
Non-Photochemical Quenching: From Light Perception to Photoprotective Gene Expression


1
State Key Laboratory of Crop Stress Adaptation and Improvement, School of Life Sciences, Henan University, Jinming Avenue, Kaifeng 475004, China
2
State Key Laboratory of Crop Biology, College of Life Sciences, Shandong Agricultural University, Taian 271018, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(2), 687; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020687
Submission received: 12 December 2021
/
Revised: 5 January 2022
/
Accepted: 5 January 2022
/
Published: 8 January 2022
(This article belongs to the Special Issue Chloroplast and Stress Signaling)
{kind=link}
Abstract
:Light is essential for photosynthesis but light levels that exceed an organism’s assimilation capacity can cause serious damage or even cell death. Plants and microalgae have developed photoprotective mechanisms collectively referred to as non-photochemical quenching to minimize such potential damage. One such mechanism is energy-dependent quenching (qE), which dissipates excess light energy as heat. Over the last 30 years, much has been learned about the molecular mechanism of qE in green algae and plants. However, the steps between light perception and qE represented a gap in our knowledge until the recent identification of light-signaling pathways that function in these processes in the green alga Chlamydomonas reinhardtii. In this review, we summarize the high light and UV-mediated signaling pathways for qE in Chlamydomonas. We discuss key questions remaining about the pathway from light perception to photoprotective gene expression in Chlamydomonas. We detail possible differences between green algae and plants in light-signaling mechanisms for qE and emphasize the importance of research on light-signaling mechanisms for qE in plants.
1. Introduction
Photosynthesis in plants and algae is a biological process that converts light energy into chemical energy, which is used in the Calvin–Benson–Bassham cycle to assimilate CO2 and produce organic compounds. Light is required for photosynthesis. However, the supply of light changes under natural conditions and photosynthetic organisms are often exposed to excess light. Excess light leads to over-excitation of the photosystems, resulting in photodamage to the photosynthetic apparatus and possibly cell death. Plants and algae have therefore evolved a photoprotective mechanism called non-photochemical quenching (NPQ) to dissipate excess light energy as heat [1,2].
NPQ involves the thermal dissipation of excess absorbed energy through the deexcitation of singlet excited chlorophyll in photosystem II (PSII) and was originally defined based on both the relaxation kinetics of PSII components and their sensitivity to chemical inhibitors [2,3]. Several different processes contribute to NPQ and are differentiated based on the molecular players involved as well as on fluorescence relaxation kinetics [4]. These types of NPQ include qE, qH, qI, qM, qT, and qZ.
Energy-dependent quenching (qE), a major contributor to NPQ, occurs in the light-harvesting complexes of PSII (LHCII). qE involves rapid induction and relaxation [5,6]. qE responds to most short-term light stress and is both activated and deactivated rapidly (from seconds to minutes). The process of qE is driven by the acidification of the thylakoid lumen that occurs under excess light conditions [7]. This acidification modifies the pigment composition of LHCII via the xanthophyll cycle and activates specific qE protein effectors, such as PSBS and/or light-harvesting complex stress-related proteins (LHCSRs), in both green algae and plants [8,9,10]. In the active state, these protein effectors increase the energy dissipation capacity of LHCII via an unknown mechanism [11,12].
The recently identified antenna-quenching component qH protects the photosynthetic apparatus under stress conditions in Arabidopsis [13,14]. qH, a sustained form of antenna quenching, is considered to be a distinct NPQ component that is independent of PSBS, pH gradient, zeaxanthin, the serine/threonine kinase STN7, inactivation of the PSII core protein D1, and other qI processes. This photoprotective mechanism requires the plastid lipocalin LCNP and is prevented by the suppressor of quenching1 (SOQ1) under non-stress conditions. LCNP and the relaxation of qh1 (ROQH1) were recently proposed to play dosage-dependent antagonistic roles in protecting the photosynthetic apparatus and in maintaining the light-harvesting efficiency in plants [15].
Photoinhibitory quenching (qI) was previously defined as all mechanisms that result in the light-induced decrease in the quantum yield of PSII due to D1 photoinactivation [4,16]. qI, also known as sustained zeaxanthin, is associated with zeaxanthin retention. qI includes all components with slow relaxation kinetics, such as photoinhibition due to PSII photoinactivation and uncharacterized modes of sustained thermal dissipation [17]. qI is among the most slowly forming and slowly relaxing components of NPQ; depending on both photoinhibition and long-term photoprotective quenching, qI takes several hours or longer to relax [14,18].
qM is a blue light-dependent quenching mechanism induced by chloroplast movement [19,20]. However, chloroplast movement was recently shown to have little influence on the effectiveness of photoprotection under high-light conditions in “shade”-grown Arabidopsis. Therefore, the existence of a chloroplast movement-dependent component of NPQ and the influence of chloroplast movements on photoinhibition should be thoroughly reevaluated [21].
qT occurs under low-light conditions and depends on state transitions involving the movement of phosphorylated antenna proteins away from PSII [22,23]. qZ is a zeaxanthin-dependent NPQ process that does not require an acidic lumen or PSBS. Instead, qZ involves the binding of zeaxanthin to monomeric antenna proteins [24,25]. qZ relaxes more slowly than qE and takes several minutes to tens of minutes to turn on and off [24,26,27].
Due to the importance of NPQ and the rapid progress in understanding the underlying mechanisms, various aspects of NPQ have been reviewed over the past several years, such as light stress and photoprotection [28,29], NPQ mechanisms [2,30,31], the evolution of NPQ mechanisms and photoprotective antenna proteins [9,32,33,34], and the photoreceptor-dependent regulation of photoprotection [35].
In this review, we summarize what is known about the high light and UV-mediated signaling pathways for qE in Chlamydomonas. We discuss several key questions that remain about the processes between light perception and photoprotective gene expression in Chlamydomonas. Finally, we delve into possible differences between green algae and plants in response to excess light as well as the importance of research on light signaling for efforts to improve agricultural productivity by regulating qE.
2. Non-Photochemical Quenching Signaling Pathways in the Green Alga Chlamydomonas reinhardtii
2.1. Induction of Photoprotective Genes
qE is controlled by the photoprotective proteins LHCSRs and/or PSBS, which function as specific qE effectors [9]. The genes encoding these two photoprotective proteins are referred to as photoprotective genes or qE genes [36]. C. reinhardtii contains three genes encoding LHCSRs, namely LHCSR1, LHCSR3.1, and LHCSR3.2, and two genes encoding PSBS, namely PSBS1 and PSBS2. LHCSR genes are present in green algae and the model moss Physcomitrium (Physcomitrella) patens but not in vascular plants [8,37,38]. PSBS genes are found throughout green algae in P. patens and in vascular plants [5]. Thus, vascular plants and green algae appear to employ different proteins to regulate photosynthetic light harvesting under excess light conditions.
