


Impact of G-Quadruplex Structures on Methylation of Model Substrates by DNA Methyltransferase Dnmt3a
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Selection and Design of Proteins and DNA Substrates
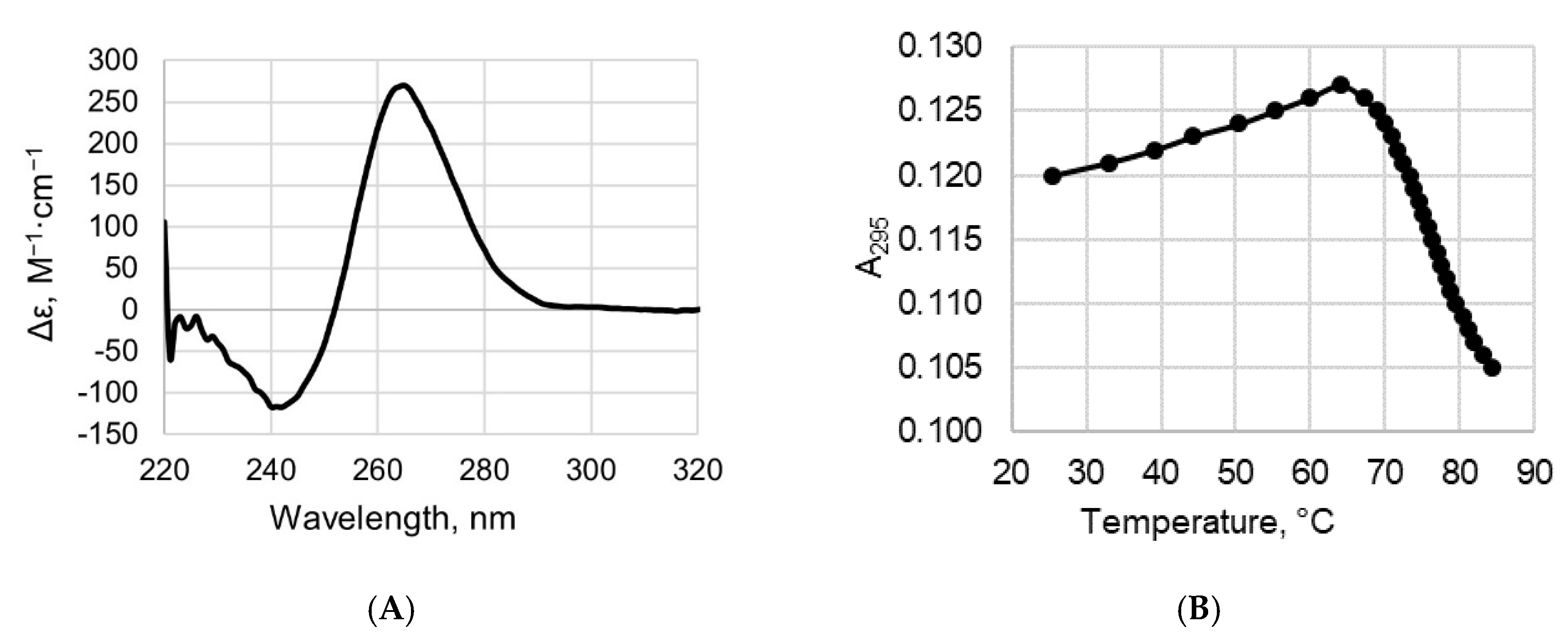
2.2. CD and UV Spectroscopy Confirm the Existence of a Parallel G4 Structure Folded within 19G4
2.3. Binding Affinity of AIR-Dnmt3a2 and Its Domains to DNA Substrates Containing the G4 Structure
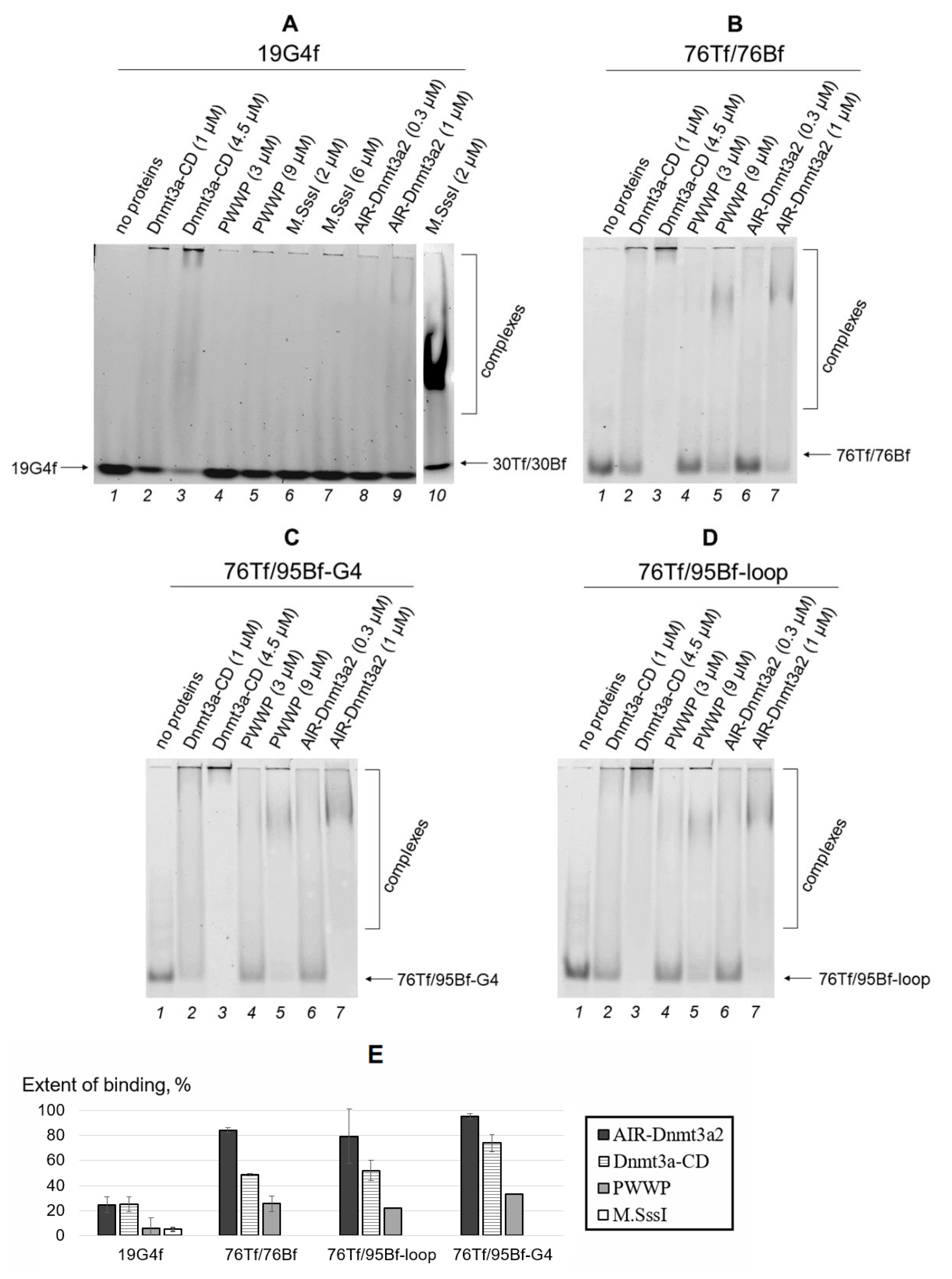
2.3.1. Electrophoretic Mobility Shift Assay (EMSA)
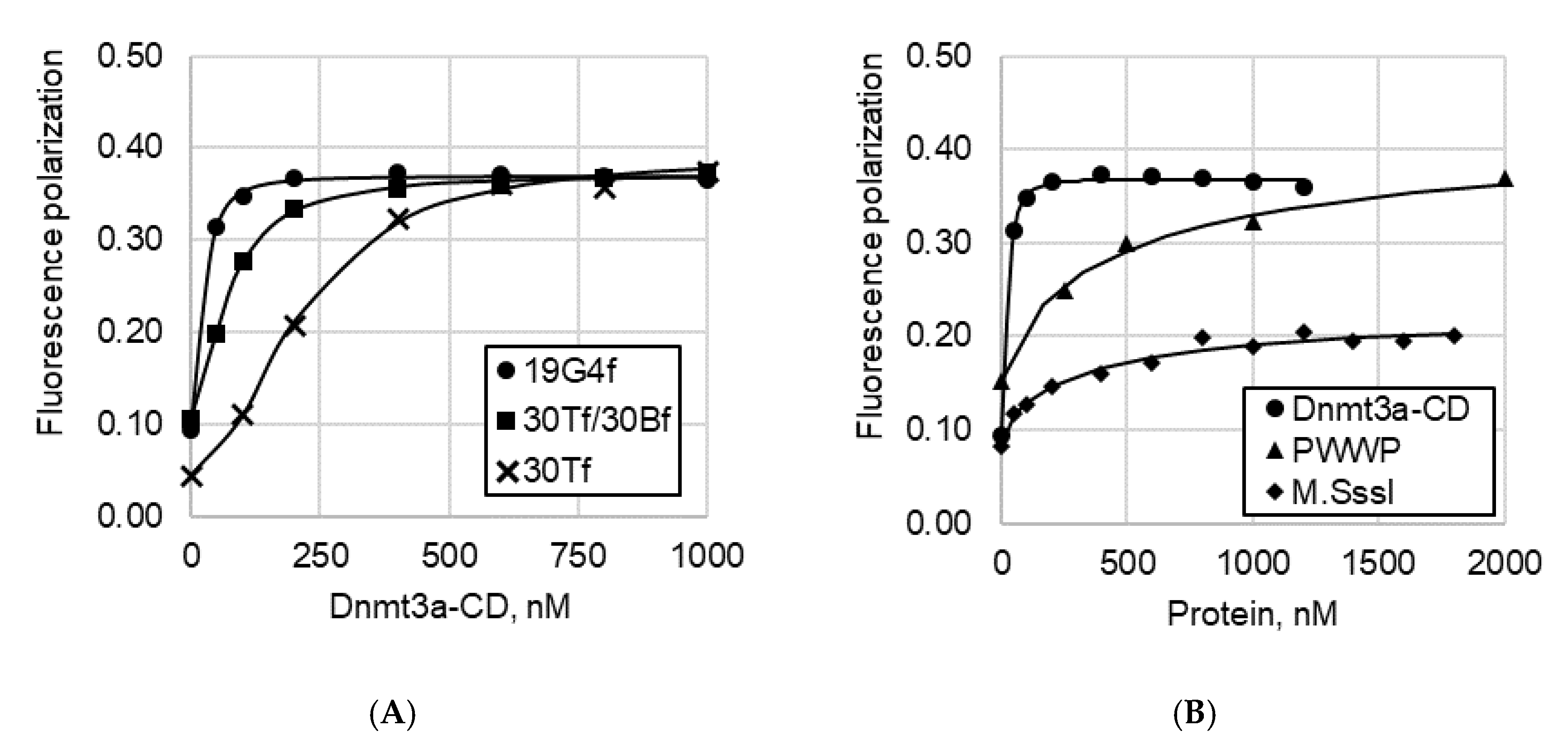
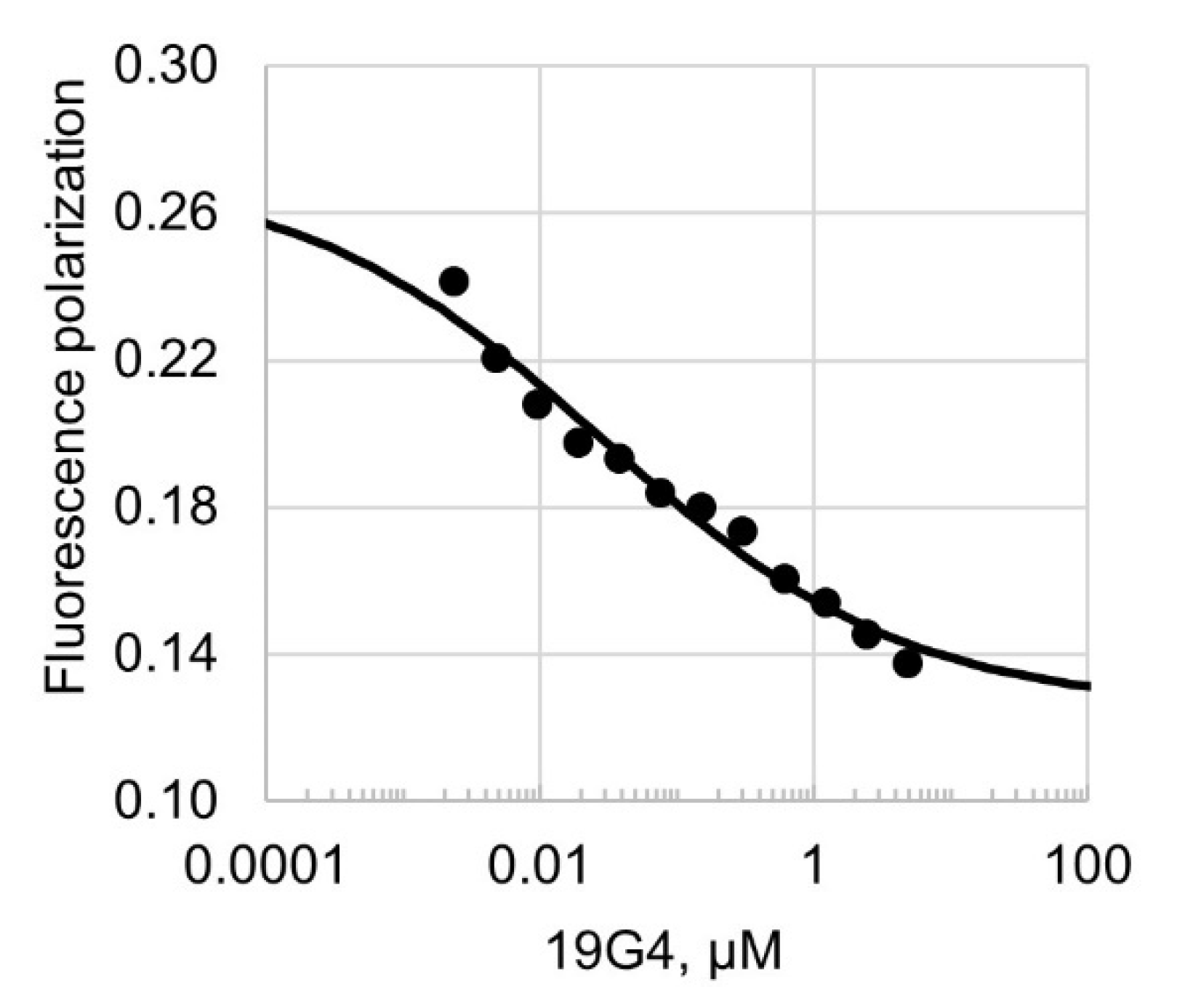
2.3.2. Fluorescence Polarization Assay
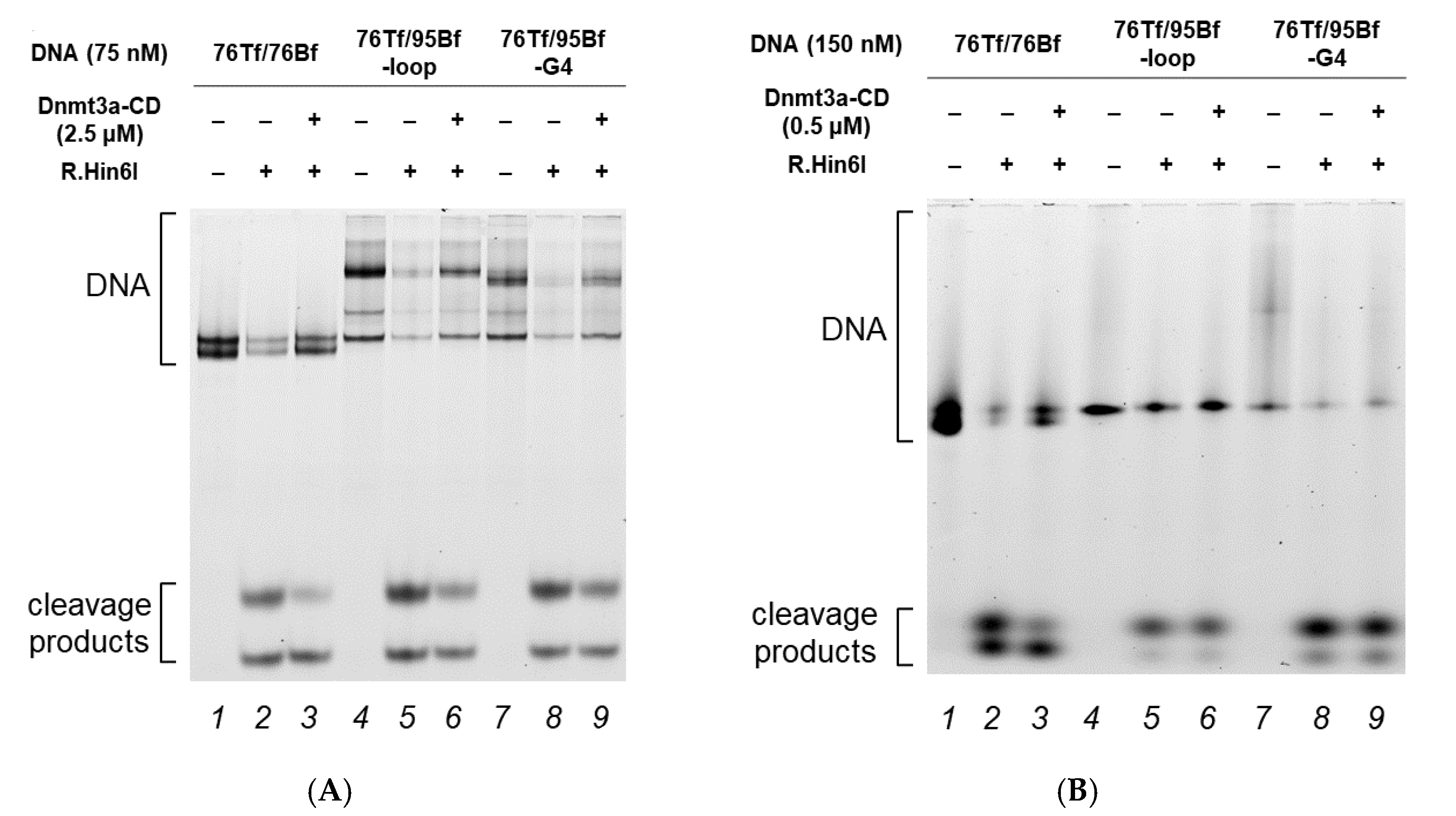
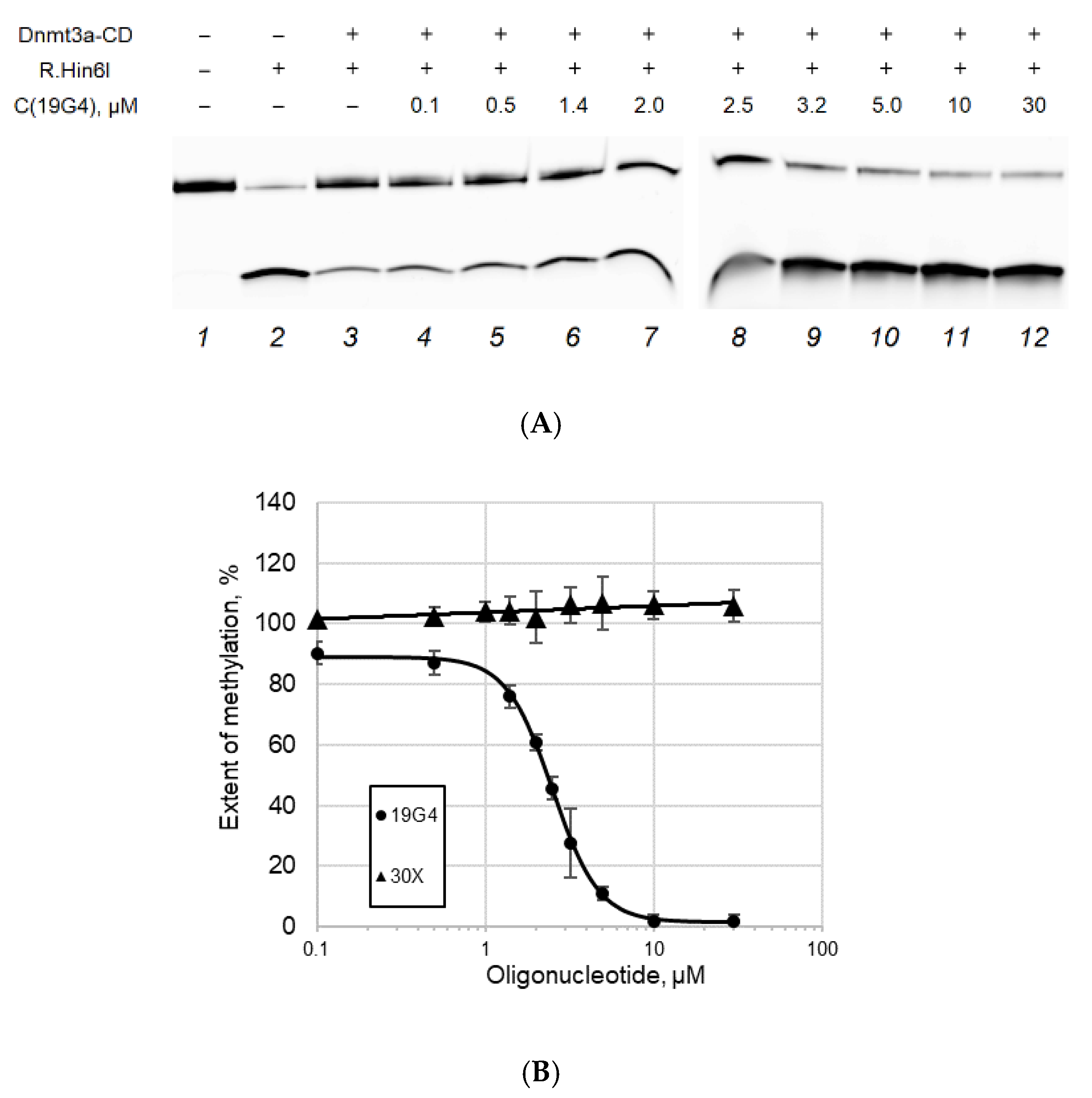
2.4. Effect of G4 Structures on Methylation by MTase Dnmt3a-CD
3. Discussion
3.1. Dnmt3a-CD, Unlike the PWWP Domain, Effectively Binds to G4 Structures
3.2. G4-Containing DNA Duplex Shows Improved Binding and Impaired Methylation by Dnmt3a-CD
3.3. G4-Forming Oligonucleotide 19G4 Inhibits Dnmt3a-CD Activity
4. Materials and Methods
4.1. Reagents
4.2. Enzymes and Proteins
4.3. Circular Dichroism Measurements
4.4. UV Spectroscopy Melting of G4 Structure
4.5. DNA Binding Measurements
4.5.1. Electrophoretic Mobility Shift Assay
4.5.2. Fluorescence Polarization Assay
4.6. Methylation Assay
4.6.1. Methylation of Double-Stranded DNA Substrates, including Those Containing G4 Structure
4.6.2. Effect of 19G4 on Methylation of 30Tf/30Bf by Dnmt3a-CD
