
Properties and Potential Antiproliferative Activity of Thrombin-Binding Aptamer (TBA) Derivatives with One or Two Additional G-Tetrads

 ,
,  , , , , , , and
, , , , , , and 
Abstract
:1. Introduction
2. Results
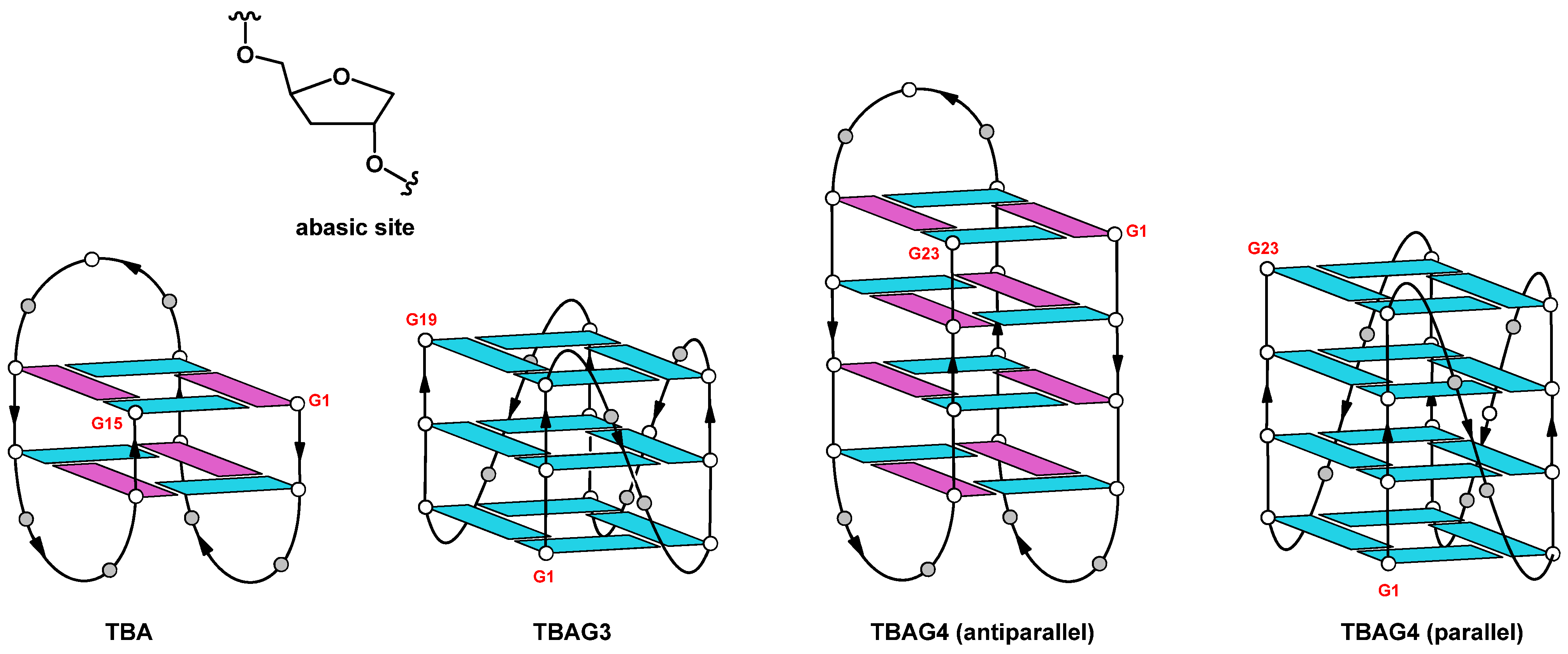
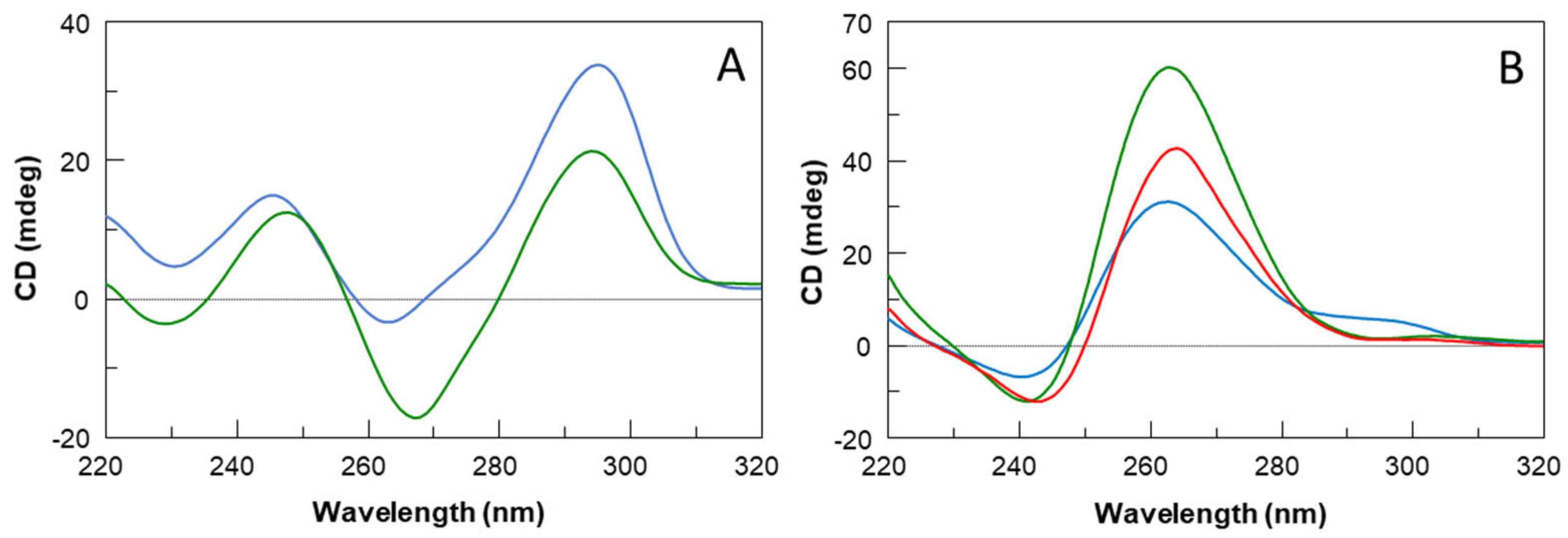
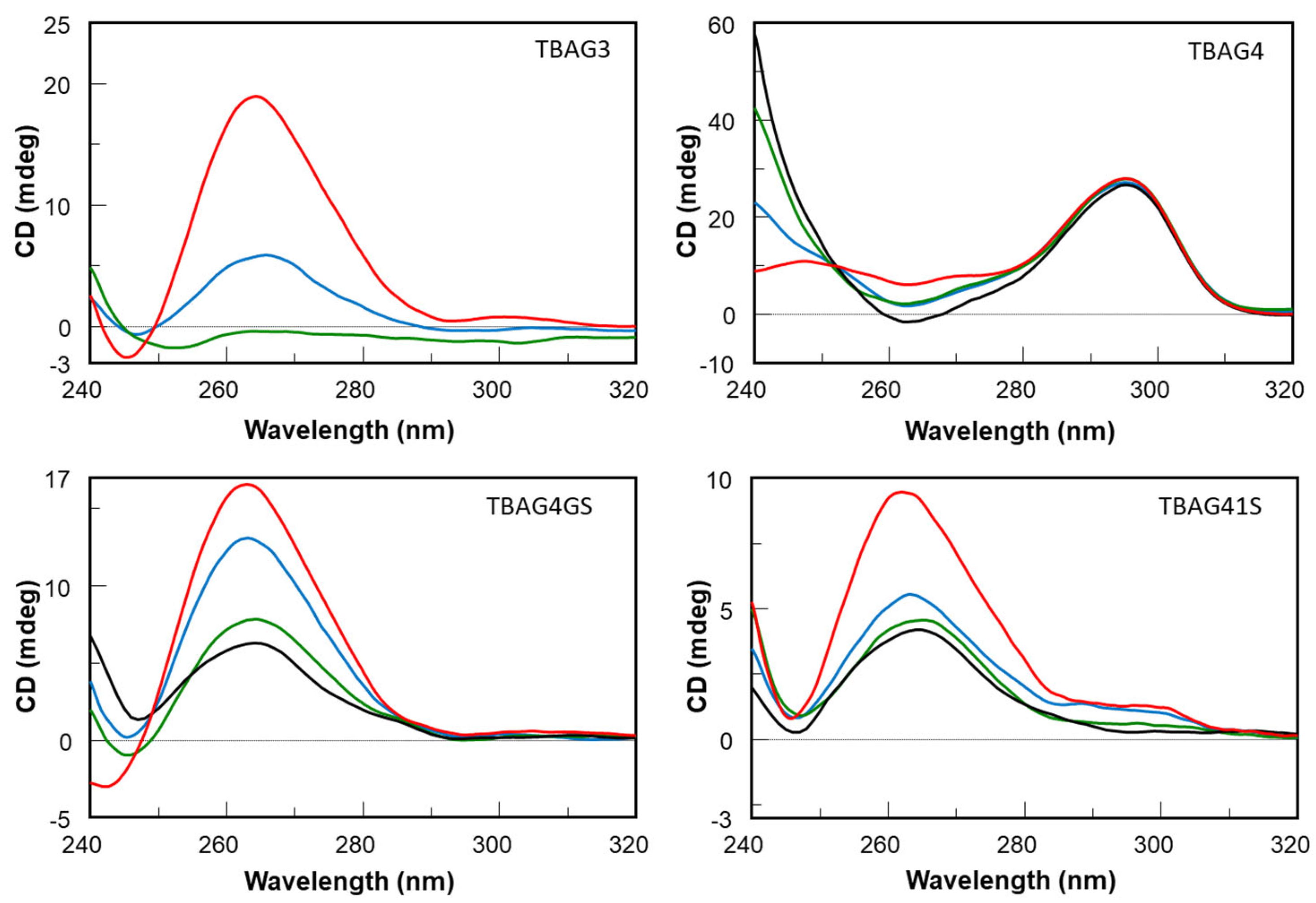
2.1. Structural Insights of the Investigated Sequences
2.2. Nuclease Stability Assay
2.3. Anticoagulant Activity
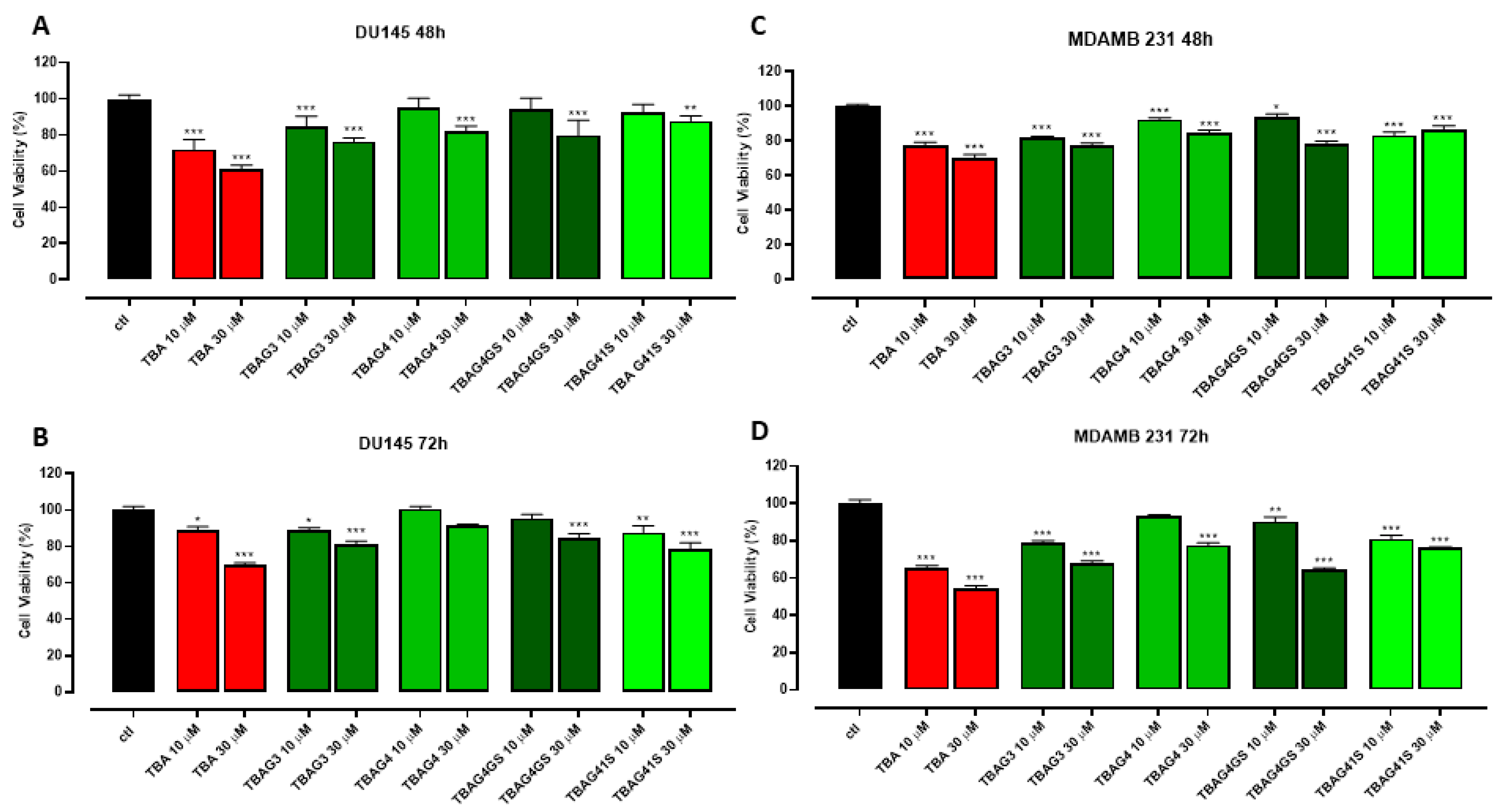
2.4. Antiproliferative Activity
2.5. Nucleolin Binding to G-Quadruplexes
3. Discussion
4. Materials and Methods
4.1. Oligonucleotide Synthesis and Purification
4.2. CD Spectroscopy
4.3. Gel Electrophoresis
4.4. Nuclease Stability Assay
4.5. Prothrombin Time (PT) Assay
4.6. MTT Assay
4.7. Surface Plasmon Resonance (SPR)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xu, Y.; Jiang, X.; Zhou, Y.; Ma, M.; Wang, M.; Ying, B. Systematic Evolution of Ligands by Exponential Enrichment Technologies and Aptamer-Based Applications: Recent Progress and Challenges in Precision Medicine of Infectious Diseases. Front. Bioeng. Biotechnol. 2021, 9, 704077. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N.; Chaires, J.B.; Gray, R.D.; Trent, J.O. Stability and Kinetics of G-Quadruplex Structures. Nucleic Acids Res. 2008, 36, 5482–5515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petraccone, L.; Erra, E.; Esposito, V.; Randazzo, A.; Galeone, A.; Barone, G.; Giancola, C. Biophysical Properties of Quadruple Helices of Modified Human