Q.; Li, Y.; Qin, J.; Cao, N.; Jia, C.; Zhang, Y.; et al. Biological function of calcium-sensing receptor (CAS) and its coupling calcium signaling in plants. Plant Physiol. Biochem. 2022, 180, 74–80. [Google Scholar] [CrossRef]
- Wang, Z.B.; Li, H.; Wei, Z.P.; Sun, H.R.; He, Y.; Gao, J.; Yang, Z.M.; You, J.F. Overexpression of UDP-glycosyltransferase genes enhanced aluminum tolerance through disrupting cell wall polysaccharide components in soybean. Plant Soil 2021, 469, 135–147. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Position | Number of Transcripts | Number of Exons | Length (AA) | Strand |
|---|---|---|---|---|---|
| ZmOSCA1.1 | Contig B73V4_ctg31 243,552–267,664 | 9 | 11 | 768 | forward strand |
| ZmOSCA1.2 | Chr3 231,673,936–231,685,374 | 15 | 13 | 785 | forward strand |
| ZmOSCA1.3 | Chr8 80,433,878–80,440,706 | 6 | 12 | 790 | forward strand |
| ZmOSCA1.4 | Chr6 152,198,261–152,212,313 | 14 | 12 | 748 | reverse strand |
| ZmOSCA1.5 | Chr1 108,282,883–108,288,994 | 5 | 11 | 810 | reverse strand |
| ZmOSCA2.1 | Chr3 106,709,675–106,715,943 | 6 | 12 | 767 | forward strand |
| ZmOSCA2.2 | Chr1 261,919,725–261,930,103 | 2 | 12 | 765 | forward strand |
| ZmOSCA2.3 | Chr5 9,735,399–9,740,575 | 8 | 11 | 749 | forward strand |
| ZmOSCA2.4 | Chr1 164,016,950–164,031,018 | 5 | 10 | 699 | forward strand |
| ZmOSCA2.5 | Chr8 163,438,578–163,443,056 | 3 | 10 | 703 | reverse strand |
| ZmOSCA3.1 | Chr2 241,256,310–241,261,093 | 1 | 6 | 732 | reverse strand |
| ZmOSCA4.1 | Chr9 155,107,951–155,110,341 | 1 | 1 | 796 | forward strand |
| Species | Total | Group Ⅰ | Group Ⅱ | Group Ⅲ | Group Ⅳ |
|---|---|---|---|---|---|
| Arabidopsis thaliana | 15 | 8 | 5 | 1 | 1 |
| Oryza sativa | 11 | 4 | 5 | 1 | 1 |
| Zea mays | 12 | 5 | 5 | 1 | 1 |
| Gene | Alpha Helix (Hh) | Extended Strand (Ee) | Beta Ture (Tt) | Random Coil (Cc) | ||||
|---|---|---|---|---|---|---|---|---|
| ZmOSCA1.1 | 317 | 41.28% | 178 | 23.18% | 44 | 5.73% | 229 | 29.82% |
| ZmOSCA1.2 | 331 | 42.17% | 175 | 22.29% | 46 | 5.86% | 233 | 29.68% |
| ZmOSCA1.3 | 338 | 42.78% | 170 | 21.52% | 51 | 6.46% | 231 | 29.24% |
| ZmOSCA1.4 | 302 | 40.37% | 167 | 22.33% | 60 | 8.02% | 219 | 29.28% |
| ZmOSCA1.5 | 380 | 46.91% | 151 | 18.64% | 50 | 6.17% | 229 | 28.27% |
| ZmOSCA2.1 | 322 | 41.98% | 173 | 22.56% | 50 | 6.52% | 222 | 28.94% |
| ZmOSCA2.2 | 292 | 28.17% | 166 | 21.70% | 53 | 6.93% | 254 | 33.20% |
| ZmOSCA2.3 | 337 | 44.99% | 165 | 22.03% | 63 | 8.41% | 184 | 24.57% |
| ZmOSCA2.4 | 315 | 45.06% | 147 | 21.03% | 42 | 6.01% | 195 | 27.90% |
| ZmOSCA2.5 | 332 | 47.23% | 128 | 18.21% | 56 | 7.97% | 187 | 26.60% |
| ZmOSCA3.1 | 329 | 44.95% | 154 | 21.04% | 52 | 7.10% | 197 | 26.91% |
| ZmOSCA4.1 | 400 | 50.25% | 122 | 15.33% | 50 | 6.28% | 224 | 28.14% |
| Identifier | Protein Annotations | Length (AA) | Protein Families (COGs): | Protein Families Function | |
|---|---|---|---|---|---|
| P2.2-1 | GRMZM2G072052_P01 | Putative calcium-binding protein CML15 | 234 | COG5126 | Ca2+ -binding protein, EF-hand superfamily |
| P2.2-2 | GRMZM2G121034_P01 | NAD(P)-binding Rossmann-fold superfamily protein; Retinol dehydrogenase 12; Uncharacterized protein; Belongs to the short-chain dehydrogenases/reductases (SDR) family | 367 | COG1028 | NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family |
| P2.2-3 | GRMZM2G450125_P01 | Beta-amylase | 573 | NOG256053 | Amylopectin maltohydrolase activity |