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gowher, H.; Jeltsch, A. Mammalian DNA Methyltransferases: New Discoveries and Open Questions. Biochem. Soc. Trans. 2018, 46, 1191–1202. [Google Scholar] [CrossRef] [PubMed]
- Jeltsch, A.; Jurkowska, R.Z. New Concepts in DNA Methylation. Trends Biochem. Sci. 2014, 39, 310–318. [Google Scholar] [CrossRef] [PubMed]
- Wong, K.K.; Lawrie, C.H.; Green, T.M. Oncogenic Roles and Inhibitors of DNMT1, DNMT3A, and DNMT3B in Acute Myeloid Leukaemia. Biomark. Insights 2019, 14, 1177271919846454. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA Methyltransferases in Cancer: Biology, Paradox, Aberrations, and Targeted Therapy. Cancers 2020, 12, 2123. [Google Scholar] [CrossRef]
- Ehrlich, M. DNA Hypermethylation in Disease: Mechanisms and Clinical Relevance. Epigenetics 2019, 14, 1141–1163. [Google Scholar] [CrossRef]
- Hamidi, T.; Singh, A.K.; Chen, T. Genetic Alterations of DNA Methylation Machinery in Human Diseases. Epigenomics 2015, 7, 247–265. [Google Scholar] [CrossRef]
- Qin, Y.; Hurley, L.H. Structures, Folding Patterns, and Functions of Intramolecular DNA G-Quadruplexes Found in Eukaryotic Promoter Regions. Biochimie 2008, 90, 1149–1171. [Google Scholar] [CrossRef]
- Cui, X.; Chen, H.; Zhang, Q.; Xu, M.; Yuan, G.; Zhou, J. Exploration of the Structure and Recognition of a G-Quadruplex in the Her2 Proto-Oncogene Promoter and Its Transcriptional Regulation. Sci. Rep. 2019, 9, 3966. [Google Scholar] [CrossRef]
- Ducani, C.; Bernardinelli, G.; Högberg, B.; Keppler, B.K.; Terenzi, A. Interplay of Three G-Quadruplex Units in the KIT Promoter. J. Am. Chem. Soc. 2019, 141, 10205–10213. [Google Scholar] [CrossRef]