The expression of LHCSR genes is induced in response to environmental cues in the green alga C. reinhardtii. However, the expression patterns of LHCSR3 and LHCSR1 are different. LHCSR3 expression is induced by high light [8,36,39,40,41], Ca2+ [42], low CO2 [43], and nutrient starvation [44], whereas LHCSR1 expression is induced by UV-B [45] and nutrient starvation [44]. These results suggest that the expression of LHCSR genes is regulated by the environment where the green alga C. reinhardtii lives. The blue-light photoreceptor phototropin (PHOT) is essential for effective LHCSR3 gene expression and protein accumulation under high-light conditions [46,47]. The UV-B photoreceptor UVR8 can induce UV-dependent LHCSR1 and PSBS gene expression and protein accumulation [41,48]. The PSBS protein accumulates in Chlamydomonas only in response to UV-B [48] and very strong white light [49,50]. UV-B induces the accumulation of high levels of PSBS and LHCSR1, and much lower levels of LHCSR3, whereas high light induces the accumulation of high levels of LHCSR3 and lower levels of HCSR1 and PSBS [48,49,50]. The induction of photoprotective genes by UV-B is independent of photosynthetic electron transfer, whereas the induction of photoprotective genes under high light depends on this process [40,48]. The distinct expression patterns of PSBS, LHCSR1, and LHCSR3 under UV-B and high light point to the evolutionary divergence of these signaling pathways, which is likely associated with the different characteristics of these proteins [48].
2.2. High-Light Signaling Pathway
LHCSR3, the major qE effector in C. reinhardtii, primarily accumulates under excess light conditions. To gain insight into how this process is regulated, the light-color dependency (action spectrum) of qE induction between 400 nm and 720 nm was investigated in C. reinhardtii in Minagawa’s group [46]. Blue light was more effective than red light at inducing the qE response and LHCSR3 accumulation, suggesting that blue-light photoreceptors are involved in the accumulation of LHCSR3 in high light. To investigate whether blue-light photoreceptors are involved in inducing LHCSR3 accumulation, Petroutsos et al. (2016) compared the kinetics of NPQ, the action spectra of qE, and the induction of LHCSR3 accumulation in mutants lacking blue-light photoreceptors, namely the animal-like cryptochrome (aCRY) [46] and PHOT. This analysis revealed that PHOT controls qE by inducing the accumulation of LHCSR3 in high light. This control requires the perception of blue light by the LOV domains of PHOT; the induction of LHCSR3 through the kinase domain of PHOT; and light dissipation in photosystem II via LHCSR3. The phot mutants display severely reduced fitness under excessive light conditions, indicating that the sensing, utilization, and dissipation of light is a concerted process that plays a vital role in the acclimation of microalgae to environments with variable light intensities and PHOT is a central player in the photoprotection of C. reinhardtii [46].
The roles of various molecules in the sensing (by photoreceptors) and utilization (by photosynthetic complexes) of environmental light have long been unclear. The findings about the roles of PHOT described above uncover the molecular link between the sensing and utilization of light, the two essential functions of photosynthetic organisms. How does PHOT control the expression of LHCSR3.1 and LHCSR3.2? It appears that the LOV domains of PHOT provide blue-light sensitivity, while the kinase domain of PHOT performs signal transduction, possibly via the initiation of a cyclic nucleotide monophosphate (cAMP and/or cGMP)-signaling cascade. Downstream of PHOT, this signal is integrated with another regulatory signal from the chloroplast that carries information about the amount of absorbed light that is not used for CO2 fixation. The chloroplast signal relies on photosynthetic electron transfer [40,42], although the precise nature of this signal remains elusive. These integrated signals regulate the expression of LHCSR3.1 and LHCSR3.2 [46]. However, the signaling pathway between PHOT and LHCSR3 is not clear.
Minagawa’s group further revealed the possible signaling components between PHOT and LHCSR3 [47]. The phot mutant does not express LHCSR3 [46], its chlorophyll is bleached within 16 h, and the cells do not survive exposure to high-light conditions (1000 µmol photons m−2 s−1) [47], suggesting that the high-light sensitivity of this mutant is suppressed by inactivation of the negative regulators of LHCSR3 and/or LHCSR1 gene expression. The authors took advantage of this high-light sensitivity to obtain mutants in the signal transduction pathways between PHOT and the LHCSR3 transcription site. They identified two phot suppressor loci involved in qE quenching: de-etiolated 1 (det1) and damaged DNA-binding 1 (ddb1). When grown under low-light conditions (40 µmol photons m−2 s−1), the growth rates of the suppressors (det1-1 phot, det1-2 phot, and ddb1 phot) and phot mutants were similar to that of the wild type (WT), and the photosynthetic performance of the suppressors was similar to that of the original phot mutant. By contrast, under high-light conditions, the phot mutant did not survive and the growth of WT cells was retarded; however, the suppressors grew even more rapidly than WT cells, suggesting that the det1 and ddb1 mutations suppress the phot mutation and confer high-light tolerance. Moreover, the suppressors recovered the qE quenching capacity of the mutants. Thus, the increased tolerance to high-light conditions in the suppressors appears to be associated with their photoprotection ability. Through a combination of genetic, biochemical, and molecular analyses, the authors concluded that DET1 and DDB1 (two downstream components of PHOT) are involved in qE quenching and in the induction of photoprotective genes including LHCSR1/3 and PSBS under high-light conditions.
Next, Minagawa’s group investigated how DET1 and DDB1 regulate qE gene expression. Based on a yeast two-hybrid analysis and an inhibitor assay, the authors concluded that DET1 and DDB1 are part of a protein complex containing CULLIN 4 (CUL4). They proposed that DET1 and DDB1 act as central mediators by forming an E3 ubiquitin ligase complex together with CrCUL4 (CRL4DET1). This complex suppresses the expression of qE genes in the dark by directly ubiquitinating its targets and promotes the expression of these genes under high-light conditions by inhibiting the activity of the E3 ligase complex CUL4–DDB1DET1 via the PHOT signal.
