
CRISPR/Cas9-Mediated Editing of AGAMOUS-like Genes Results in a Late-Bolting Phenotype in Chinese Cabbage (Brassica rapa ssp. pekinensis)

Abstract
:1. Introduction
2. Results
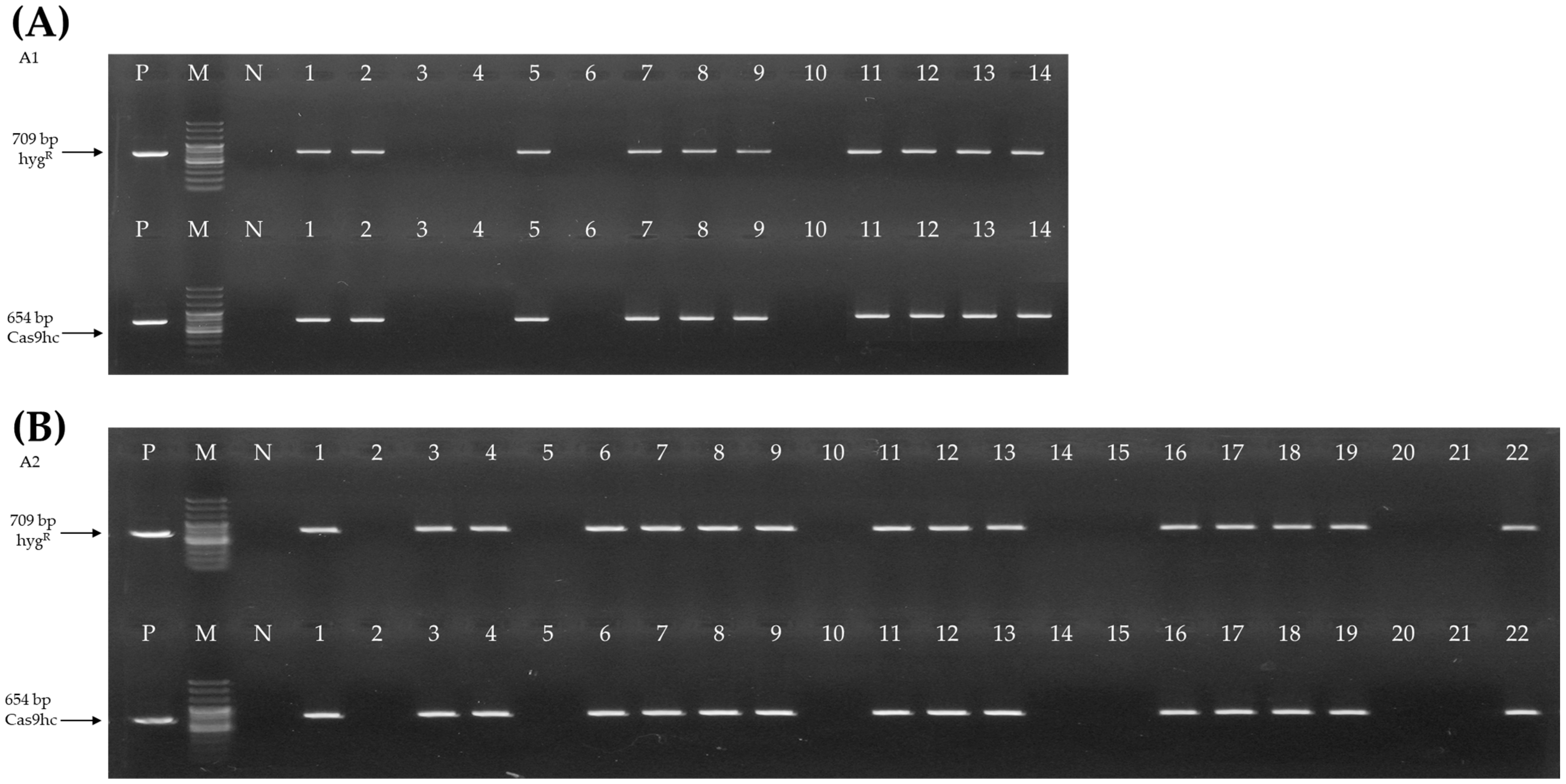
2.1. Vector Construction and Agrobacterium-Mediated Transformation of Chinese Cabbage
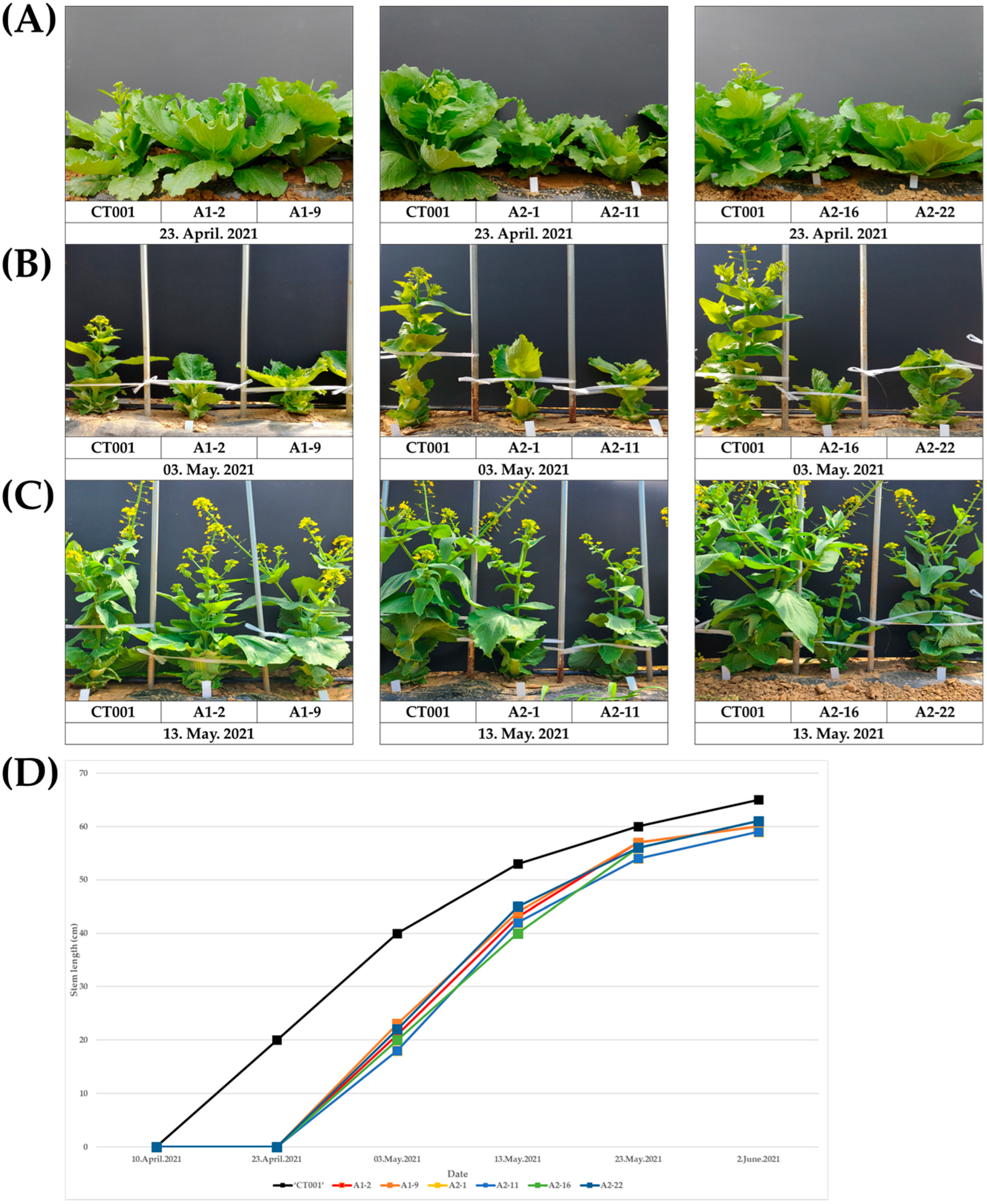
2.2. Selection and Bolting Time Analysis of E0 AGL-Edited Chinese Cabbage Lines
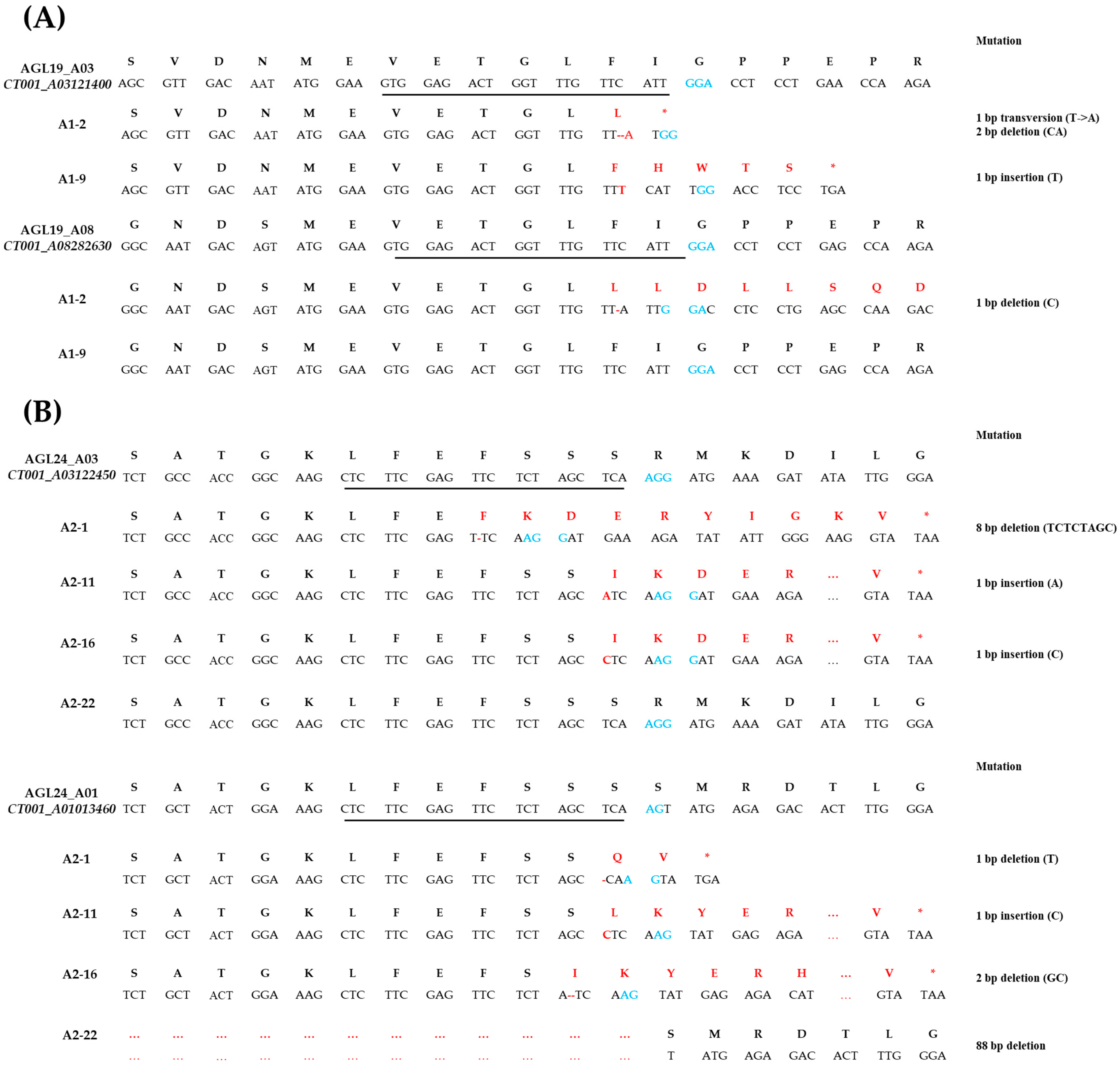
2.3. Mutagenesis of AGL Induced by CRISPR/Cas9 in E0 AGL-Edited Lines
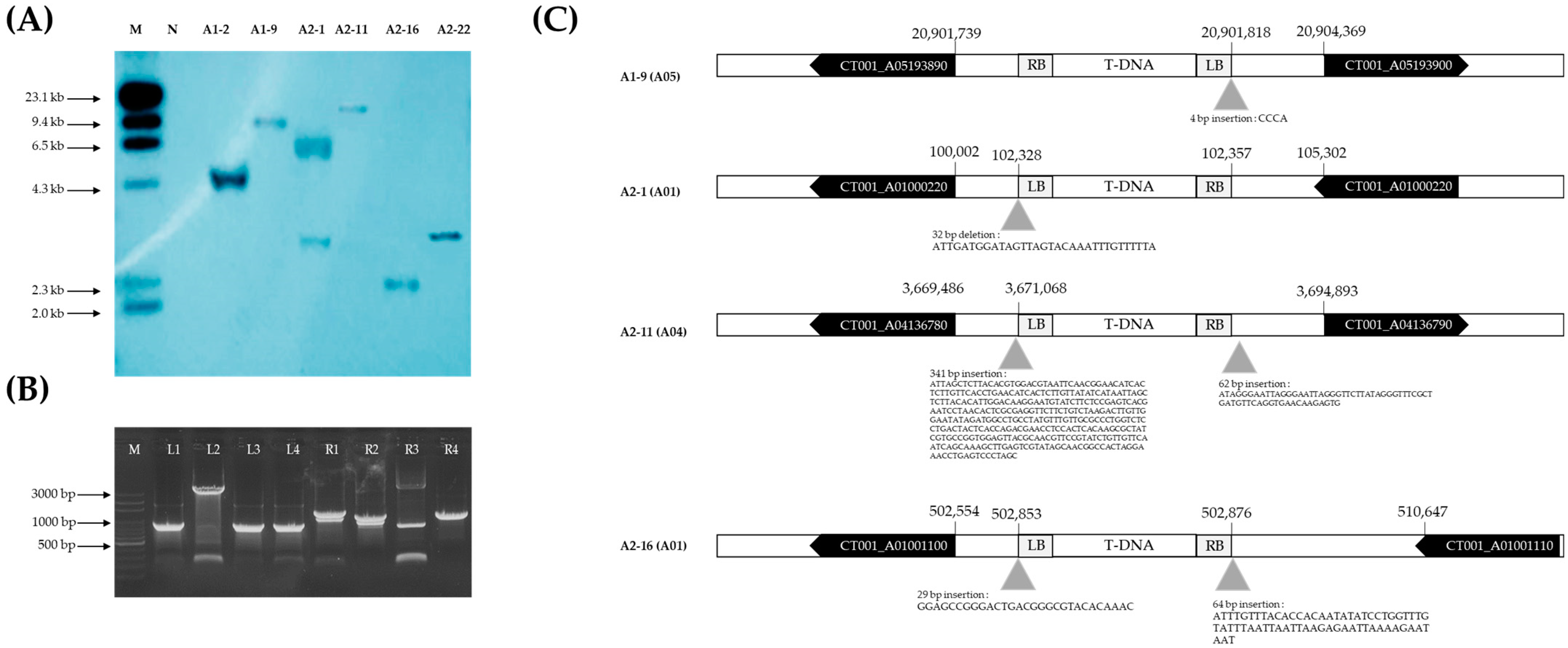
2.4. T-DNA Copy Number and Flanking Site Sequence Analysis of Late-Bolting E0 AGL-Edited Lines
2.5. Mutagenesis Inheritance Analysis in T-DNA-Free E1 AGL-Edited Lines
3. Discussion
4. Materials and Methods
4.1. sgRNA Design and Construction of Gene-Editing Vectors
4.2. Development of AGL-Edited Chinese Cabbage Lines and Selection Using PCR Analysis
4.3. Bolting Time Record and Stem Length Measurement of Selected AGL-Edited Lines
4.4. Identifying the Number of T-DNA Copies and Insertion Site
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xie, F.; Zha, J.; Tang, H.; Xu, Y.; Liu, X.; Wan, Z. Combining ability and heterosis analysis for mineral elements by using cytoplasmic male-sterile systems in non-heading Chinese cabbage (Brassica rapa). Crop Pasture Sci. 2018, 69, 296–302. [Google Scholar] [CrossRef]
- Kim, C.G. The Impact of Climate Change on the Agricultural Sector: Implications of the Agro-Industry For low Carbon, Green Growth Strategy and Roadmap for the East Asian Region; Korea Rural Economic Institute: Naju, Republic of Korea, 2008. [Google Scholar]
- Wang, Y.; Chen, L.; Su, W.; Hao, Y.; Liu, H.; Sun, G.; Chen, R.; Song, S. Effect of Nitrate Concentration on the Growth, Bolting and Related Gene Expression in Flowering Chinese Cabbage. Agron. J. 2021, 11, 936. [Google Scholar] [CrossRef]