- Kaiser, C.E.; van Ert, N.A.; Agrawal, P.; Chawla, R.; Yang, D.; Hurley, L.H. Insight into the Complexity of the I-Motif and G-Quadruplex DNA Structures Formed in the KRAS Promoter and Subsequent Drug-Induced Gene Repression. J. Am. Chem. Soc. 2017, 139, 8522–8536. [Google Scholar] [CrossRef]
- Monsen, R.C.; DeLeeuw, L.W.; Dean, W.L.; Gray, R.D.; Chakravarthy, S.; Hopkins, J.B.; Chaires, J.B.; Trent, J.O. Long Promoter Sequences Form Higher-Order G-Quadruplexes: An Integrative Structural Biology Study of c-Myc, k-Ras and c-Kit Promoter Sequences. Nucleic Acids Res. 2022, 50, 4127–4147. [Google Scholar] [CrossRef] [PubMed]
- Dolinnaya, N.G.; Ogloblina, A.M.; Yakubovskaya, M.G. Structure, Properties, and Biological Relevance of the DNA and RNA G-Quadruplexes: Overview 50 Years after Their Discovery. Biochemistry 2016, 81, 1602–1649. [Google Scholar] [CrossRef] [PubMed]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative Visualization of DNA G-Quadruplex Structures in Human Cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Burge, S.; Parkinson, G.N.; Hazel, P.; Todd, A.K.; Neidle, S. Quadruplex DNA: Sequence, Topology and Structure. Nucleic Acids Res. 2006, 34, 5402–5415. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-Quadruplexes in Gene Promoters: A Novel Anticancer Strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Varizhuk, A.; Isaakova, E.; Pozmogova, G. DNA G-Quadruplexes (G4s) Modulate Epigenetic (Re)Programming and Chromatin Remodeling. BioEssays 2019, 41, 1900091. [Google Scholar] [CrossRef]