Two questions are then raised: the possible targets of the E3 ligase complex CUL–DDB1DET1 and the possible signaling components that act downstream of this complex. Krishna Niyogi’s group recently identified SPA1 and CUL4, the components of a putative E3 ligase, as critical factors in a signaling pathway that controls the light-induced expression of qE genes. The authors proposed that the E3 ubiquitin ligase SPA1/COP1 acts upstream of CONSTANS (CrCO), a transcription factor that controls the qE capacity via cis-regulatory CrCO-binding sites in photoprotective genes [36]. Indeed, Jun Minagawa’s group showed that the transcription factors CrCO and Nuclear transcription Factor Ys (NF-Ys) form a complex that governs light-dependent photoprotective responses in C. reinhardtii. Moreover, they determined that the signal from the light perception to the CONSTANS/NF-Ys’ complex is directly inhibited by the E3 ubiquitin ligase SPA1/COP1 [41]. We note that in both studies, no accumulation of LHCRS1 or PSBS was induced by high light or UV in the crco mutants, while only a small amount of LHCRS3 accumulated in high light but not in UV [36,41], suggesting that CrCO is a key transcription factor controlling qE gene expression. Thus, CrCO likely functions downstream of the E3 ligase complex CUL4–DDB1DET1, although other transcription factors might be involved in regulating qE-associated genes as well.
To reveal the possible high-light signaling pathways, Aihara et al. (2019) investigated the transcript patterns of LHCSR1/3 and PSB1/2 in the phot mutant as well as the suppressors under high-light conditions [47]. High light induced a significant increase in the transcript levels of PSB1/2, LHCSR1, and LHCSR3.1/3.2 in the WT, whereas this high light-induced upregulation was suppressed in the phot mutant. These results reveal that there are at least two high light-induced signaling pathways for photoprotective genes: one that is PHOT-dependent and one that is PHOT-independent. Indeed, high levels of PSB1/2 and LHCSR1/3 transcripts were detected in both dark and high-light conditions in the det1 phot mutants. By contrast, in the det1 phot mutants, PSB1/2 and LHCSR1/3 were expressed at low levels in the dark but at high levels under high-light conditions, suggesting that the additional high-light signals are DET1-dependent. Therefore, LHCSR1 and PSBS1/2 expression is independent of active photosynthesis, whereas LHCSR3 expression is dependent on active photosynthesis, suggesting that there are two additional DET1-dependent high-light signaling pathways: one that is independent of active photosynthesis (for LHCSR1 and PSBS1/2 expression) and one that is dependent on active photosynthesis (for LHCSR3 expression; see Figure 1A).
It should be noted that the accumulation of LHSCR3 but not LHSCR1 or PSBS was induced by high light in the crco and crco spa1-1 mutants, pointing to the existence of an SPA1/CrCO-independent but high light-dependent pathway [36]. Notably, the phenotypes of the phot and crco mutants are similar, suggesting that PHOT and CrCO might act in the same pathway. Thus, PHOT might act as a photoreceptor in the SPA1/CrCO-independent pathway. It is also possible that the SPA1/CrCO-independent pathway is under the same regulation as the chloroplast-DET1-dependent high-light signaling pathway (see Figure 1A).
The blue light-dependent PHOT-signaling pathway is not limited to green algae but appears to be common to other photosynthetic organisms. This pathway might have been acquired during their evolution in a blue light-dominated environment, such as water columns. NPQ in cyanobacteria is also triggered by strong blue light [51,52]. Several lines of evidence point to a link between blue light and photoprotection in diatoms, although the underlying mechanism remains elusive [53,54,55,56,57].
2.3. UVR8-Signaling Pathway
UV-B is the most energy-rich component of sunlight and is potentially damaging to organisms. For example, UV-B has significant effects on photosynthetic processes [58]; photodamage peaks in the UV-B part of the spectrum [59,60]. In Chlamydomonas, various nucleus-encoded photoprotective genes, such as PSBS, LHSCR1, and LHSCR3, are induced by exposure to low doses of UV-B [45]. These findings indicate that there is a direct link between signaling by the UV-B photoreceptor UVR8 and photoprotection in Chlamydomonas. Indeed, under UV-B conditions, UVR8 and its partner COP1 can initiate a signaling pathway that induces the expression of photoprotective genes which strongly contributes to photoprotection under high light. Moreover, the competence for qE induced by acclimation to UV-B strongly contributes to photoprotection upon subsequent exposure to high light, suggesting that the UV-B signal might act as a proxy for high light, priming the cells for photoprotection [48].
Which factors mediate the induction of PSBS and LHCSR1 gene expression by the UVR8 signal? Several Deficient-in-LHCSR-expression (DSR) mutants (with reduced LHCSR gene expression) were obtained through a genetic screening via a bioluminescence reporter assay in Minagawa’s group [61]. Four mutants were recently characterized, including mutants of DSR10 and DSR15 with a mutation in CONSTANS and mutants of DSR28 and CC4286 with a mutation in NF-YB [41]. Compared to the WT (LHCSR1-Luc717), the DSR15 (crco-2) and DSR28 (nfyb-1) mutants exhibited pigment bleaching, a strong decrease in the maximal efficiency of PSII, an almost complete loss of qE capacity, and undetectable levels of key photoprotective proteins including LHCSR1, LHCSR3, and PSBS following exposure to excess light (including a low dose of UV-B), suggesting that both CrCO and NF-YB are essential for the functional activation of qE-dependent photoprotection in C. reinhardtii. The authors discovered that CrCO, NF-YB, and NF-YC interact with each other to form a CO/NF-YB/NF-YC complex and that all photoprotective genes (including LHCSR1, LHCSR3.1/3.2, and PSBS1/2) contain at least one CO-responsive element (CORE; CCACA22) as well as an NF-Y cis-element (CCAAT12) in the regions upstream of their start codons. Moreover, NF-YB associated with the promoters of the photoprotective genes in low light and this association appeared to be facilitated by exposure to high light. Considering the binding features of plant NF-YB/NF-YC in A. thaliana, these findings suggest that NF-YB weakly associates with the promoter regions of photoprotective genes under low light and that this association is reinforced by both CrCO accumulation and the formation of a CrCO/NF-YB/NF-YC complex under high light. Thus, the CrCO/NF-YB/NF-YC complex acts as a transcriptional module involved in regulating the expression of photoprotective genes in C. reinhardtii.