- Wei, X.; Rahim, M.A.; Zhao, Y.; Yang, S.; Wang, Z.; Su, H.; Li, L.; Niu, L.; Harun-Ur-Rashid, M.; Yuan, Y.; et al. Comparative transcriptome analysis of early-and late-bolting traits in Chinese cabbage (Brassica rapa). Front. Genet. 2021, 12, 590830. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Huang, X.; Huang, X.; Su, W.; Hao, Y.; Liu, H.; Chen, R.; Song, S. BcSOC1 promotes bolting and stem elongation in flowering Chinese cabbage. Int. J. Mol. Sci. 2022, 23, 3459. [Google Scholar] [CrossRef]
- Liu, C.; Xi, W.; Shen, L.; Tan, C.; Yu, H. Regulation of floral patterning by flowering time genes. Dev. Cell. 2009, 16, 711–722. [Google Scholar] [CrossRef] [Green Version]
- Weigel, D.; Meyerowitz, E.M. Activation of floral homeotic genes in Arabidopsis. Science 1993, 261, 1723–1726. [Google Scholar] [CrossRef]
- Schönrock, N.; Bouveret, R.; Leroy, O.; Borghi, L.; Köhler, C.; Gruissem, W.; Hennig, L. Polycomb-group proteins repressthe floral activator AGL19 in the FLC-independent vernalization pathway. Genes Dev. 2006, 20, 1667–1678. [Google Scholar] [CrossRef] [Green Version]
- Michaels, S.D.; Ditta, G.; Gustafson-Brown, C.; Pelaz, S.; Yanofsky, M.; Amasino, R.M. AGL24 acts as a promoter of flowering in Arabidopsis and is positively regulated by vernalization. Plant J. 2003, 33, 867–874. [Google Scholar] [CrossRef]
- Jiang, W.; Zhou, W.; Li, Z.; Yan, K.; Wang, Y.; Wang, Z.; Song, M.; Tang, Q. Interactions of flowering promoting factor AGL19 with integrator factors AGL24 and SOC1 in Brassica oleracea var. italica. Acta Hortic. Sin. 2017, 44, 1905–1913. [Google Scholar]
- Hong, J.K.; Suh, E.J.; Park, S.R.; Park, J.; Lee, Y.H. Multiplex CRISPR/Cas9 Mutagenesis of BrVRN1 Delays Flowering Time in Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Agriculture 2021, 11, 1286. [Google Scholar] [CrossRef]