- Cree, S.L.; Fredericks, R.; Miller, A.; Pearce, F.G.; Filichev, V.; Fee, C.; Kennedy, M.A. DNA G-Quadruplexes Show Strong Interaction with DNA Methyltransferases in Vitro. FEBS Lett. 2016, 590, 2870–2883. [Google Scholar] [CrossRef]
- Mao, S.-Q.; Ghanbarian, A.T.; Spiegel, J.; Martínez Cuesta, S.; Beraldi, D.; di Antonio, M.; Marsico, G.; Hänsel-Hertsch, R.; Tannahill, D.; Balasubramanian, S. DNA G-Quadruplex Structures Mold the DNA Methylome. Nat. Struct. Mol. Biol. 2018, 25, 951–957. [Google Scholar] [CrossRef]
- Zhang, Z.M.; Lu, R.; Wang, P.; Yu, Y.; Chen, D.; Gao, L.; Liu, S.; Ji, D.; Rothbart, S.B.; Wang, Y.; et al. Structural Basis for DNMT3A-Mediated de Novo DNA Methylation. Nature 2018, 554, 387–391. [Google Scholar] [CrossRef]
- Jurkowska, R.Z.; Rajavelu, A.; Anspach, N.; Urbanke, C.; Jankevicius, G.; Ragozin, S.; Nellen, W.; Jeltsch, A. Oligomerization and Binding of the Dnmt3a DNA Methyltransferase to Parallel DNA Molecules: Heterochromatic Localization and Role of Dnmt3L. J. Biol. Chem. 2011, 286, 24200–24207. [Google Scholar] [CrossRef] [Green Version]
- Jurkowska, R.Z.; Jurkowski, T.P.; Jeltsch, A. Structure and Function of Mammalian DNA Methyltransferases. ChemBioChem 2011, 12, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Tsujimoto, N.; Li, E. The PWWP Domain of Dnmt3a and Dnmt3b Is Required for Directing DNA Methylation to the Major Satellite Repeats at Pericentric Heterochromatin. Mol. Cell. Biol. 2004, 24, 9048–9058. [Google Scholar] [CrossRef] [PubMed]