Then, Minagawa’s group further investigated the possible mechanism for the accumulation of CrCO, finding that the SPA1/COP1-dependent E3 ubiquitin ligase is involved in the degradation of this protein [41]. To clarify the link between the E3 ubiquitin ligase, CrCO accumulation, and photoprotective proteins, they treated various lines with the proteasome inhibitor MG132. Treatment of the crco-2/CrCO line with MG132 resulted in the accumulation of CrCO even under low-light conditions. The overaccumulation of photoprotective proteins (mainly LHCSR1 and PSBS) was also observed in the spa1 mutant even under low-light conditions due to the accumulation of CrCO. These findings indicate that the SPA1/COP1 modules function in the photoprotection in C. reinhardtii by controlling the degradation of CrCO. Under low-light conditions, CrCO is degraded by the proteasome following its ubiquitination by the SPA1/COP1-dependent E3 ubiquitin ligase complex, thus inhibiting qE gene expression. However, upon exposure to UV-B, during which UVR8 deactivates the E3 ubiquitin ligase through the formation of the UVR8/SPA1/COP1 protein complex, the degradation of CrCO is inhibited and qE gene expression is activated [41] (see Figure 1B).
2.4. CrCO in the Regulation of Photoprotection
Dr. Minagawa’s and Dr. Niyogi’s groups simultaneously discovered that qE activity was almost completely absent in crco mutants, suggesting that CO is essential for the activation of qE-dependent photoprotection in C. reinhardtii [36,41]. The expression of LHCSR1, PSBS, and LHCSR3 was significantly induced by both high light and UV in WT cells but was not detected in the crco-2 mutant [41]. Similar results were obtained in another crco mutant, except that the expression of LHCSR3 was not detected in low light but was detected in high light at a relatively low level comparable to that in the WT under low-light conditions [36]. Moreover, CrCO was shown to function as a transcription factor that regulates the expression of photoprotective genes [36,41]. These results indicate that the expression of photoprotective genes is tightly controlled by CrCO, although an SPA1/CrCO-independent but high light-dependent pathway might exist [36]. Thus, CrCO is thought to play a central role in the regulation of photoprotection. Moreover, the high light and UV-B-signaling pathways might regulate the expression of photoprotective genes by converging on CrCO, although an SPA1/CrCO-independent but high light-dependent pathway might also exist.
Based on recent achievements in elucidating high light and UV-B-signaling pathways, we propose models for high light and UV-B-induced LHCSR1, LHCSR3, and PSBS expression in C. reinhardtii (Figure 1).
2.5. Activation of Photoprotection by Different Wavelengths of Light
As discussed above, NPQ is activated through wavelength-specific light-signaling pathways mediated by PHOT (blue light) and UVR8 (UV-B) photoreceptors in the unicellular green alga C. reinhardtii. What is the biological significance of the activation of photoprotection by different wavelengths of light? It appears that LHCSR1, PSBS, and LHCSR1 have different action spectra: LHCSR1 and PSBS proteins mainly accumulate under UV light, whereas LHCSR3 accumulates under blue and red light. Moreover, UV-illumination at relatively low intensity activates NPQ more rapidly compared to blue or red light at relatively high intensity. This is because UV treatment induces photoprotective gene expression and protein accumulation at a significantly faster rate and with greater magnitude compared to blue or red-light treatment. UVR8 is responsible for the UV-dependent rapid activation of NPQ; this type of photoprotection is indispensable in C. reinhardtii under high-light conditions in the absence of LHCSR3, a photoprotective effector that is primarily activated via blue-light perception [62]. These findings suggest that the UVR8-dependent rapid activation of photoprotection might represent a complementary mechanism under conditions where blue light-induced photoprotection is not sufficient to dissipate excess light energy. In addition, the UVR8-dependent rapid activation of photoprotection may function as “preemptive photoacclimation” prior to the “subsequent photoacclimation” enabled by high light-dependent photoprotection [45,48,62].
3. Questions and Perspectives
Various aspects of photoprotection have been uncovered in recent decades through molecular, biochemical, biophysical, structural, genetic, and physiological studies. Here, we summarized the recent progress in elucidating the signaling pathways from light perception to photoprotective gene expression in the green alga Chlamydomonas reinhardtii. The progress highlighted here clearly demonstrates a link between light perception and photoprotection. We believe that the discovery of such a link will strengthen the connection between the research areas of photobiology and photosynthesis. The next step is to investigate how algae and plants sense color to regulate photoprotection and photosynthesis, a topic that has previously been ignored.
The discovery that several signaling pathways regulate the expression of photoprotective genes in Chlamydomonas (Figure 1) has prompted several questions. For example, what is the intimate link between the UVR8-dependent pathway and the high light-dependent pathway? Is there a switch that balances the UVR8-dependent pathway and PHOT-dependent pathway under natural sunlight conditions? Why (and how) are photoprotective genes induced at a significantly faster rate and with greater magnitude under UV treatment compared to blue or red light treatment? Why is the UVR8-dependent pathway independent of photosynthesis, whereas the PHOT-dependent pathway is dependent on this process? What is the nature of the photosynthetic signal and how is it integrated with the photosynthesis-dependent pathways to regulate LHCSR3 activity? Answering these questions will advance our knowledge of how light signaling regulates photoprotection in Chlamydomonas.
Although several light-signaling pathways for photoprotection have been deciphered in Chlamydomonas, how plants recognize and sense light color to regulate photoprotective gene expression is completely unknown. Moreover, our knowledge about how light signaling regulates photoprotection and the related genetic mechanisms in plants is still relatively limited. We anticipate exciting discoveries about how light signaling regulates qE in plants. It appears that green algae and plants may employ different light-signaling mechanisms in response to excess light. For example, Chlamydomonas contains five photoprotective genes (LHCSR1, LHSCR3.1/3.2, and PSBS1/2), whereas Arabidopsis has only two (PSBS1/2). In addition, in Chlamydomonas, as discussed above, CrCO is a key transcription factor that regulates the expression of photoprotective genes under high-light and UV conditions, and is central to the regulation of photoprotection, whereas CO and its ubiquitination mediate photoperiodic flowering and are crucial for the regulation of photoperiodic flowering in Arabidopsis [63,64]. Furthermore, Chlamydomonas has a distinct UV response compared to land plants: in Chlamydomonas, the UV response is initiated by relatively long wavelengths of UV light, including UV-A/B, whereas Arabidopsis preferentially senses relatively short wavelengths of UV (mainly UV-B/C) [62,65,66,67]. Therefore, it appears that the signaling pathway for photoprotection that depends on the blue-light photoreceptor PHOT in Chlamydomonas is not present in plants; this PHOT-dependent control of photoprotection was apparently lost during the colonization of land [46]. In Arabidopsis, PHOT mediates chloroplast relocation and qM-type NPQ under high-light conditions [19,68], although the existence of a chloroplast movement-dependent component of NPQ is in doubt [21]. However, it is possible that another response that is dependent on the blue-light photoreceptor CRY1 is involved in photoprotection, as CRY1 predominantly mediates the high light-induced expression of Arabidopsis ELI1 and 2, encoding EARLY LIGHT-INDUCED PROTEIN1 (ELIP1) and ELIP2 [69].