| P2.2-4 | GRMZM2G177570_P01 | Annotation not available | 707 | COG4886 | Leucine-rich repeat (LRR) protein |
| P2.2-5 | GRMZM2G046111_P01 | Plant invertase/pectin Methylesterase inhibitor superfamily protein | 243 | NOG10079 | Enzyme inhibitor activity |
| P2.2-6 | GRMZM2G045854_P01 | Gbf-interacting protein 1 isoform x1; Putative DUF1296 domain-containing family protein | 540 | NOG236453 | Protein of unknown function (DUF1296) |
| P2.2-7 | GRMZM2G033926_P01 | WAPL (Wings apart-like protein Regulation of heterochromatin) protein | 334 | COG0204 | 1-acyl-sn-glycerol-3-phosphate acyltransferase |
| P2.2-8 | GRMZM2G180054_P01 | Putative calcium-binding protein CML15 | 109 | COG5126 | Ca2+-binding protein, EF-hand superfamily |
| P2.2-9 | GRMZM2G025833_P01 | Beta-amylase | 544 | NOG256053 | Amylopectin maltohydrolase activity |
| P2.2-10 | GRMZM5G847466_P01 | Calcium-binding protein CML; Calmodulin | 172 | COG5126 | Ca2+-binding protein, EF-hand superfamily |
| P2.2-11 | GRMZM2G147014_P01 | Dehydrin COR410; Belongs to the plant dehydrin family | 290 | NOG17228 | Response to water |
| P2.2-12 | GRMZM2G474777_P01 | Tyrosine-sulfated glycopeptide receptor 1; Putative phytosulfokine receptor (LRR repeat-containing protein kinase) family protein | 665 | COG4886 | Leucine-rich repeat (LRR) protein |
| P2.2-13 | GRMZM2G154114_P01 | Annotation not available | 381 | COG0515 | Serine/threonine-protein kinase |
| P2.2-14 | GRMZM2G373522_P01 | Dehydrin3; Dehydrin; Uncharacterized protein; Belongs to the plant dehydrin family | 289 | NOG17228 | Response to water |
| P2.2-15 | GRMZM2G095452_P01 | Transcription repressor OFP12 | 269 | NOG269813 | Negative regulation of transcription, DNA-templated |
| P2.2-16 | GRMZM2G464885_P01 | Uncharacterized protein loc100192507; Short-chain dehydrogenase TIC 32 chloroplastic | 314 | COG1028 | NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family |
| P2.2-17 | GRMZM2G043191_P01 | Uncharacterized protein loc100279724; Type IV inositol polyphosphate 5-phosphatase 11 | 347 | COG5411 | Phosphatidylinositol dephosphorylation |
| P2.2-18 | GRMZM2G175423_P01 | Sorbitol dehydrogenase homolog 1; Sorbitol dehydrogenase | 366 | COG1063 | Threonine dehydrogenase or related Zn-dependent dehydrogenase |
| P2.2-19 | GRMZM2G009940_P02 | Short-chain dehydrogenase TIC 32 chloroplastic; Retinol dehydrogenase 14; Uncharacterized protein; Belongs to the short-chain dehydrogenases/reductases (SDR) family | 316 | COG1028 | NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family |
| P2.2-20 | GRMZM2G128577_P02 | Short-chain dehydrogenase TIC 32 chloroplastic; Belongs to the short-chain dehydrogenases/reductases (SDR) family | 315 | COG1028 | NAD(P)-dependent dehydrogenase, short-chain alcohol dehydrogenase family |
| P2.2-21 | GRMZM2G702253_P02 | Annotation not available | 304 | COG0345 | Pyrroline-5-carboxylate reductase |
| P2.2-22 | GRMZM2G119079_P01 | Dual-specificity protein-like phosphatase 1; Protein-tyrosine phosphatase mitochondrial 1 | 347 | KOG1719 | Dual specificity phosphatase |
| P2.2-23 | GRMZM2G029546_P03 | Dnaj heat shock n-terminal domain-containing protein; Electron transporter/heat shock protein-binding protein; Uncharacterized protein | 343 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.2-24 | GRMZM2G055238_P01 | Uncharacterized loc100272659; Uncharacterized protein | 403 | NOG06404 | Allantoin: proton symporter activity |
| P2.2-25 | GRMZM2G135746_P01 | Uncharacterized protein; Ureide permease 5 | 403 | NOG06404 | Allantoin: proton symporter activity |