- Dhayalan, A.; Rajavelu, A.; Rathert, P.; Tamas, R.; Jurkowska, R.Z.; Ragozin, S.; Jeltsch, A. The Dnmt3a PWWP Domain Reads Histone 3 Lysine 36 Trimethylation and Guides DNA Methylation. J. Biol. Chem. 2010, 285, 26114–26120. [Google Scholar] [CrossRef] [PubMed]
- Chédin, F. The DNMT3 Family of Mammalian De Novo DNA Methyltransferases. Prog. Mol. Biol. Transl. Sci. 2011, 101, 255–285. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Wang, L.; Li, J.; Ding, Z.; Xiao, J.; Yin, X.; He, S.; Shi, P.; Dong, L.; Li, G.; et al. Structural Insight into Autoinhibition and Histone H3-Induced Activation of DNMT3A. Nature 2015, 517, 640–644. [Google Scholar] [CrossRef]
- Purdy, M.M.; Holz-Schietinger, C.; Reich, N.O. Identification of a Second DNA Binding Site in Human DNA Methyltransferase 3A by Substrate Inhibition and Domain Deletion. Arch. Biochem. Biophys. 2010, 498, 13–22. [Google Scholar] [CrossRef]
- Nur, I.; Szyf, M.; Razin, A.; Glaser, G.; Rottem, S.; Razin, S. Procaryotic and Eucaryotic Traits of DNA Methylation in Spiroplasmas (Mycoplasmas). J. Bacteriol. 1985, 164, 19–24. [Google Scholar] [CrossRef]
- Pavlova, A.; Monakhova, M.; Ogloblina, A.M.; Andreeva, N.A.; Laptev, G.Y.; Polshakov, V.I.; Gromova, E.S.; Zvereva, M.I.; Yakubovskaya, M.G.; Oretskaya, T.S.; et al. Responses of DNA Mismatch Repair Proteins to a Stable G-Quadruplex Embedded into a DNA Duplex Structure. Int. J. Mol. Sci. 2020, 21, 8773. [Google Scholar] [CrossRef]