Photoprotection represents a promising target for crop improvement. Indeed, enhancing NPQ recovery by upregulating PSBS, violaxanthin de-epoxidase, and zeaxanthin epoxidase genes increased the quantum yield of CO2 assimilation, plant biomass, and yield in tobacco [70]. Enhancing the photoprotective capacity in rice by increasing PSBS protein levels resulted in enhanced biomass and grain yield [71]. Therefore, understanding the signaling pathway from light perception to photoprotective gene expression in plants including crops will be crucial for improving photoprotection by genetically exploiting photoprotection in plants exposed to sunlight and its intrinsic UV-B fraction. This, in turn, will enhance agricultural productivity. We look forward to seeing the light-signaling pathway that regulates NPQ in plants unveiled in the near future.
Author Contributions
D.L. and C.L. conceived and planned this review paper. Y.Z., A.Z. and C.L. prepared and drafted the manuscript. D.L. and C.L. revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Key Research and Development Program of China (grant number 2020YFA0907600) and the National Natural Science Foundation of China (grant numbers 31730102 and 31401035).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Müller, P.; Li, X.-P.; Niyogi, K.K. Non-photochemical quenching. A response to excess light energy. Plant Physiol. 2001, 125, 1558–1566. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V. Non-photochemical chlorophyll fluorescence quenching: Mechanism and effectiveness in protection against photodamage. Plant Physiol. 2016, 170, 1903–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walters, R.G.; Horton, P. Resolution of components of non-photochemical chlorophyll fluorescence quenching in barley leaves. Photosynth. Res. 1991, 27, 121–133. [Google Scholar] [CrossRef]
- Malnoë, A. Photoinhibition or photoprotection of photosynthesis, which one is it? Update on newly termed sustained quenching component, qH. Environ. Exp. Bot. 2018, 154, 123–133. [Google Scholar] [CrossRef]
- Li, X.-P.; Björkman, O.; Shih, C.; Grossman, A.R.; Rosenquist, M.; Jansson, S.; Niyogi, K.K. A pigment-binding protein essential for regulation of photosynthetic light harvesting. Nature 2000, 403, 391–395. [Google Scholar] [CrossRef]
- Sylak-Glassman, E.J.; Malnoë, A.; Re, E.D.; Brooks, M.D.; Fischer, A.L.; Niyogi, K.K.; Fleming, G.R. Distinct roles of the photosystem II protein PsbS and zeaxanthin in the regulation of light harvesting in plants revealed by fluorescence lifetime snapshots. Proc. Natl. Acad. Sci. USA 2014, 111, 17498–17503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horton, P.; Ruban, A.V.; Walters, R.G. Regulation of light harvesting in green plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1996, 47, 655–684. [Google Scholar] [CrossRef]
- Peers, G.; Truong, T.B.; Ostendorf, E.; Busch, A.; Elrad, D.; Grossman, A.R.; Hippler, M.; Niyogi, K.K. An ancient light-harvesting protein is critical for the regulation of algal photosynthesis. Nature 2009, 462, 518–521. [Google Scholar] [CrossRef]
- Niyogi, K.K.; Truong, T.B. Evolution of flexible non-photochemical quenching mechanisms that regulate light harvesting in oxygenic photosynthesis. Curr. Opin. Plant Biol. 2013, 16, 307–314. [Google Scholar] [CrossRef]
- Tokutsu, R.; Minagawa, J. Energy-dissipative supercomplex of photosystem II associated with LHCSR3 in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2013, 110, 10016–10021. [Google Scholar] [CrossRef] [Green Version]
- Ruban, A.V.; Berera, R.; Ilioaia, C.; van Stokkum, I.H.; Kennis, J.T.; Pascal, A.A.; van Amerongen, H.; Robert, B.; Horton, P.; van Grondelle, R. Identification of a mechanism of photoprotective energy dissipation in higher plants. Nature 2007, 450, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Ahn, T.K.; Avenson, T.J.; Ballottari, M.; Cheng, Y.-C.; Niyogi, K.K.; Bassi, R.; Fleming, G.R. Architecture of a charge-transfer state regulating light harvesting in a plant antenna protein. Science 2008, 320, 794–797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brooks, M.D.; Sylak-Glassman, E.J.; Fleming, G.R.; Niyogi, K.K. A thioredoxin-like/β-propeller protein maintains the efficiency of light harvesting in Arabidopsis. Proc. Natl Acad. Sci. USA 2013, 110, E2733–E2740. [Google Scholar] [CrossRef] [Green Version]
- Malnoë, A.; Schultink, A.; Shahrasbi, S.; Rumeau, D.; Havaux, M.; Niyogi, K.K. The plastid lipocalin LCNP is required for sustained photoprotective energy dissipation in Arabidopsis. Plant. Cell 2017, 30, 196–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amstutz, C.L.; Fristedt, R.; Schultink, A.; Merchant, S.S.; Niyogi, K.K.; Malnoë, A. An atypical short-chain dehydrogenase-reductase functions in the relaxation of photoprotective qH in Arabidopsis. Nat. Plants 2020, 6, 154–166. [Google Scholar] [CrossRef]