Telomeric DNA. Biopolymers 2005, 77, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Roxo, C.; Kotkowiak, W.; Pasternak, A. G-Quadruplex-Forming Aptamers—Characteristics, Applications, and Perspectives. Molecules 2019, 24, 3781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riccardi, C.; Napolitano, E.; Platella, C.; Musumeci, D.; Montesarchio, D. G-Quadruplex-Based Aptamers Targeting Human Thrombin: Discovery, Chemical Modifications and Antithrombotic Effects. Pharmacol. Ther. 2021, 217, 107649. [Google Scholar] [CrossRef]
- Dapić, V.; Abdomerović, V.; Marrington, R.; Peberdy, J.; Rodger, A.; Trent, J.O.; Bates, P.J. Biophysical and Biological Properties of Quadruplex Oligodeoxyribonucleotides. Nucleic Acids Res. 2003, 31, 2097–2107. [Google Scholar] [CrossRef] [Green Version]
- Mayer, G.; Rohrbach, F.; Pötzsch, B.; Müller, J. Aptamer-Based Modulation of Blood Coagulation. Hamostaseologie 2011, 31, 258–263. [Google Scholar] [CrossRef]
- Scuotto, M.; Rivieccio, E.; Varone, A.; Corda, D.; Bucci, M.; Vellecco, V.; Cirino, G.; Virgilio, A.; Esposito, V.; Galeone, A.; et al. Site Specific Replacements of a Single Loop Nucleoside with a Dibenzyl Linker May Switch the Activity of TBA from Anticoagulant to Antiproliferative. Nucleic Acids Res. 2015, 43, 7702–7716. [Google Scholar] [CrossRef] [Green Version]
- Esposito, V.; Russo, A.; Amato, T.; Vellecco, V.; Bucci, M.; Mayol, L.; Russo, G.; Virgilio, A.; Galeone, A. The “Janus Face” of the Thrombin Binding Aptamer: Investigating the Anticoagulant and Antiproliferative Properties through Straightforward Chemical Modifications. Bioorg. Chem. 2018, 76, 202–209. [Google Scholar] [CrossRef]