- Sergeev, A.V.; Kirsanova, O.V.; Loiko, A.G.; Nomerotskaya, E.I.; Gromova, E.S. Detection of DNA Methylation by Dnmt3a Methyltransferase Using Methyl-Dependent Restriction Endonucleases. Mol. Biol. 2018, 52, 272–278. [Google Scholar] [CrossRef]
- Pavlova, A.V.; Kubareva, E.A.; Monakhova, M.V.; Zvereva, M.I.; Dolinnaya, N.G. Impact of G-Quadruplexes on the Regulation of Genome Integrity, DNA Damage and Repair. Biomolecules 2021, 11, 1284. [Google Scholar] [CrossRef]
- Klimasauskas, S.; Kumar, S.; Roberts, R.J.; Cheng, X. Hhal Methyltransferase Flips Its Target Base out of the DNA Helix. Cell 1994, 76, 357–369. [Google Scholar] [CrossRef]
- O’Hagan, M.P.; Mergny, J.-L.; Waller, Z.A.E. G-Quadruplexes in Prague: A Bohemian Rhapsody. Biochimie 2018, 147, 170–180. [Google Scholar] [CrossRef] [PubMed]
- Lipps, H.J.; Rhodes, D. G-Quadruplex Structures: In Vivo Evidence and Function. Trends Cell Biol. 2009, 19, 414–422. [Google Scholar] [CrossRef]
- Maizels, N. G4 Motifs in Human Genes. Ann. N. Y. Acad. Sci. 2012, 1267, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Emperle, M.; Rajavelu, A.; Reinhardt, R.; Jurkowska, R.Z.; Jeltsch, A. Cooperative DNA Binding and Protein/DNA Fiber Formation Increases the Activity of the Dnmt3a DNA Methyltransferase. J. Biol. Chem. 2014, 289, 29602–29613. [Google Scholar] [CrossRef]
- Khrabrova, D.A.; Loiko, A.G.; Tolkacheva, A.A.; Cherepanova, N.A.; Zvereva, M.I.; Kirsanova, O.; Gromova, E.S. Functional Analysis of DNMT3A DNA Methyltransferase Mutations Reported in Patients with Acute Myeloid Leukemia. Biomolecules 2019, 10, 8. [Google Scholar] [CrossRef]