- Krause, G.H. Photoinhibition of photosynthesis. An evaluation of damaging and protective mechanisms. Physiol. Plant. 1988, 74, 566–574. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Adams, W.W., III. Photoprotection and other responses of plants to high light stress. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1992, 43, 599–626. [Google Scholar] [CrossRef]
- Ruban, A.V.; Horton, P. An investigation of the sustained component of nonphotochemical quenching of chlorophyll fluorescence in isolated chloroplasts and leaves of spinach. Plant. Physiol. 1995, 108, 721–726. [Google Scholar] [CrossRef] [Green Version]
- Cazzaniga, S.; Dall’ Osto, L.; Kong, S.G.; Wada, M.; Bassi, R. Interaction between avoidance of photon absorption, excess energy dissipation and zeaxanthin synthesis against photooxidative stress in Arabidopsis. Plant. J. 2013, 76, 568–579. [Google Scholar] [CrossRef]
- Dall’Osto, L.; Cazzaniga, S.; Wada, M.; Bassi, R. On the origin of a slowly reversible fluorescence decay component in the Arabidopsis npq4 mutant. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 2013022. [Google Scholar] [CrossRef] [Green Version]
- Wilson, S.; Ruban, A.V. Rethinking the influence of chloroplast movements on non-photochemical quenching and photoprotection. Plant. Physiol. 2020, 183, 1213–1223. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll fluorescence and photosynthesis: The basics. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Ruban, A.V.; Johnson, M.P. Dynamics of higher plant photosystem cross-section associated with state transitions. Photosynth. Res. 2009, 99, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Dall’Osto, L.; Caffarri, S.; Bassi, R. A mechanism of nonphotochemical energy dissipation, independent from PsbS, revealed by a conformational change in the antenna protein CP26. Plant. Cell 2005, 17, 1217–1232. [Google Scholar] [CrossRef] [Green Version]
- Betterle, N.; Ballottari, M.; Hienerwadel, R.; Dall’Osto, L.; Bassi, R. Dynamics of zeaxanthin binding to the photosystem II monomeric antenna protein Lhcb6 (CP24) and modulation of its photoprotection properties. Arch. Biochem. Biophys. 2010, 504, 67–77. [Google Scholar] [CrossRef]
- Nilkens, M.; Kress, E.; Lambrev, P.; Miloslavina, Y.; Müller, M.; Holzwarth, A.R.; Jahns, P. Identification of a slowly inducible zeaxanthin-dependent component of non-photochemical quenching of chlorophyll fluorescence generated under steady-state conditions in Arabidopsis. Biochim. Biophys. Acta 2010, 1797, 466–475. [Google Scholar] [CrossRef] [Green Version]
- Jahns, P.; Holzwarth, A.R. The role of the xanthophyll cycle and of lutein in photoprotection of photosystem II. Biochim. Biophys. Acta 2012, 1817, 182–193. [Google Scholar] [CrossRef] [Green Version]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light stress and photoprotection in Chlamydomonas reinhardtii. Plant. J. 2015, 82, 449–465. [Google Scholar] [CrossRef]
- Demmig-Adams, B.; Stewart, J.J.; Adams, W.W.W., III. Less photoprotection can be good in some genetic and environmental contexts. Bioch. J. 2019, 476, 2017–2029. [Google Scholar] [CrossRef]
- Murchie, E.H.; Ruban, A.V. Dynamic non-photochemical quenching in plants: From molecular mechanism to productivity. Plant. J. 2020, 101, 885–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruban, A.V.; Wilson, S. The mechanism of non-photochemical quenching in plants: Localization and driving forces. Plant. Cell Physiol. 2021, 62, 1063–1072. [Google Scholar] [CrossRef]
- Giovagnetti, V.; Ruban, A.V. The evolution of the photoprotective antenna proteins in oxygenic photosynthetic eukaryotes. Biochem. Soc. Trans. 2018, 46, 1263–1277. [Google Scholar] [CrossRef] [PubMed]
- Pinnola, A.; Bassi, R. Molecular mechanisms involved in plant photoprotection. Biochem. Soc. Trans. 2018, 46, 467–482. [Google Scholar] [CrossRef]
- Pinnola, A. The rise and fall of Light-Harvesting Complex Stress-Related proteins as photoprotection agents during evolution. J. Exp. Bot. 2019, 70, 5527–5535. [Google Scholar] [CrossRef] [PubMed]
- Allorent, G.; Petroutsos, D. Photoreceptor-dependent regulation of photoprotection. Curr. Opin. Plant. Biol. 2017, 37, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Gabilly, S.T.; Baker, C.R.; Wakao, S.; Crisanto, T.; Guan, K.; Bi, K.; Guiet, E.; Guadagno, C.R.; Niyogia, K.K. Regulation of photoprotection gene expression in Chlamydomonas by a putative E3 ubiquitin ligase complex and a homolog of CONSTANS. Proc. Natl. Acad. Sci. USA 2019, 116, 17556–17562. [Google Scholar] [CrossRef] [Green Version]
- Alboresi, A.; Caffarri, S.; Nogue, F.; Bassi, R.; Morosinotto, T. In silico and biochemical analysis of Physcomitrella patens photosynthetic antenna: Identification of subunits which evolved upon land adaptation. PLoS ONE 2008, 3, e2033. [Google Scholar] [CrossRef] [Green Version]
- Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.; Shapiro, H.; Nishiyama, T.; Perroud, P.F.; Rensing, S.A.; Lindquist, E.A.; et al. The genome of the moss Physcomitrella patens reveals evolutionary insights into the conquest of land by plants. Science 2008, 319, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Ledford, H.K.; Baroli, I.; Shin, J.W.; Fischer, B.B.; Eggen, R.I.L.; Niyogi, K.K. Comparative profiling of lipid-soluble antioxidants and transcripts reveals two phases of photo-oxidative stress in a xanthophyll-deficient mutant of Chlamydomonas reinhardtii. Mol. Genet. Genom. 2004, 272, 470–479. [Google Scholar] [CrossRef]
- Maruyama, S.; Tokutsu, R.; Minagawa, J. Transcriptional regulation of the stress-responsive light harvesting complex genes in Chlamydomonas reinhardtii. Plant. Cell Physiol. 2014, 55, 1304–1310. [Google Scholar] [CrossRef]
- Tokutsu, R.; Fujimura-Kamada, K.; Matsuo, T.; Yamasaki, T.; Minagawa, J. The CONSTANS flowering complex controls the protective response of photosynthesis in the green alga Chlamydomonas. Nat. Commun. 2019, 10, 4099. [Google Scholar] [CrossRef]
- Petroutsos, D.; Busch, A.; Janssen, I.; Trompelt, K.; Bergner, S.V.; Weinl, S.; Holtkamp, M.; Karst, U.; Kudla, J.; Hippler, M. The chloroplast calcium sensor CAS is required for photoacclimation in Chlamydomonas reinhardtii. Plant. Cell 2011, 23, 2950–2963. [Google Scholar] [CrossRef] [Green Version]
- Yamano, T.; Miura, K.; Fukuzawa, H. Expression analysis of genes associated with the induction of the carbon-concentrating mechanism in Chlamydomonas reinhardtii. Plant. Physiol. 2008, 147, 340–354. [Google Scholar] [CrossRef] [Green Version]
- Moseley, J.L.; Chang, C.W.; Grossman, A.R. Genome-based approaches to understanding phosphorus deprivation responses and PSR1 control in Chlamydomonas reinhardtii. Eukaryot Cell 2006, 5, 26–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilbrook, K.; Dubois, M.; Crocco, C.D.; Yin, R.; Chappuis, R.; Allorent, G.; Schmid-Siegert, E.; Goldschmidt-Clermont, M.; Ulm, R. UV-B perception and acclimation in Chlamydomonas reinhardtii. Plant. Cell 2016, 28, 966–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petroutsos, D.; Tokutsu, R.; Maruyama, S.; Flori, S.; Greiner, A.; Magneschi, L.; Cusant, L.; Kottke, T.; Mittag, M.; Hegemann, P.; et al. A blue-light photoreceptor mediates the feedback regulation of photosynthesis. Nature 2016, 537, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Aihara, Y.; Fujimura-Kamada, K.; Yamasaki, T.; Minagawa, J. Algal photoprotection is regulated by the E3 ligase CUL4–DDB1DET1. Nat. Plants 2019, 5, 34–40. [Google Scholar] [CrossRef]
- Allorent, G.; Lefebvre-Legendre, L.; Chappuis, R.; Kuntz, M.; Truong, T.B.; Niyogi, K.K.; Ulm, R.; Goldschmidt-Clermont, M. UV-B photoreceptor-mediated protection of the photosynthetic machinery in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 14864–14869. [Google Scholar] [CrossRef] [Green Version]
- Correa-Galvis, V.; Redekop, P.; Guan, K.; Griess, K.; Truong, T.B.; Wakao, S.; Niyogi, K.K.; Jahns, P. Photosystem II subunit PsbS is involved in the induction of LHCSR protein-dependent energy dissipation in Chlamydomonas reinhardtii. J. Biol. Chem. 2016, 291, 17478–17487. [Google Scholar] [CrossRef] [Green Version]
- Tibiletti, T.; Auroy, P.; Peltier, G.; Caffarri, S. Chlamydomonas reinhardtii PsbS protein is functional and accumulates rapidly and transiently under high light. Plant. Physiol. 2016, 171, 2717–2730. [Google Scholar] [CrossRef] [Green Version]
- Kirilovsky, D.; Kerfeld, C.A. The orange carotenoid protein: A blue-green light photoactive protein. Photochem. Photobiol. Sci. 2013, 12, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Kirilovsky, D.; Kerfeld, C.A. Cyanobacterial photoprotection by the orange carotenoid protein. Nat. Plants 2016, 2, 16180. [Google Scholar] [CrossRef] [Green Version]
- Coesel, S.; Mangogna, M.; Ishikawa, T.; Heijde, M.; Rogato, A.; Finazzi, G.; Todo, T.; Bowler, C.; Falciatore, A. Diatom PtCPF1 is a new cryptochrome/photolyase family member with DNA repair and transcription regulation activity. EMBO Rep. 2009, 10, 655–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, B.S.; Jungandreas, A.; Jakob, T.; Weisheit, W.; Mittag, M.; Wilhelm, C. Blue light is essential for high light acclimation and photoprotection in the diatom Phaeodactylum tricornutum. J. Exp. Bot. 2013, 64, 483–493. [Google Scholar] [CrossRef] [Green Version]
- Juhas, M.; von Zadow, A.; Spexard, M.; Schmidt, M.; Kottke, T.; Büchel, C. A novel cryptochrome in the diatom Phaeodactylum tricornutum influences the regulation of light-harvesting protein levels. FEBS J. 2014, 281, 2299–2311. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, F. Blue-light-regulated transcription factor Aureochrome, in photosynthetic stramenopiles. J. Plant. Res. 2016, 129, 189–197. [Google Scholar] [CrossRef]
- Serif, M.; Lepetit, B.; Weißert, K.; Kroth, P.G.; Rio Bartulos, C. A fast and reliable strategy to generate TALEN-mediated gene knockouts in the diatom Phaeodactylum tricornutum. Algal Res. 2017, 23, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Kataria, S.; Jajoo, A.; Guruprasad, K.N. Impact of increasing Ultraviolet-B (UV-B) radiation on photosynthetic processes. J. Photochem. Photobiol. B 2014, 137, 55–66. [Google Scholar] [CrossRef]