- Kotkowiak, W.; Lisowiec-Wachnicka, J.; Grynda, J.; Kierzek, R.; Wengel, J.; Pasternak, A. Thermodynamic, Anticoagulant, and Antiproliferative Properties of Thrombin Binding Aptamer Containing Novel UNA Derivative. Mol. Ther. Nucleic Acids 2018, 10, 304–316. [Google Scholar] [CrossRef]
- Esposito, V.; Russo, A.; Amato, T.; Varra, M.; Vellecco, V.; Bucci, M.; Russo, G.; Virgilio, A.; Galeone, A. Backbone Modified TBA Analogues Endowed with Antiproliferative Activity. BBA-Gen. Subj. 2017, 1861, 1213–1221. [Google Scholar] [CrossRef] [Green Version]
- Esposito, V.; Russo, A.; Vellecco, V.; Bucci, M.; Russo, G.; Mayol, L.; Virgilio, A.; Galeone, A. Thrombin Binding Aptamer Analogues Containing Inversion of Polarity Sites Endowed with Antiproliferative and Anti-Motility Properties against Calu-6 Cells. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 2645–2650. [Google Scholar] [CrossRef]
- Virgilio, A.; Esposito, V.; Pecoraro, A.; Russo, A.; Vellecco, V.; Pepe, A.; Bucci, M.; Russo, G.; Galeone, A. Structural Properties and Anticoagulant/Cytotoxic Activities of Heterochiral Enantiomeric Thrombin Binding Aptamer (TBA) Derivatives. Nucleic Acids Res. 2020, 48, 12556–12565. [Google Scholar] [CrossRef]
- Zhang, N.; Bing, T.; Liu, X.; Qi, C.; Shen, L.; Wang, L.; Shangguan, D. Cytotoxicity of Guanine-Based Degradation Products Contributes to the Antiproliferative Activity of Guanine-Rich Oligonucleotides. Chem. Sci. 2015, 6, 3831–3838. [Google Scholar] [CrossRef]