- Wojciechowski, M.; Czapinska, H.; Bochtler, M. CpG Underrepresentation and the Bacterial CpG-Specific DNA Methyltransferase M.Mpel. Proc. Natl. Acad. Sci. USA 2013, 110, 105–110. [Google Scholar] [CrossRef]
- Jeltsch, A.; Jurkowska, R.Z. Multimerization of the Dnmt3a DNA Methyltransferase and Its Functional Implications, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2013; Volume 117, ISBN 9780123869319. [Google Scholar]
- Sergeev, A.; Tevyashova, A.N.; Vorobyov, A.P.; Gromova, E.S. The Effect of Antitumor Antibiotic Olivomycin A and Its New Semi-Synthetic Derivative Olivamide on the Activity of Murine DNA Methyltransferase Dnmt3a. Biochemistry 2019, 84, 62–70. [Google Scholar] [CrossRef]
- Sergeev, A.; Vorobyov, A.; Yakubovskaya, M.; Kirsanova, O.; Gromova, E.S. Novel Anticancer Drug Curaxin CBL0137 Impairs DNA Methylation by Eukaryotic DNA Methyltransferase Dnmt3a. Bioorg. Med. Chem. Lett. 2020, 30, 127296. [Google Scholar] [CrossRef]
- Brunetti, L.; Gundry, M.C.; Goodell, M.A. DNMT3A in Leukemia. Cold Spring Harb. Perspect. Med. 2017, 7, a030320. [Google Scholar] [CrossRef] [PubMed]
- Darii, M.; Kirsanova, O.; Drutsa, V.L.; Kochetkov, S.N.; Gromova, E.S. Isolation and Site-Directed Mutagenesis of DNA Methyltransferase SssI. Mol. Biol. 2007, 41, 110–117. [Google Scholar] [CrossRef]
- Lukashevich, O.; Baskunov, V.B.; Darii, M.; Kolbanovskiy, A.; Baykov, A.A.; Gromova, E.S. Dnmt3a-CD Is Less Susceptible to Bulky Benzo[a]Pyrene Diol Epoxide-Derived DNA Lesions than Prokaryotic DNA Methyltransferases. Biochemistry 2011, 50, 875–881. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Kd *, nM | ||
