
Transcriptome and Metabolite Profiling of Tomato SGR-Knockout Null Lines Using the CRISPR/Cas9 System

 , , and
, , and
Abstract
:1. Introduction
2. Results
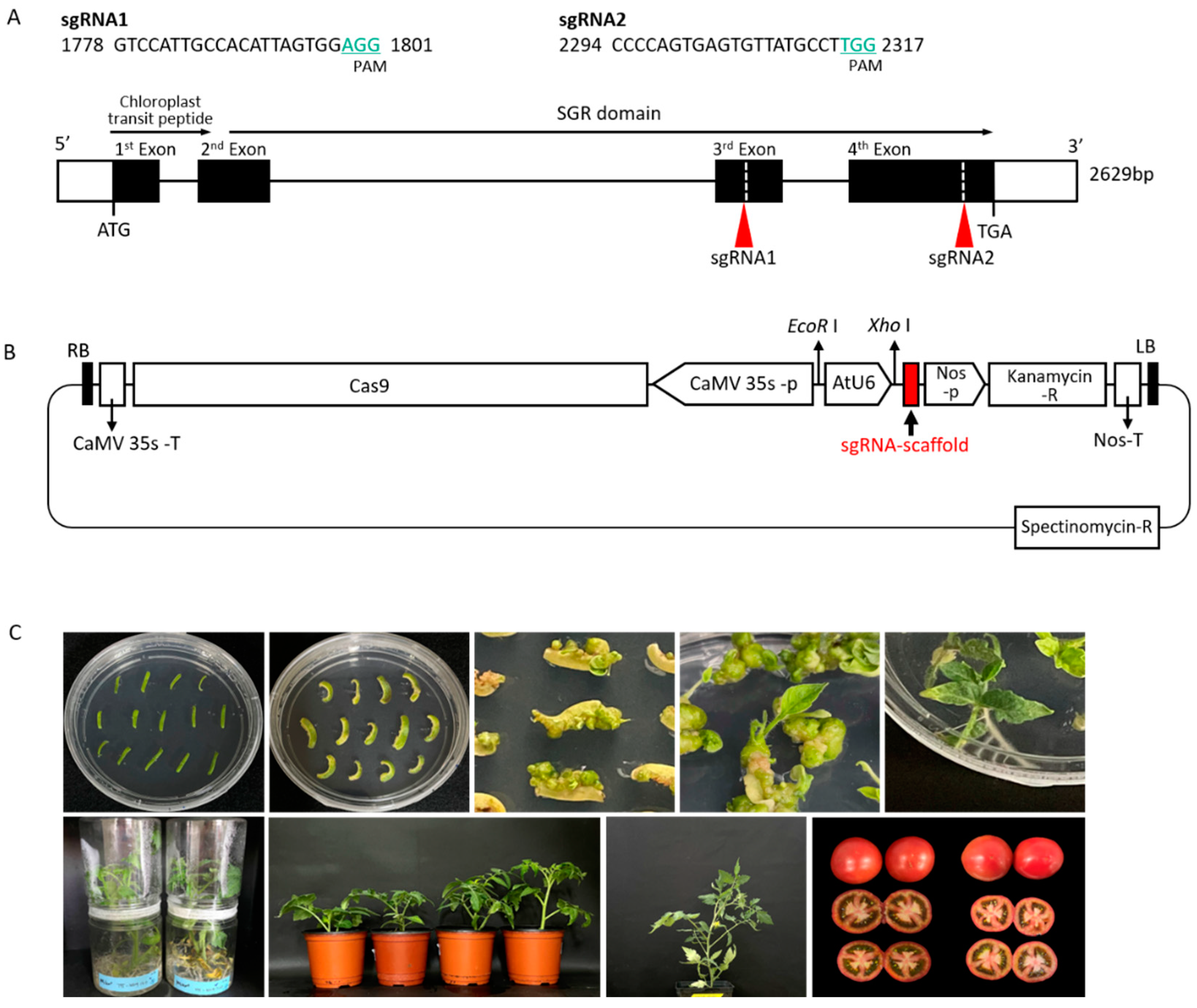
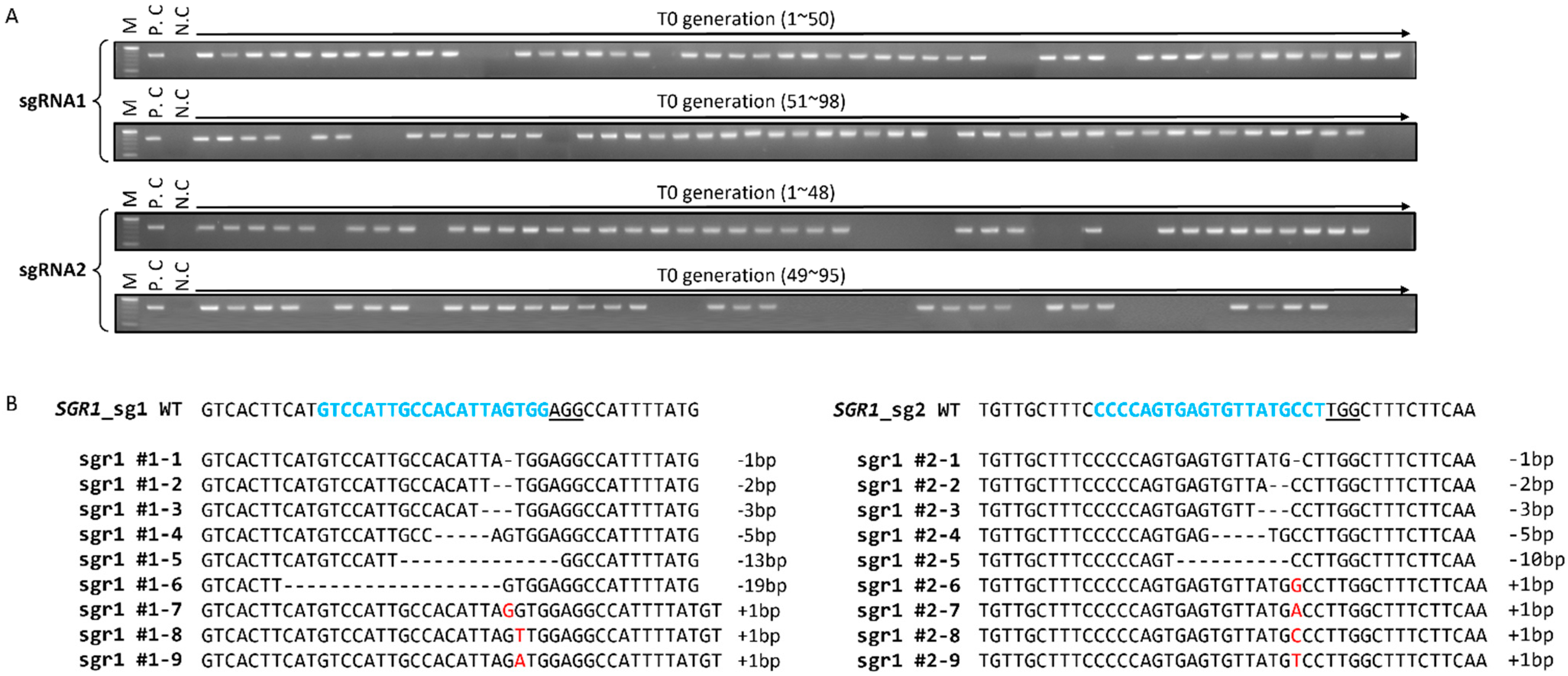
2.1. CRISPR/Cas9-Targeted Mutagenesis of SlSGR1
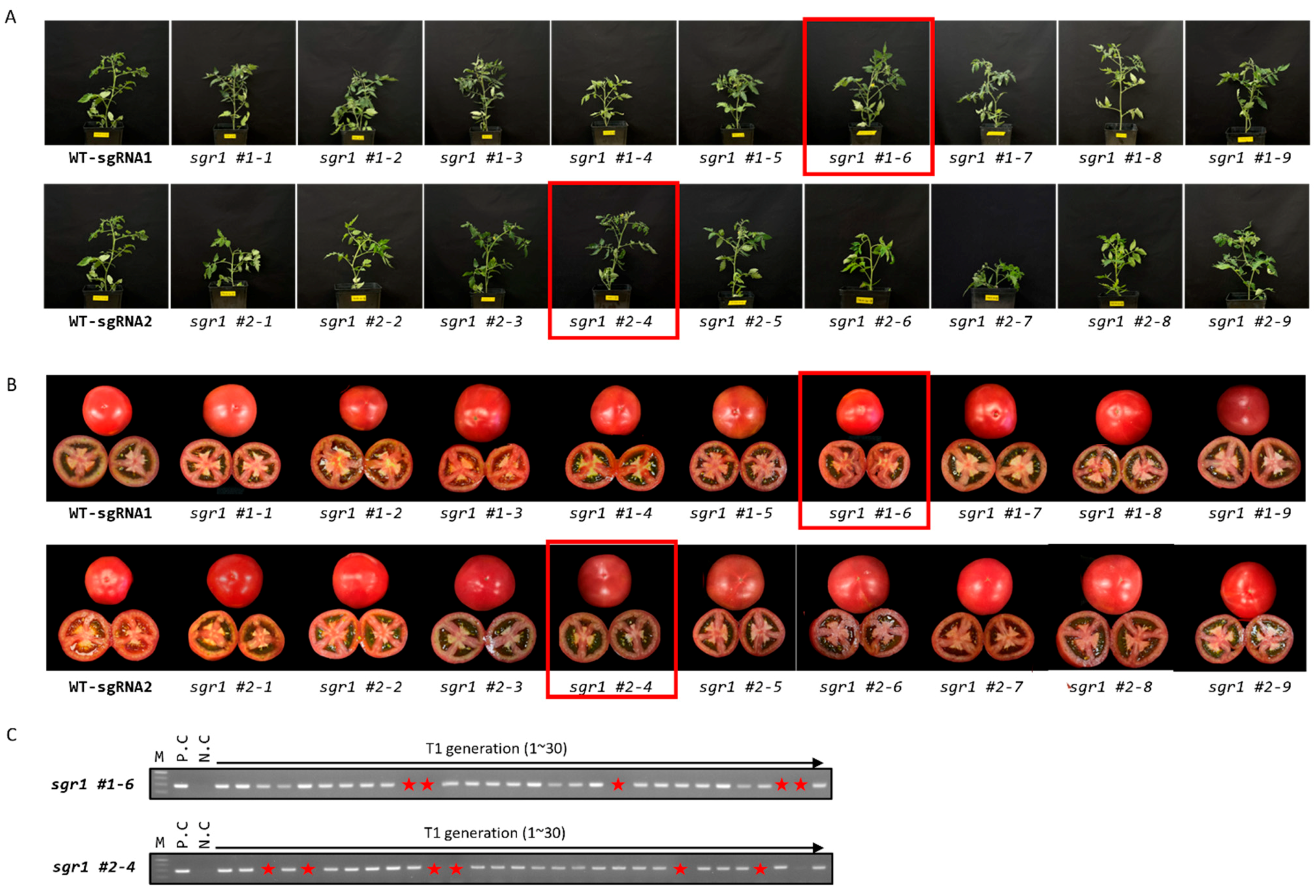
2.2. Selection and Characterization of sgr1 Null Lines
2.3. Carotenoid Profiles in the Leaves and Fruits of slsgr1 Mutant Lines
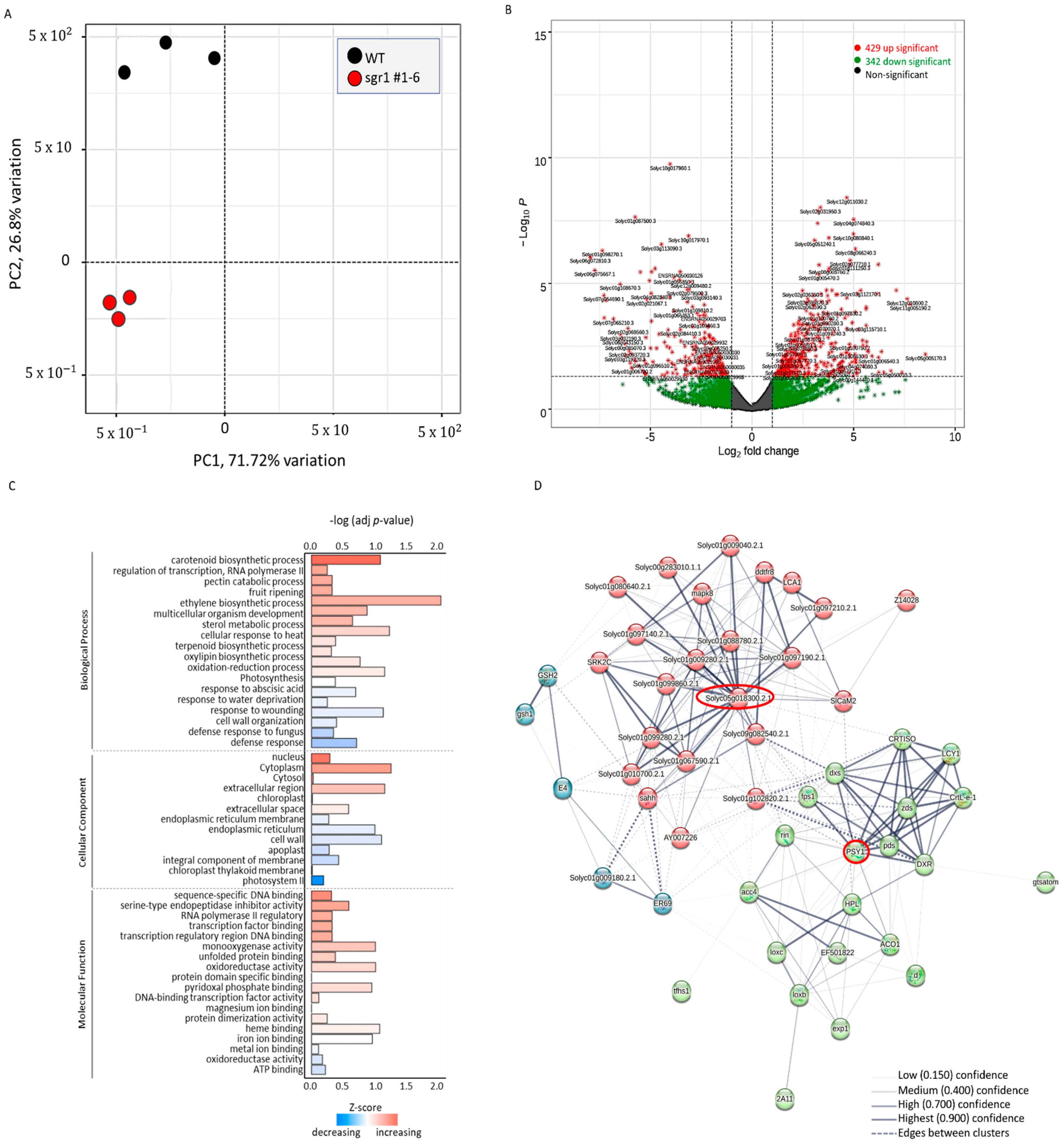
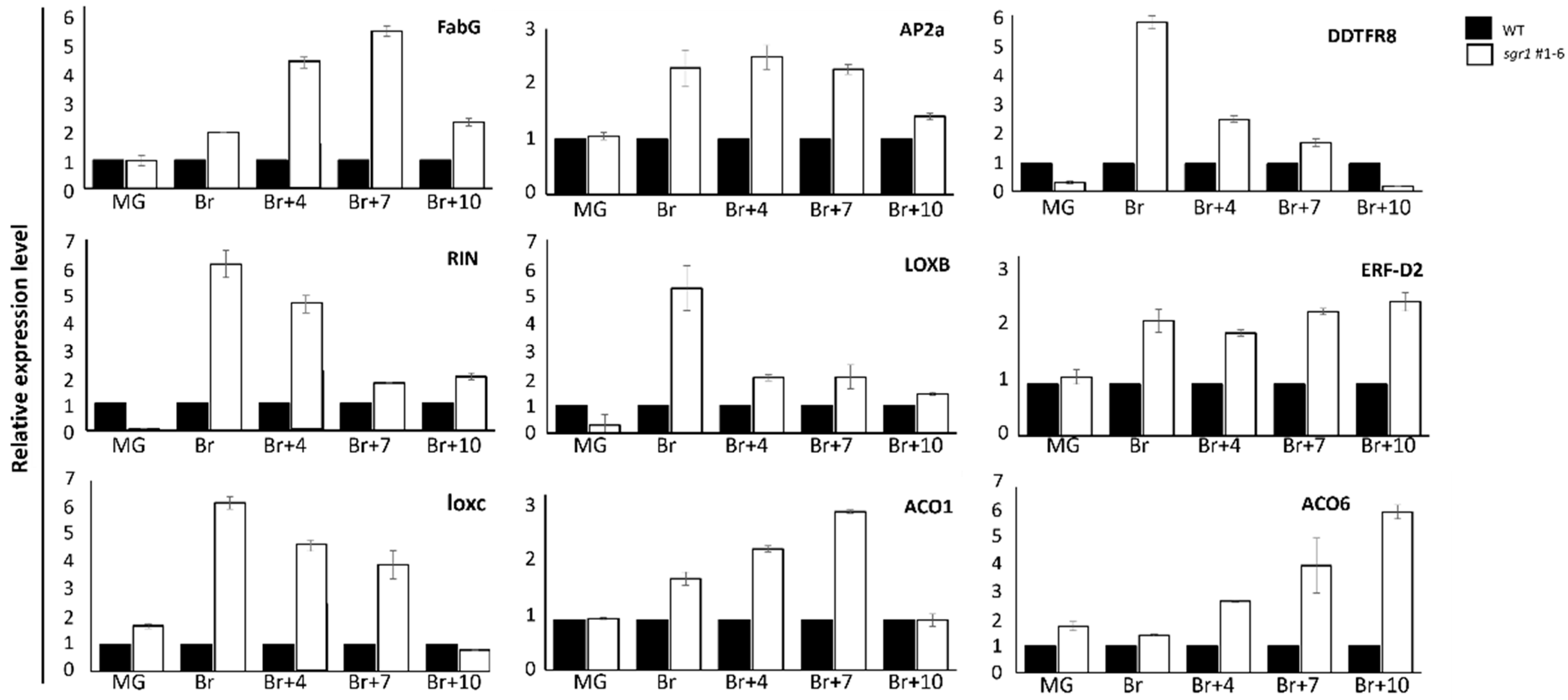
2.4. Transcriptome Analysis of sgr#1−6 Mutant and WT Plants
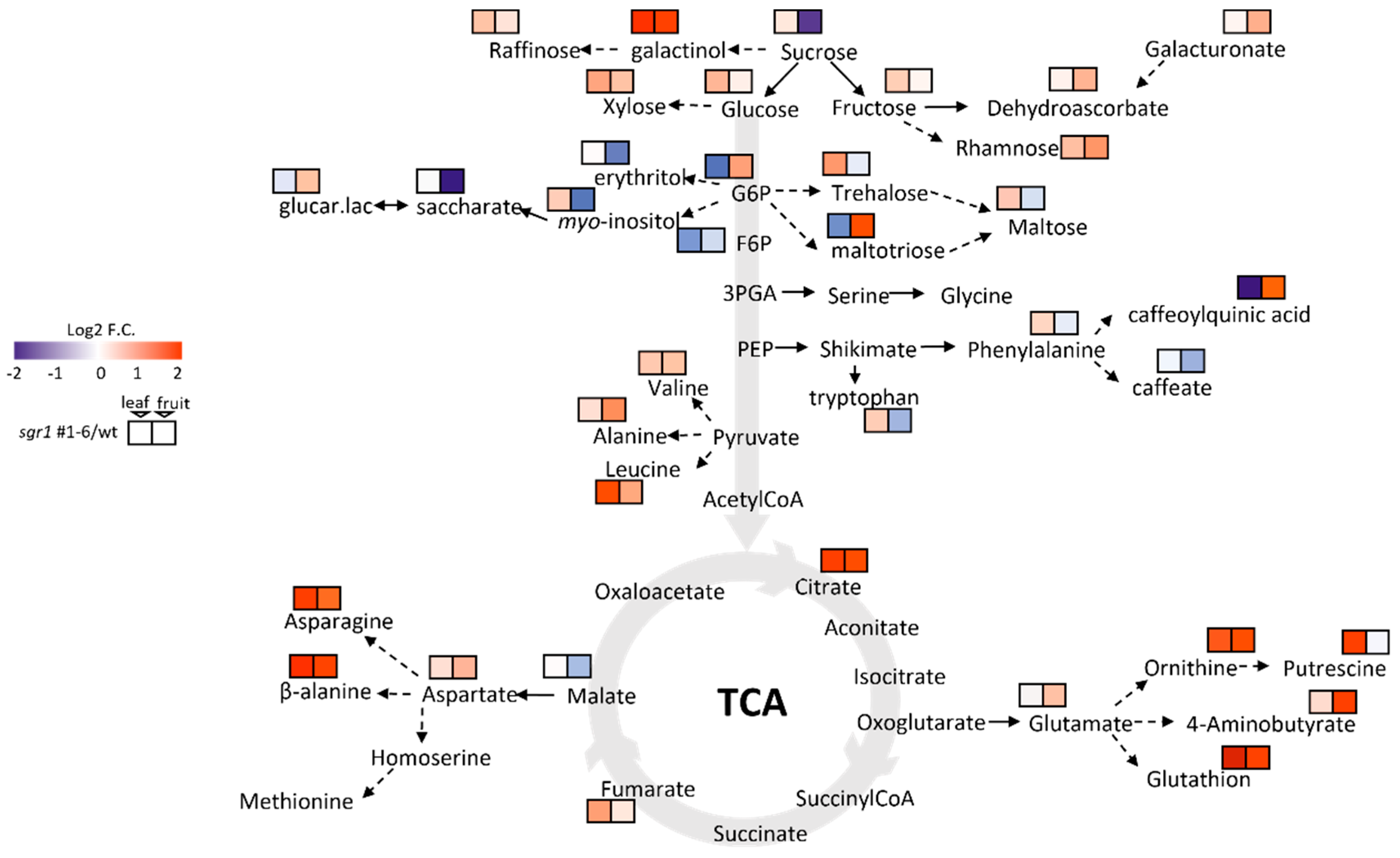
2.5. Metabolite Profiling of the sgr1 #1−6 Null Lines
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Conditions
4.2. Plasmid Construction and Genetic Transformation of Tomato
4.3. Mutation Analysis of Transgenic Lines
4.4. Determination of Chlorophyll and Carotenoid Levels
4.5. RNA Extraction, Library Preparation and RNA-Seq
4.6. qRT-PCR Analysis
4.7. Resequencing Analysis
4.8. Metabolite Profile Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; Xu, J. Meta-analysis of the association between dietary lycopene intake and ovarian cancer risk in postmenopausal women. Sci. Rep. 2014, 4, 5674. [Google Scholar] [CrossRef] [Green Version]
- Pouchieu, C.; Galan, P.; Ducros, V.; Latino-Martel, P.; Hercberg, S.; Touvier, M. Plasma carotenoids and retinol and overall and breast cancer risk: A nested case-control study. Nutr. Cancer 2014, 66, 980–988. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Lee, A.H.; Su, D.; Binns, C.W. Fruit and vegetable consumption associated with reduced risk of epithelial ovarian cancer in southern Chinese women. Gynecol. Oncol. 2014, 132, 241–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chappell, J. The biochemistry and molecular biology of isoprenoid metabolism. Plant Physiol. 1995, 107, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Giuliano, G.; Bartley, G.E.; Scolnik, P.A. Regulation of carotenoid biosynthesis during tomato development. Plant Cell 1993, 5, 379–387. [Google Scholar]