- Wang, J.; Bing, T.; Zhang, N.; Shen, L.; He, J.; Liu, X.; Wang, L.; Shangguan, D. The Mechanism of the Selective Antiproliferation Effect of Guanine-Based Biomolecules and Its Compensation. ACS Chem. Biol. 2019, 14, 1164–1173. [Google Scholar] [CrossRef]
- Zhou, S.; Xiao, W.; Pan, X.; Zhu, M.; Yang, Z.; Zhang, F.; Zheng, C. Thrombin Promotes Proliferation of Human Lung Fibroblasts via Protease Activated Receptor-1-Dependent and NF-ΚB-Independent Pathways. Cell Biol. Int. 2014, 38, 747–756. [Google Scholar] [CrossRef]
- Reddel, C.; Tan, C.; Chen, V. Thrombin Generation and Cancer: Contributors and Consequences. Cancers 2019, 11, 100. [Google Scholar] [CrossRef] [Green Version]
- Bates, P.J.; Reyes-Reyes, E.M.; Malik, M.T.; Murphy, E.M.; O’Toole, M.G.; Trent, J.O. G-Quadruplex Oligonucleotide AS1411 as a Cancer-Targeting Agent: Uses and Mechanisms. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 1414–1428. [Google Scholar] [CrossRef]
- Avino, A.; Fabrega, C.; Tintore, M.; Eritja, R. Thrombin Binding Aptamer, More than a Simple Aptamer: Chemically Modified Derivatives and Biomedical Applications. Curr. Pharm. Des. 2012, 18, 2036–2047. [Google Scholar] [CrossRef] [Green Version]
- Han, J.H.; Kim, J.H.; Kim, S.K.; Jang, Y.J. Conformational Change of a G-Quadruplex under Molecular Crowding Conditions. J. Biomol. Struct. Dyn. 2020, 38, 2575–2581. [Google Scholar] [CrossRef]