| P2.2-26 | GRMZM2G060296_P01 | Signal recognition particle receptor subunit alpha; Signal recognition particle protein subunit 9 | 625 | COG0552 | Signal recognition particle GTPase |
| P2.2-27 | GRMZM2G025528_P05 | Uncharacterized protein; Ureide permease 5 | 242 | NOG06404 | Allantoin: proton symporter activity |
| P2.2-28 | GRMZM2G091891_P02 | Uncharacterized loc100502471; Ureide permease 5 | 411 | NOG06404 | Allantoin: proton symporter activity |
| P2.2-29 | GRMZM2G022837_P01 | Menaquinone-specific isochorismate synthase; Isochorismate synthase 2 chloroplastic | 562 | COG0147 | Anthranilate/para-aminobenzoate synthases component I |
| P2.2-30 | GRMZM2G319454_P01IDP592 | Uncharacterized protein loc100278112; Transmembrane-like protein | 170 | KOG4831 | Unnamed protein |
| Identifier | Protein Annotations | Length (AA) | Protein Families (COGs): | Protein Families Function | |
|---|---|---|---|---|---|
| P2.3-1 | GRMZM6G732597_P01 | Annotation not available | 732 | COG5594 | Calcium-activated cation channel activity |
| P2.3-2 | AC186577.3_FGP006 | Pectinesterase inhibitor 28; Cell wall/vacuolar inhibitor of fructosidase 2 | - | NOG14257 | Pectinesterase inhibitor activity |
| P2.3-3 | GRMZM2G368698_P02 | Cell wall/vacuolar inhibitor of fructosidase 2; C/VIF2; Uncharacterized protein | 214 | NOG14257 | Pectinesterase inhibitor activity |
| P2.3-4 | GRMZM2G300141_P01 | Cell wall/vacuolar inhibitor of fructosidase 2; Uncharacterized protein | 222 | NOG14257 | Pectinesterase inhibitor activity |
| P2.3-5 | GRMZM2G101945_P01 | Pectinesterase inhibitor 28; Cell wall/vacuolar inhibitor of fructosidase 2 | 233 | NOG14257 | Pectinesterase inhibitor activity |
| P2.3-6 | GRMZM5G891247_P01 | Uncharacterized protein; Cell wall/vacuolar inhibitor of fructosidase 2 | 222 | NOG14257 | Pectinesterase inhibitor activity |
| P2.3-7 | GRMZM2G055180_P01 | Ethylene-responsive transcription factor 2 | 279 | NOG243370 | Ethylene-activated signaling pathway |
| P2.3-8 | GRMZM2G466044_P01 | Annotation not available | 282 | NOG243370 | Ethylene-activated signaling pathway |
| P2.3-9 | GRMZM2G033926_P01 | WAPL (Wings apart-like protein regulation of heterochromatin) protein | 334 | COG0204 | 1-acyl-sn-glycerol-3-phosphate acyltransferase |
| P2.3-10 | GRMZM2G358139_P01 | Annotation not available | 282 | NOG01264 | S-adenosylmethionine-dependent methyltransferase activity |
| P2.3-11 | GRMZM2G059801_P01 | Annotation not available | 325 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.3-12 | GRMZM2G080516_P01 | AP2-EREBP transcription factor; Putative AP2/EREBP transcription factor superfamily protein; Uncharacterized protein | 270 | NOG243370 | Ethylene-activated signaling pathway |
| P2.3-13 | GRMZM2G029546_P03 | Dnaj heat shock n-terminal domain-containing protein; Electron transporter/heat shock protein-binding protein; Uncharacterized protein | 343 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.3-14 | GRMZM2G343149_P01 | DNAJ heat shock N-terminal domain-containing protein; DnaJ domain-containing protein; Uncharacterized protein | 473 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.3-15 | AC204277.3_FGP006 | Uncharacterized loc100383917; Putative methyltransferase PMT23 | - | NOG01264 | S-adenosylmethionine-dependent methyltransferase activity |
| P2.3-16 | GRMZM2G045249_P01 | Probable methyltransferase pmt23 isoform x1; Putative methyltransferase PMT23 | 508 | NOG01264 | S-adenosylmethionine-dependent methyltransferase activity |