- Jeong, S.Y.; Ahn, H.M.; Ryu, J.H.; Oh, Y.B.; Sivanandhan, G.; Won, K.H.; Park, Y.D.; Kim, J.S.; Kim, H.R.; Lim, Y.P.; et al. Generation of early-flowering Chinese cabbage (Brassica rapa spp. pekinensis) through CRISPR/Cas9-mediated genome editing. Plant. Biotechnol. Rep. 2019, 13, 491–499. [Google Scholar] [CrossRef]
- Faisal, M.; Abdel-Salam, E.M.; Alatar, A.A. Artificial microRNA-based RNA interference and specific gene silencing for developing insect resistance in Solanum lycopersicum. Agron. J. 2021, 11, 136. [Google Scholar] [CrossRef]
- Shin, Y.H.; Lee, S.H.; Park, Y.D. Development of mite (Tetranychus urticae)-resistant transgenic Chinese cabbage using plant-mediated RNA interference. Hortic. Environ. Biotechnol. 2020, 61, 305–315. [Google Scholar] [CrossRef]
- Meng, X.; Wang, N.; He, H.; Tan, Q.; Wen, B.; Zhang, R.; Fu, X.; Xiao, W.; Chen, X.; Li, D.; et al. Prunus persica transcription factor PpNAC56 enhances heat resistance in transgenic tomatoes. Plant Physiol. Biochem. 2022, 182, 194–201. [Google Scholar] [CrossRef]
- Hu, L.; Lu, H.; Liu, Q.; Chen, X.; Jiang, X. Overexpression of mtl D gene in transgenic Populus tomentosa improves salt tolerance through accumulation of mannitol. Tree Physiol. 2005, 25, 1273–1281. [Google Scholar] [CrossRef] [Green Version]
- Alquézar, B.; Bennici, S.; Carmona, L.; Gentile, A.; Peña, L. Generation of Transfer-DNA-Free Base-Edited Citrus Plants. Front. Plant Sci. 2022, 13, 835282. [Google Scholar] [CrossRef]
- Zhang, A.; Liu, Y.; Wang, F.; Li, T.; Chen, Z.; Kong, D.; Bi, J.; Zhang, F.; Luo, X.; Wang, J.; et al. Enhanced rice salinity tolerance via CRISPR/Cas9-targeted mutagenesis of the OsRR22 gene. Mol. Breed. 2019, 39, 47. [Google Scholar] [CrossRef] [Green Version]
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A single transcript CRISPR/Cas9 mediated mutagenesis of CaERF28 confers anthracnose resistance in chilli pepper (Capsicum annuum L.). Planta 2021, 254, 5. [Google Scholar] [CrossRef]