- Svetlova, J.; Sardushkin, M.; Kolganova, N.; Timofeev, E. Recognition Interface of the Thrombin Binding Aptamer Requires Antiparallel Topology of the Quadruplex Core. Biomolecules 2021, 11, 1332. [Google Scholar] [CrossRef] [PubMed]
- Randazzo, A.; Galeone, A.; Esposito, V.; Varra, M.; Mayol, L. Interaction of Distamycin a and Netropsin with Quadruplex and Duplex Structures: A Comparative 1H-NMR Study. Nucleosides Nucleotides Nucleic Acids 2002, 21, 535–545. [Google Scholar] [CrossRef] [PubMed]
- Hahn, J.; Wickham, S.F.J.; Shih, W.M.; Perrault, S.D. Addressing the Instability of DNA Nanostructures in Tissue Culture. ACS Nano 2014, 8, 8765–8775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrone, R.; Butovskaya, E.; Lago, S.; Garzino-Demo, A.; Pannecouque, C.; Palù, G.; Richter, S.N. The G-Quadruplex-Forming Aptamer AS1411 Potently Inhibits HIV-1 Attachment to the Host Cell. Int. J. Antimicrob. Agents 2016, 47, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Virgilio, A.; Benigno, D.; Pecoraro, A.; Russo, A.; Russo, G.; Esposito, V.; Galeone, A. Exploring New Potential Anticancer Activities of the G-Quadruplexes Formed by [(GTG2T(G3T)3] and Its Derivatives with an Abasic Site Replacing Single Thymidine. Int. J. Mol. Sci. 2021, 22, 7040. [Google Scholar] [CrossRef]
- Peng, C.G.; Damha, M.J. G-Quadruplex Induced Stabilization by 2′-Deoxy-2′-Fluoro-d-Arabinonucleic Acids (2′F-ANA). Nucleic Acids Res. 2007, 35, 4977–4988. [Google Scholar] [CrossRef]