- Fraser, P.D.; Truesdale, M.R.; Bird, C.R.; Schuch, W.; Bramley, P.M. Carotenoid biosynthesis during tomato fruit development (evidence for tissue-specific gene expression). Plant Physiol. 1994, 105, 405–413. [Google Scholar] [CrossRef] [Green Version]
- Fraser, P.D.; Kiano, J.W.; Truesdale, M.R.; Schuch, W.; Bramley, P.M. Phytoene synthase-2 enzyme activity in tomato does not contribute to carotenoid synthesis in ripening fruit. Plant Mol. Biol. 1999, 40, 687–698. [Google Scholar] [CrossRef]
- Rosati, C.; Aquilani, R.; Dharmapuri, S.; Pallara, P.; Marusic, C.; Tavazza, R.; Bouvier, F.; Camara, B.; Giuliano, G. Metabolic engineering of beta-carotene and lycopene content in tomato fruit. Plant J. 2000, 24, 413–420. [Google Scholar] [CrossRef]
- Galpaz, N.; Wang, Q.; Menda, N.; Zamir, D.; Hirschberg, J. Abscisic acid deficiency in the tomato mutant high-pigment 3 leading to increased plastid number and higher fruit lycopene content. Plant J. 2008, 53, 717–730. [Google Scholar] [CrossRef]
- Chen, W.; He, S.; Liu, D.; Patil, G.B.; Zhai, H.; Wang, F.; Stephenson, T.J.; Wang, Y.; Wang, B.; Valliyodan, B.; et al. A sweet potato geranylgeranyl pyrophosphate synthase gene, IbGGPS, increases carotenoid content and enhances osmotic stress tolerance in Arabidopsis thaliana. PLoS ONE 2015, 10, e0137623. [Google Scholar]
- Ronen, G.; Carmel-Goren, L.; Zamir, D.; Hirschberg, J. An alternative pathway to β-carotene formation in plant chromoplasts discovered by map-based cloning of Beta and old-gold color mutations in tomato. Proc. Natl. Acad. Sci. USA 2000, 97, 11102–11107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fraser, P.D.; Enfissi, E.M.; Halket, J.M.; Truesdale, M.R.; Yu, D.; Gerrish, C.; Bramley, P.M. Manipulation of phytoene levels in tomato fruit: Effects on isoprenoids, plastids, and intermediary metabolism. Plant Cell 2007, 19, 3194–3211. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Yuan, B.; Zhang, M.; Wang, L.; Cui, M.; Wang, Q.; Leng, P. Fruit-specific RNAi-mediated suppression of SlNCED1 increases both lycopene and β-carotene contents in tomato fruit. J. Exp. Bot. 2012, 63, 3097–3108. [Google Scholar] [CrossRef] [Green Version]
- Luo, Z.; Zhang, J.; Li, J.; Yang, C.; Wang, T.; Ouyang, B.; Li, H.; Giovannoni, J.; Ye, Z. A STAY-GREEN protein S l SGR 1 regulates lycopene and β-carotene accumulation by interacting directly with S l PSY 1 during ripening processes in tomato. New Phytol. 2013, 198, 442–452. [Google Scholar] [CrossRef] [PubMed]
- Ren, G.; An, K.; Liao, Y.; Zhou, X.; Cao, Y.; Zhao, H.; Ge, X.; Kuai, B. Identification of a novel chloroplast protein AtNYE1 regulating chlorophyll degradation during leaf senescence in Arabidopsis. Plant Physiol. 2007, 144, 1429–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Schelbert, S.; Park, S.Y.; Han, S.H.; Lee, B.D.; Andrès, C.B.; Kessler, F.; Hörtensteiner, S.; Paek, N.C. STAY-GREEN and chlorophyll catabolic enzymes interact at light-harvesting complex II for chlorophyll detoxification during leaf senescence in Arabidopsis. Plant Cell 2012, 24, 507–518. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Park, S.Y.; Kim, Y.S.; Wang, S.H.; Yoo, S.C.; Hörtensteiner, S.; Paek, N.C. Arabidopsis STAY-GREEN2 is a negative regulator of chlorophyll degradation during leaf senescence. Mol. Plant 2014, 7, 1288–1302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakuraba, Y.; Kim, D.; Kim, Y.S.; Hörtensteiner, S.; Paek, N.C. Arabidopsis STAYGREEN-LIKE (SGRL) promotes abiotic stress-induced leaf yellowing during vegetative growth. FEBS Lett. 2014, 588, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Shimoda, Y.; Matsuda, K.; Tanaka, A.; Ito, H. Mg-dechelation of chlorophyll a by Stay-Green activates chlorophyll b degradation through expressing Non-Yellow Coloring 1 in Arabidopsis thaliana. J. Plant Physiol. 2018, 222, 94–102. [Google Scholar] [CrossRef]
- Ono, K.; Kimura, M.; Matsuura, H.; Tanaka, A.; Ito, H. Jasmonate production through chlorophyll a degradation by Stay-Green in Arabidopsis thaliana. J. Plant Physiol. 2019, 238, 53–62. [Google Scholar] [CrossRef]
- Xie, Z.; Wu, S.; Chen, J.; Zhu, X.; Zhou, X.; Hörtensteiner, S.; Ren, G.; Kuai, B. The C-terminal cysteine-rich motif of NYE1/SGR1 is indispensable for its function in chlorophyll degradation in Arabidopsis. Plant Mol. Biol. 2019, 101, 257–268. [Google Scholar] [CrossRef] [PubMed]
- Cheung, A.Y.; Mcnellis, T.; Pieokos, B. Maintenance of chloroplast components during chromoplast differentiation in the tomato mutant green flesh. Plant Physiol. 1993, 101, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Akhtar, M.S.; Goldschmidt, E.E.; John, I.; Rodoni, S.; Matile, P.; Grierson, D. Altered patterns of senescence and ripening in gf, a stay-green mutant of tomato (Lycopersicon esculentum Mill.). J. Exp. Bot. 1999, 50, 1115–1122. [Google Scholar] [CrossRef]
- Barry, C.S.; McQuinn, R.P.; Chung, M.Y.; Besuden, A.; Giovannoni, J.J. Amino acid substitutions in homologs of the STAY-GREEN protein are responsible for the green-flesh and chlorophyll retainer mutations of tomato and pepper. Plant Physiol. 2008, 147, 179–187. [Google Scholar] [CrossRef] [Green Version]
- Hu, Z.-L.; Deng, L.; Yan, B.; Pan, Y.; Luo, M.; Chen, X.-Q.; Hu, T.-Z.; Chen, G.-P. Silencing of the LeSGR1 gene in tomato inhibits chlorophyll degradation and exhibits a stay-green phenotype. Biol. Plant 2011, 55, 27–34. [Google Scholar] [CrossRef]