- Marraffini, L.A.; Sontheimer, E.J. CRISPR interference: RNA-directed adaptive immunity in bacteria and archaea. Nat. Rev. Genet. 2010, 11, 181–190. [Google Scholar] [CrossRef]
- Jinek, M.; Chylinski, K.; Fonfara, I.; Hauer, M.; Doudna, J.A.; Charpentier, E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity. Science 2012, 337, 816–821. [Google Scholar] [CrossRef]
- Anzalone, A.V.; Koblan, L.W.; Liu, D.R. Genome editing with CRISPR–Cas nucleases, base editors, transposases and prime editors. Nat. Biotechnol. 2020, 38, 824–844. [Google Scholar] [CrossRef] [PubMed]
- Yeh, C.D.; Richardson, C.D.; Corn, J.E. Advances in genome editing through control of DNA repair pathways. Nat. Cell Biol. 2019, 21, 1468–1478. [Google Scholar] [CrossRef] [PubMed]
- Lieber, M.R. The mechanism of double-strand DNA break repair by the nonhomologous DNA end-joining pathway. Annu. Rev. Biochem. 2010, 79, 181–211. [Google Scholar] [CrossRef] [Green Version]
- Hentze, M.W.; Kulozik, A.E. A perfect message: RNA surveillance and nonsense-mediated decay. Cell 1999, 96, 307–310. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Xu, Y.; Tan, E.L.; Kumar, P.P. AGAMOUS-LIKE 24, a dosage-dependent mediator of the flowering signals. Proc. Natl. Acad. Sci. USA 2002, 99, 16336–16341. [Google Scholar] [CrossRef] [Green Version]
- Han, P.; García-Ponce, B.; Fonseca-Salazar, G.; Alvarez-Buylla, E.R.; Yu, H. AGAMOUS-LIKE 17, a novel flowering promoter, acts in a FT-independent photoperiod pathway. Plant J. 2008, 55, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Mandel, M.A.; Gustafson-Brown, C.; Savidge, B.; Yanofsky, M.F. Molecular characterization of the Arabidopsis floral homeotic gene APETALA1. Nature 1992, 360, 273–277. [Google Scholar] [CrossRef]
- Wagner, D.; Sablowski, R.W.; Meyerowitz, E.M. Transcriptional activation of APETALA1 by LEAFY. Science 1999, 285, 582–584. [Google Scholar] [CrossRef]