| P2.3-17 | GRMZM2G473960_P01 | Uncharacterized protein loc100286326;Seed maturation protein | 112 | NOG44451 | Late embryogenesis abundant (LEA) group 1 |
| P2.3-18 | GRMZM2G060296_P01 | Signal recognition particle receptor subunit alpha; Signal recognition particle protein subunit 9 | 625 | COG0552 | Signal recognition particle GTPase |
| P2.3-19 | GRMZM2G054465_P01 | Uncharacterized protein loc109623449; Tryptophan synthase beta type 2 | 409 | COG0133 | Tryptophan synthase beta chain |
| P2.3-20 | GRMZM2G087758_P01 | Chaperone DnaJ-domain superfamily protein; DnaJ domain-containing protein; Uncharacterized protein | 258 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.3-21 | GRMZM2G064898_P01 | ORMDL family protein | 162 | COG5081 | Negative regulation of ceramide biosynthetic process |
| P2.3-22 | GRMZM2G319454_P01 | Uncharacterized protein loc100278112; Transmembrane-like protein | 170 | KOG4831 | Unnamed protein |
| P2.3-23 | GRMZM2G044720_P01 | Uncharacterized loc100273730; ORMDL family protein | 162 | COG5081 | Negative regulation of ceramide biosynthetic process |
| P2.3-24 | GRMZM2G125072_P01 | DNAJ heat shock N-terminal domain-containing protein; 3Fe-4S ferredoxin; Uncharacterized protein | 304 | COG0484 | DnaJ-class molecular chaperone with C-terminal Zn finger domain |
| P2.3-25 | GRMZM2G045854_P01 | Gbf-interacting protein 1 isoform x1; Putative DUF1296 domain-containing family protein | 540 | NOG236453 | Protein of unknown function (DUF1296) |
| P2.3-26 | GRMZM2G478664_P01 | Annotation not available | 1058 | COG0060 | Isoleucyl-tRNA synthetase |
| P2.3-27 | GRMZM2G175423_P01 | Sorbitol dehydrogenase homolog 1; Sorbitol dehydrogenase | 366 | COG1063 | Threonine dehydrogenase or related Zn-dependent dehydrogenase |
| P2.3-28 | GRMZM2G702253_P02 | Annotation not available | 304 | COG0345 | Pyrroline-5-carboxylate reductase |
| P2.3-29 | GRMZM2G474326_P01 | Pathogenesis-related genes transcriptional activator pti5; Ethylene-responsive transcription factor 2 | 229 | NOG243370 | Ethylene-activated signaling pathway |
| P2.3-30 | GRMZM5G820122_P01 | Abc transporter c family mrp4 precursor; ABC transporter that may affect phytic acid transport and compartmentalization. | 1510 | COG1132 | The ABC-type multidrug transport system, ATPase and permease component |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, Y.; Li, B.; Hou, L.; Yu, J.; Jia, C.; Wang, Z.; Chen, S.; Zhang, M.; Qin, J.; et al. Preliminary Expression Analysis of the OSCA Gene Family in Maize and Their Involvement in Temperature Stress. Int. J. Mol. Sci. 2022, 23, 13658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113658
Li Y, Zhang Y, Li B, Hou L, Yu J, Jia C, Wang Z, Chen S, Zhang M, Qin J, et al. Preliminary Expression Analysis of the OSCA Gene Family in Maize and Their Involvement in Temperature Stress. International Journal of Molecular Sciences. 2022; 23(21):13658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113658
Chicago/Turabian StyleLi, Yuanyang, Yubin Zhang, Bin Li, Liyuan Hou, Jianing Yu, Chengguo Jia, Zhe Wang, Siqi Chen, Mingzhe Zhang, Jianchun Qin, and et al. 2022. "Preliminary Expression Analysis of the OSCA Gene Family in Maize and Their Involvement in Temperature Stress" International Journal of Molecular Sciences 23, no. 21: 13658. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232113658