- Park, S.Y.; Yu, J.W.; Park, J.S.; Li, J.; Yoo, S.C.; Lee, N.Y.; Lee, S.-K.; Jeong, S.-W.; Seo, H.S.; Koh, H.-J.; et al. The senescence-induced STAYGREEN protein regulates chlorophyll degradation. Plant Cell 2007, 19, 1649–1664. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Li, M.; Liang, N.; Yan, H.; Wei, Y.; Xu, X.; Liu, J.; Xu, Z.; Chen, F.; Wu, G. Molecular cloning and function analysis of the stay green gene in rice. Plant J. 2007, 52, 197–209. [Google Scholar] [CrossRef]
- Sato, Y.; Morita, R.; Katsuma, S.; Nishimura, M.; Tanaka, A.; Kusaba, M. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J. 2009, 57, 120–131. [Google Scholar] [CrossRef]
- Jiang, H.; Chen, Y.; Li, M.; Xu, X.; Wu, G. Overexpression of SGR results in oxidative stress and lesion-mimic cell death in rice seedlings. J. Integr. Plant Biol. 2011, 53, 375–387. [Google Scholar] [CrossRef]
- Rong, H.; Tang, Y.; Zhang, H.; Wu, P.; Chen, Y.; Li, M.; Wu, G.; Jiang, H. The Stay-Green Rice like (SGRL) gene regulates chlorophyll degradation in rice. J. Plant Physiol. 2013, 170, 1367–1373. [Google Scholar] [CrossRef]
- Borovsky, Y.; Paran, I. Chlorophyll breakdown during pepper fruit ripening in the chlorophyll retainer mutation is impaired at the homolog of the senescence-inducible stay-green gene. Appl. Genet. 2008, 117, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Han, L.; Pislariu, C.; Nakashima, J.; Fu, C.; Jiang, Q.; Quan, L.; Blancaflor, E.B.; Tang, Y.; Bouton, J.H.; et al. From model to crop: Functional analysis of a STAY-GREEN gene in the model legume Medicago truncatula and effective use of the gene for alfalfa improvement. Plant Physiol. 2011, 157, 1483–1496. [Google Scholar] [CrossRef]