- Ng, M.; Yanofsky, M.F. Function and evolution of the plant MADS-box gene family. Nature reviews. Genetics 2001, 2, 186–195. [Google Scholar] [CrossRef]
- Parcy, F.; Nilsson, O.; Busch, M.A.; Lee, I.; Weigel, D. A genetic framework for floral patterning. Nature 1998, 395, 561–566. [Google Scholar] [CrossRef]
- Lenhard, M.; Bohnert, A.; Jürgens, G.; Laux, T. Termination of stem cell maintenance in Arabidopsis floral meristems by interactions between WUSCHEL and AGAMOUS. Cell 2001, 105, 805–814. [Google Scholar] [CrossRef] [PubMed]
- Lohmann, J.U.; Hong, R.L.; Hobe, M.; Busch, M.A.; Parcy, F.; Simon, R.; Weigel, D. A molecular link between stem cell regulation and floral patterning in Arabidopsis. Cell 2001, 105, 793–803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, H.F.; Huang, C.H.; Chou, L.T.; Yang, C.H. Ectopic expression of an orchid (Oncidium Gower Ramsey) AGL6-like gene promotes flowering by activating flowering time genes in Arabidopsis thaliana. Plant Cell Physiol. 2003, 44, 783–794. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Li, W.; Dong, X.; Guo, W.; Shu, H. Ectopic expression of a hyacinth AGL6 homolog caused earlier flowering and homeotic conversion in Arabidopsis. Sci. China Ser. C Life Sci. 2007, 50, 676–689. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Liang, W.; Jia, R.; Yin, C.; Zong, J.; Kong, H.; Zhang, D. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res. 2010, 20, 299–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, S.K.; Lee, J.S.; Ahn, J.H. Overexpression of AGAMOUS-LIKE 28 (AGL28) promotes flowering by upregulating expression of floral promoters within the autonomous pathway. Biochem. Biophys. Res. Commun. 2006, 348, 929–936. [Google Scholar] [CrossRef]
- Kim, D.H. Current understanding of flowering pathways in plants: Focusing on the vernalization pathway in Arabidopsis and several vegetable crop plants. Hortic. Environ. Biotechnol. 2020, 61, 209–227. [Google Scholar] [CrossRef]