- Virgilio, A.; Pecoraro, A.; Benigno, D.; Russo, A.; Russo, G.; Esposito, V.; Galeone, A. Antiproliferative Effects of the Aptamer d(GGGT)4 and Its Analogues with an Abasic-Site Mimic Loop on Different Cancer Cells. Int. J. Mol. Sci. 2022, 23, 5952. [Google Scholar] [CrossRef]
- Ogloblina, A.M.; Khristich, A.N.; Karpechenko, N.Y.; Semina, S.E.; Belitsky, G.A.; Dolinnaya, N.G.; Yakubovskaya, M.G. Multi-Targeted Effects of G4-Aptamers and Their Antiproliferative Activity against Cancer Cells. Biochimie 2018, 145, 163–173. [Google Scholar] [CrossRef]
- Lago, S.; Tosoni, E.; Nadai, M.; Palumbo, M.; Richter, S.N. The Cellular Protein Nucleolin Preferentially Binds Long-Looped G-Quadruplex Nucleic Acids. BBA-Gen. Subj. 2016, 1861, 1371–1381. [Google Scholar] [CrossRef]
- Métifiot, M.; Amrane, S.; Mergny, J.L.; Andreola, M.L. Anticancer Molecule AS1411 Exhibits Low Nanomolar Antiviral Activity against HIV-1. Biochimie 2015, 118, 173–175. [Google Scholar] [CrossRef]
- Carella, A.; Roviello, V.; Iannitti, R.; Palumbo, R.; la Manna, S.; Marasco, D.; Trifuoggi, M.; Diana, R.; Roviello, G.N. Evaluating the Biological Properties of Synthetic 4-Nitrophenyl Functionalized Benzofuran Derivatives with Telomeric DNA Binding and Antiproliferative Activities. Int. J. Biol. Macromol. 2019, 121, 77–88. [Google Scholar] [CrossRef]
- Burra, S.; Marasco, D.; Malfatti, M.C.; Antoniali, G.; Virgilio, A.; Esposito, V.; Demple, B.; Galeone, A.; Tell, G. Human AP-Endonuclease (Ape1) Activity on Telomeric G4 Structures Is Modulated by Acetylatable Lysine Residues in the N-Terminal Sequence. DNA Repair 2019, 73, 129–143. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NAME | SEQUENCE | Tm (°C) ± 1 | KD (µM) |