- Shi, S.; Miao, H.; Du, X.; Gu, J.; Xiao, K. GmSGR1, a stay-green gene in soybean (Glycine max L.), plays an important role in regulating early leaf-yellowing phenotype and plant productivity under nitrogen deprivation. Acta Physiol. Plant 2016, 38, 97. [Google Scholar] [CrossRef]
- Hörtensteiner, S. Stay-green regulates chlorophyll and chlorophyll-binding protein degradation during senescence. Trends Plant Sci. 2009, 14, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Wei, W.; Fan, Z.; Zhao, X.; Zhang, Y.; Jing, Y.; Zhu, B.; Zhu, H.; Shan, W.; Chen, J.; et al. Re-evaluation of the nor mutation and the role of the NAC-NOR transcription factor in tomato fruit ripening. J. Exp. Bot. 2020, 71, 3560–3574. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Zeng, N.; Cheng, K.; Li, J.; Wang, K.; Zhang, C.; Zhu, H. Changes in fruit pigment accumulation, chloroplast development, and transcriptome analysis in the CRISPR/Cas9-mediated knockout of Stay-green 1 (slsgr1) mutant. Food Qual. Saf. 2022, 6, fyab029. [Google Scholar] [CrossRef]
- Ronen, G.; Cohen, M.; Zamir, D.; Hirschberg, J. Regulation of carotenoid biosynthesis during tomato fruit development: Expression of the gene for lycopene epsilon-cyclase is down-regulated during ripening and is elevated in the mutant Delta. Plant J. 1999, 17, 341–351. [Google Scholar] [CrossRef]
- Li, X.; Wang, Y.; Chen, S.; Tian, H.; Fu, D.; Zhu, B.; Luo, Y.; Zhu, H. Lycopene is enriched in tomato fruit by CRISPR/Cas9-mediated multiplex genome editing. Front. Plant. Sci. 2018, 9, 559. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.L.; Wang, L.; Tian, Y.; Jia, N.; Chen, S.; Shi, N.-B.; Huang, X.; Zhou, C.; Yu, Y.; et al. Regulation of ethylene-responsive SlWRKYs involved in color change during tomato fruit ripening. Sci. Rep. 2017, 7, 16674. [Google Scholar] [CrossRef] [Green Version]