|---|---|---|---|
| 19G4f | Controls | ||
| 30Tf | 30Tf/30Bf | ||
| Dnmt3a-CD | 30 ± 7 | 200 ± 30 | 80 ± 20 |
| PWWP | 390 ± 130 | - | - |
| M.SssI | 320 ± 150 | - | - |
| DNA Duplex | Extent of DNA Methylation *, % | |
|---|---|---|
| Dnmt3a-CD:DNA Ratio | ||
| 33:1 | 3:1 | |
| 76Tf/76Bf | 56 ± 7 | 42 ± 5 |
| 76Tf/95Bf-G4 | 48 ± 8 | 8 ± 3 |
| 76Tf/95Bf-loop | 50 ± 5 | 12 ± 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Loiko, A.G.; Sergeev, A.V.; Genatullina, A.I.; Monakhova, M.V.; Kubareva, E.A.; Dolinnaya, N.G.; Gromova, E.S. Impact of G-Quadruplex Structures on Methylation of Model Substrates by DNA Methyltransferase Dnmt3a. Int. J. Mol. Sci. 2022, 23, 10226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810226
Loiko AG, Sergeev AV, Genatullina AI, Monakhova MV, Kubareva EA, Dolinnaya NG, Gromova ES. Impact of G-Quadruplex Structures on Methylation of Model Substrates by DNA Methyltransferase Dnmt3a. International Journal of Molecular Sciences. 2022; 23(18):10226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810226
Chicago/Turabian StyleLoiko, Andrei G., Alexander V. Sergeev, Adelya I. Genatullina, Mayya V. Monakhova, Elena A. Kubareva, Nina G. Dolinnaya, and Elizaveta S. Gromova. 2022. "Impact of G-Quadruplex Structures on Methylation of Model Substrates by DNA Methyltransferase Dnmt3a" International Journal of Molecular Sciences 23, no. 18: 10226. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms231810226