- Takahashi, S.; Milward, S.E.; Yamori, W.; Evans, J.R.; Hillier, W.; Badger, M.R. The solar action spectrum of photosystem II damage. Plant. Physiol. 2010, 153, 988–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zavafer, A.; Chow, W.S.; Cheah, M.H. The action spectrum of photosystem II photoinactivation in visible light. J. Photochem. Photobiol. B 2015, 152, 247–260. [Google Scholar] [CrossRef]
- Tokutsu, R.; Fujimura-Kamada, K.; Yamasaki, T.; Matsuo, T.; Minagawa, J. Isolation of photoprotective signal transduction mutants by systematic bioluminescence screening in Chlamydomonas reinhardtii. Sci. Rep. 2019, 9, 2820. [Google Scholar] [CrossRef] [Green Version]
- Tokutsu, R.; Fujimura-Kamada, K.; Yamasaki, T.; Okajima, K.; Minagawa, J. UV-A/B radiation rapidly activates photoprotective mechanisms in Chlamydomonas reinhardtii. Plant. Physiol. 2021, 185, 1894–1902. [Google Scholar] [CrossRef] [PubMed]
- Suárez-López, P.; Wheatley, K.; Robson, F.; Onouchi, H.; Valverde, F.; Coupland, G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature 2001, 410, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.H.; Shim, J.S.; Kinmonth-Schultz, H.A.; Imaizumi, T. Photoperiodic flowering: Time measurement mechanisms in leaves. Annu. Rev. Plant. Biol. 2015, 66, 441–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rizzini, L.; Favory, J.J.; Cloix, C.; Faggionato, D.; O’Hara, A.; Kaiserli, E.; Baumeister, R.; Schäfer, E.; Nagy, F.; Jenkins, G.I.; et al. Perception of UV-B by the Arabidopsis UVR8 protein. Science 2011, 332, 103–106. [Google Scholar] [CrossRef] [Green Version]
- Christie, J.M.; Arvai, A.S.; Baxter, K.J.; Heilmann, M.; Pratt, A.J.; O’Hara, A.; Kelly, S.M.; Hothorn, M.; Smith, B.O.; Hitomi, K.; et al. Plant UVR8 photoreceptor senses UV-B by tryptophan-mediated disruption of cross-dimer salt bridges. Science 2012, 335, 1492–1496. [Google Scholar] [CrossRef] [Green Version]
- Wu, D.; Hu, Q.; Yan, Z.; Chen, W.; Yan, C.; Huang, X.; Zhang, J.; Yang, P.; Deng, H.; Wang, J.; et al. Structural basis of ultraviolet-B perception by UVR8. Nature 2012, 484, 214–219. [Google Scholar]
- Sakai, T.; Kagawa, T.; Kasahara, M.; Swartz, T.E.; Christie, J.M.; Briggs, W.R.; Wada, M.; Okada, K. Arabidopsis nph1 and npl1: Blue light receptors that mediate both phototropism and chloroplast relocation. Proc. Natl. Acad. Sci. USA 2001, 98, 6969–6974. [Google Scholar] [CrossRef] [Green Version]
- Kleine, T.; Kindgren, P.; Benedict, K.; Hendrickson, L.; Strand, A. Genome-wide gene expression analysis reveals a critical role for CRYPTOCHROME1 in the response of Arabidopsis to high irradiance. Plant. Physiol. 2007, 144, 1391–1406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Hubbart, S.; Smillie Ian, R.A.; Heatley, M.; Swarup, R.; Foo, C.C.; Zhao, L.; Murchie, E.H. Enhanced thylakoid photoprotection can increase yield and canopy radiation use efficiency in rice. Commun. Biol. 2018, 1, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
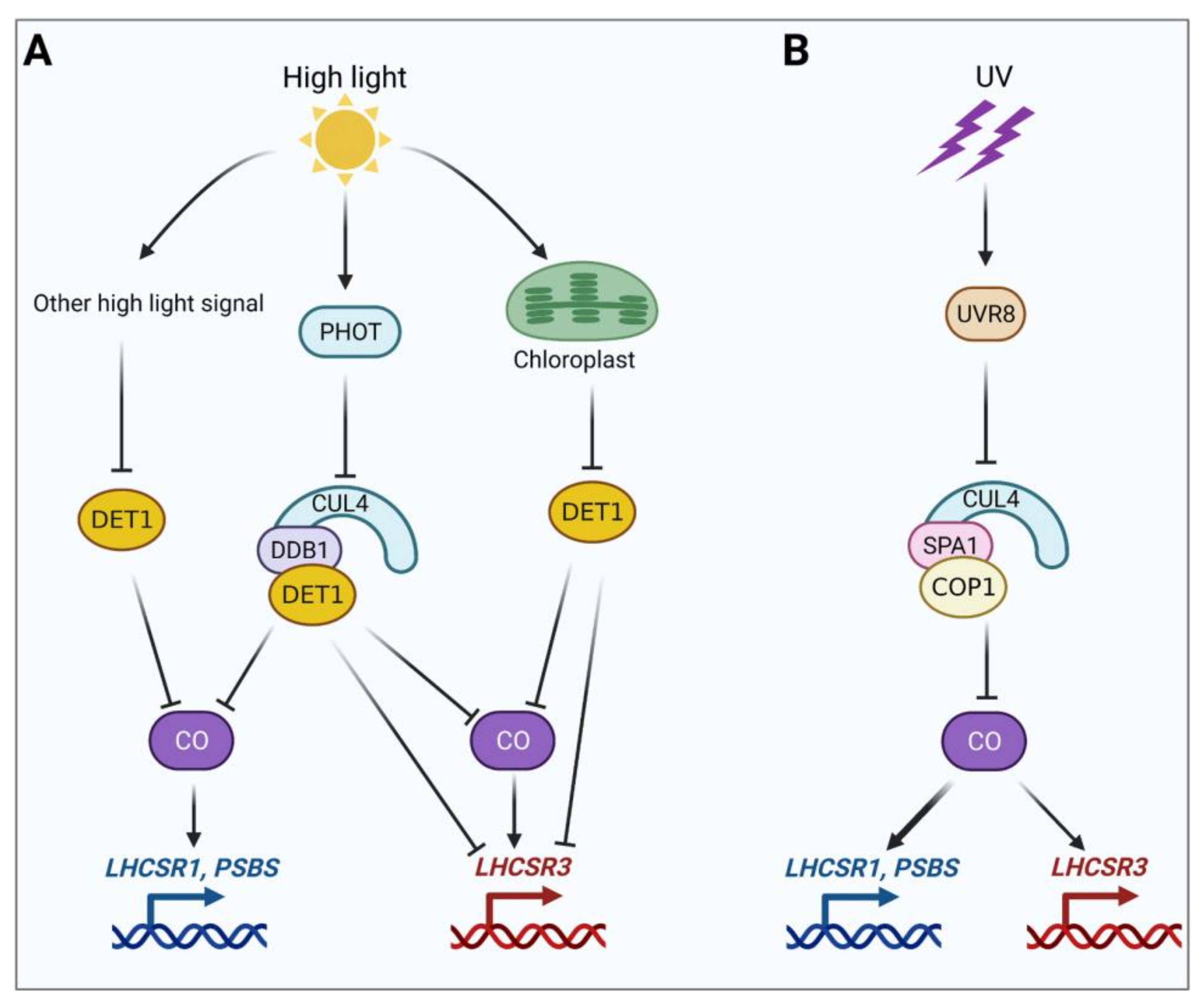
Figure 1.
Models for the induction of LHCSR1, LHCSR3, and PSBS expression by high light and UV-B in C. reinhardtii. (A) Induction of LHCSR1, LHCSR3, and PSBS expression by high light via the putative E3 ligase complex CUL4-DDB1DET1 and other DET1-dependent high-light signaling pathways. (B) Induction of LHCSR1, LHCSR3, and PSBS expression by UV-B via the SPA1/COP1-dependent E3 ligase complex. See the main text for a discussion of the related signaling pathways. This figure was created with the help of the Biorender.
Figure 1.
Models for the induction of LHCSR1, LHCSR3, and PSBS expression by high light and UV-B in C. reinhardtii. (A) Induction of LHCSR1, LHCSR3, and PSBS expression by high light via the putative E3 ligase complex CUL4-DDB1DET1 and other DET1-dependent high-light signaling pathways. (B) Induction of LHCSR1, LHCSR3, and PSBS expression by UV-B via the SPA1/COP1-dependent E3 ligase complex. See the main text for a discussion of the related signaling pathways. This figure was created with the help of the Biorender.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lu, D.; Zhang, Y.; Zhang, A.; Lu, C. Non-Photochemical Quenching: From Light Perception to Photoprotective Gene Expression. Int. J. Mol. Sci. 2022, 23, 687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020687
AMA Style
Lu D, Zhang Y, Zhang A, Lu C. Non-Photochemical Quenching: From Light Perception to Photoprotective Gene Expression. International Journal of Molecular Sciences. 2022; 23(2):687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020687
Chicago/Turabian StyleLu, Dandan, Yi Zhang, Aihong Zhang, and Congming Lu. 2022. "Non-Photochemical Quenching: From Light Perception to Photoprotective Gene Expression" International Journal of Molecular Sciences 23, no. 2: 687. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020687
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.