- Bai, L.; Chen, Q.; Jiang, L.; Lin, Y.; Ye, Y.; Liu, P.; Wang, X.; Tang, H. Comparative transcriptome analysis uncovers the regulatory functions of long noncoding RNAs in fruit development and color changes of Fragaria pentaphylla. Hortic. Res. 2019, 6, 42. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.; Sandusky, P.; Bowlby, N.R.; Aebersold, R.; Green, B.R.; Vlahaskis, S.; Yocum, C.F.; Pichersky, E. Characterization of a spinach psbS cDNA encoding the 22 kDa protein of photosystem II. FEBS Lett. 1992, 314, 67–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pichersky, E.; Tanksley, S.D.; Piechulla, B.; Stayton, M.M.; Dunsmuir, P. Nucleotide sequence and chromosomal location of Cab-7, the tomato gene encoding the type II chlorophyll a/b-binding polypeptide of photosystem I. Plant Mol. Biol. 1988, 11, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Bang, I.H.; Kwon, O.K.; Hao, L.; Park, D.; Chung, M.J.; Oh, B.C.; Lee, S.; Bae, E.J.; Park, B.H. Deacetylation of XBP1s by sirtuin 6 confers resistance to ER stress-induced hepatic steatosis. Exp. Mol. Med. 2019, 51, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Farcuh, M.; Rivero, R.M.; Sadka, A.; Blumwald, E. Ethylene regulation of sugar metabolism in climacteric and non-climacteric plums. Postharvest Biol. Technol. 2018, 139, 20–30. [Google Scholar] [CrossRef]
- Lauxmann, M.A.; Borsani, J.; Osorio, S.; Lombardo, V.A.; Budde, C.O.; Bustamante, C.A.; Monti, L.L.; Andreo, C.S.; Fernie, A.R.; Drincovich, M.F.; et al. Deciphering the metabolic pathways influencing heat and cold responses during post-harvest physiology of peach fruit. Plant Cell Environ. 2014, 37, 601–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luengwilai, K.; Beckles, D.M.; Roessner, U.; Dias, D.A.; Lui, V.; Siriphanich, J. Identification of physiological changes and key metabolites coincident with postharvest internal browning of pineapple (Ananas comosus L.) fruit. Postharvest Biol. Technol. 2018, 137, 56–65. [Google Scholar] [CrossRef]