- Searle, I.; He, Y.; Turck, F.; Vincent, C.; Fornara, F.; Krober, S.; Amasino, R.A.; Coupland, G. The transcription factor FLC confers a flowering response to vernalization by repressing meristem competence and systemic signaling in Arabidopsis. Genes Dev. 2006, 20, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Michaels, S.D.; Amasino, R.M. Flowering Locus C encodes a novel MADS domain protein that acts as a repressor of flowering. Plant Cell. 1999, 11, 949–956. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wu, J.J.; Tang, T.; Liu, K.D.; Dai, C. CRISPR/Cas9-mediated genome editing efficiently creates specific mutations at multiple loci using one sgRNA in Brassica napus. Sci. Rep. 2017, 7, 7489. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Wang, Y.; Li, T.; Qiu, H.; Xia, Z.; Dong, Y. Target-specific mutations efficiency at multiple loci of CRISPR/Cas9 system using one sgRNA in soybean. Transgenic Res. 2021, 30, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Shin, N.R.; Shin, Y.H.; Kim, H.S.; Park, Y.D. Function Analysis of the PR55/B Gene Related to Self-Incompatibility in Chinese Cabbage Using CRISPR/Cas9. Int. J. Mol. Sci. 2022, 23, 5062. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Luo, D.; Usman, B.; Nawaz, G.; Zhao, N.; Liu, F.; Li, R. Development of high yielding glutinous cytoplasmic male sterile rice (Oryza sativa L.) lines through CRISPR/Cas9 based mutagenesis of Wx and TGW6 and proteomic analysis of anther. Agronomy 2018, 8, 290. [Google Scholar] [CrossRef] [Green Version]
- Usman, B.; Nawaz, G.; Zhao, N.; Liu, Y.; Li, R. Generation of high yielding and fragrant rice (Oryza sativa L.) lines by CRISPR/Cas9 targeted mutagenesis of three homoeologs of cytochrome P450 gene family and OsBADH2 and transcriptome and proteome profiling of revealed changes triggered by mutations. Plants 2020, 9, 788. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, R.; Gao, J.; Gu, T.; Song, G.; Li, W.; Li, D.; Li, Y.; Li, G. Highly efficient and heritable targeted mutagenesis in wheat via the Agrobacterium tumefaciens-mediated CRISPR/Cas9 system. Int. J. Mol. Sci. 2019, 20, 4257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.F.; Li, H.; Qin, R.Y.; Li, J.; Qiu, C.H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Generation of inheritable and “transgene clean” targeted genome-modified rice in later generations using the CRISPR/Cas9 system. Sci. Rep. 2015, 5, 11491. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.P.; Xing, H.L.; Dong, L.; Zhang, H.Y.; Han, C.Y.; Wang, X.C.; Chen, Q.J. Egg cell-specific promoter-controlled CRISPR/Cas9 efficiently generates homozygous mutants for multiple target genes in Arabidopsis in a single generation. Genome Biol. 2015, 16, 905. [Google Scholar] [CrossRef]
- Jyothishwaran, G.; Kotresha, D.; Selvaraj, T.; Srideshikan, S.M.; Rajvanshi, P.K.; Jayabaskaran, C. A modified freeze–thaw method for efficient transformation of Agrobacterium tumefaciens. Curr. Sci. 2007, 93, 770–772. [Google Scholar]
- Lee, G.H.; Yu, J.G.; Park, Y.D. Development of an effective PCR technique for analyzing T-DNA integration sites in Brassica species and its application. Hortic. Sci. 2015, 33, 242–250. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| E0 Gene-Edited Line | E1 Gene-Edited Line | Chr. z | Target Gene | Mutation Type y |
|---|---|---|---|---|
| A1-2 | A1-2-5 | A03 | CT001_A03121400 | – |
| A08 | CT001_A08282630 | – | ||
| A1-9 | A1-9-2 | A03 | CT001_A03121400 | 1 bp In (T) |
| A08 | CT001_A08282630 | – | ||
| A2-1 | A2-1-2 | A03 | CT001_A03122450 | 60 bp Del |
| A01 | CT001_A01013460 | 1 bp Del (T) | ||
| A2-11 | A2-11-1 | A03 | CT001_A03122450 | 1 bp In (A) |
| A01 | CT001_A01013460 | 1 bp In (C) | ||