|---|---|---|---|
| TBA | GGTTGGTGTGGTTGG | 50 | 62.5 |
| TBAG3 | GGGTTGGGTGTGGGTTGGG | 77 | 1293 |
| TBAG4 | GGGGTTGGGGTGTGGGGTTGGGG | >90 | 13.5 |
| TBAG41S | GGGGSSGGGGTGTGGGGTTGGGG | >70 | 41.5 |
| TBAG4GS | GGGGTTGGGGSSSGGGGTTGGGG | 85 | 32.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benigno, D.; Virgilio, A.; Bello, I.; La Manna, S.; Vellecco, V.; Bucci, M.; Marasco, D.; Panza, E.; Esposito, V.; Galeone, A. Properties and Potential Antiproliferative Activity of Thrombin-Binding Aptamer (TBA) Derivatives with One or Two Additional G-Tetrads. Int. J. Mol. Sci. 2022, 23, 14921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314921
Benigno D, Virgilio A, Bello I, La Manna S, Vellecco V, Bucci M, Marasco D, Panza E, Esposito V, Galeone A. Properties and Potential Antiproliferative Activity of Thrombin-Binding Aptamer (TBA) Derivatives with One or Two Additional G-Tetrads. International Journal of Molecular Sciences. 2022; 23(23):14921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314921
Chicago/Turabian StyleBenigno, Daniela, Antonella Virgilio, Ivana Bello, Sara La Manna, Valentina Vellecco, Mariarosaria Bucci, Daniela Marasco, Elisabetta Panza, Veronica Esposito, and Aldo Galeone. 2022. "Properties and Potential Antiproliferative Activity of Thrombin-Binding Aptamer (TBA) Derivatives with One or Two Additional G-Tetrads" International Journal of Molecular Sciences 23, no. 23: 14921. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232314921