| A2-11-6 | A03 | CT001_A03122450 | 1 bp In (A) | |
| A01 | CT001_A01013460 | 1 bp Tv, 7 bp Del | ||
| A2-16 | A2-16-4 | A03 | CT001_A03122450 | 1 bp In (C) |
| A01 | CT001_A01013460 | 6 bp Del | ||
| A2-16-12 | A03 | CT001_A03122450 | 1 bp In (C) | |
| A01 | CT001_A01013460 | 6 bp Del | ||
| A2-22 | A2-22-5 | A03 | CT001_A03122450 | – |
| A01 | CT001_A01013460 | 120 bp Del |
| Reaction | Number of Cycles | Thermal Settings |
|---|---|---|
| Primary (AD-LSP1/RSP1) z | 1 | 95 °C 2 min |
| 5 | 94 °C 30 s, 72 °C 4 min | |
| 2 | 94 °C 30 s, 25 °C ramping to 72 °C in 4 min | |
| 15 | 94 °C 30 s, 72 °C 4 min 94 °C 20 s, 72 °C 4 min 94 °C 30 s, 44 °C 1 min, 72 °C 2 min 30 s | |
| 1 | 72 °C 5 min, 16 °C hold | |
| Secondary (AD-LSP2/RSP2) | 1 | 95 °C 2 min |
| 5 | 94 °C 20 s, 72 °C 4 min | |
| 15 | 94 °C 30 s, 72 °C 4 min 94 °C 20 s, 72 °C 4 min 94 °C 30 s, 44 °C 1 min, 72 °C 2 min 30 s | |
| 5 | 94 °C 20 s, 44 °C 1 min, 72 °C 2 min 30 s | |
| 1 | 72 °C 5 min, 16 °C hold |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shin, Y.-H.; Lee, H.-M.; Park, Y.-D. CRISPR/Cas9-Mediated Editing of AGAMOUS-like Genes Results in a Late-Bolting Phenotype in Chinese Cabbage (Brassica rapa ssp. pekinensis). Int. J. Mol. Sci. 2022, 23, 15009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315009
Shin Y-H, Lee H-M, Park Y-D. CRISPR/Cas9-Mediated Editing of AGAMOUS-like Genes Results in a Late-Bolting Phenotype in Chinese Cabbage (Brassica rapa ssp. pekinensis). International Journal of Molecular Sciences. 2022; 23(23):15009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315009
Chicago/Turabian StyleShin, Yun-Hee, Hyun-Min Lee, and Young-Doo Park. 2022. "CRISPR/Cas9-Mediated Editing of AGAMOUS-like Genes Results in a Late-Bolting Phenotype in Chinese Cabbage (Brassica rapa ssp. pekinensis)" International Journal of Molecular Sciences 23, no. 23: 15009. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315009