- Sato, S.; Tabata, S.; Hirakawa, H.; Asamizu, E.; Shirasawa, K.; Isobe, S.; Kaneko, T.; Nakamura, Y.; Shibata, D.; Aoki, K.; et al. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general-purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.A.; Jüppner, J.; Bajdzienko, K.; Giavalisco, P. Protocol: A fast, comprehensive and reproducible one-step extraction method for the rapid preparation of polar and semi-polar metabolites, lipids, proteins, starch and cell wall polymers from a single sample. Plant Methods 2016, 12, 45. [Google Scholar] [CrossRef] [Green Version]
- Luedemann, A.; Strassburg, K.; Erban, A.; Kopka, J. TagFinder for the quantitative analysis of gas chromatography-mass spectrometry (GC-MS)-based metabolite profiling experiments. Bioinformatics 2008, 24, 732–737. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Region | No. of Regenerated Plants | No. of Transgenic Plants | No. of Edited Plants | Genotype | |||
|---|---|---|---|---|---|---|---|
| Homoallelic | Heteroallelic | Biallelic | Multiallelic | ||||
| SlSGR1-sg 1 | 98 | 87 | 37 | 8 | 6 | 11 | 12 |
| SlSGR1-sg 2 | 95 | 69 | 31 | 4 | 7 | 9 | 11 |
| T1 Generation | Total Seeds | No. of Resistance | No. of Susceptible | X2-Test | p-Value |
|---|---|---|---|---|---|
| sgr1 #1−6 | 40 | 31 | 9 | 0.72 | 0.5 < p < 0.2 |
| sgr1 #2−4 | 39 | 30 | 9 | 0.78 | 0.5 < p < 0.2 |
| Lines | Lutein | β-Carotene | Lycopene | Others | Total |
|---|---|---|---|---|---|
| Mean ± SD * | Mean ± SD * | Mean ± SD * | Mean ± SD * | Mean ± SD * | |
| WT | 7.3 ± 1.9 | 18.1 ± 2.9 | 225.9 ± 25.6 | 142.5 ± 11.0 | 393.8 ±41.5 |
| sgr1 #1−6 | 4.3 ± 0.1 | 48.0 ± 6.6 | 999.5 ± 3.5 | 362.4 ± 15.3 | 1414.2 ± 25.5 |
| sgr1 #2−4 | 5.8 ± 0.1 | 30.8 ± 2.2 | 809.9 ± 8.8 | 317.4 ± 8.1 | 1263.9 ± 19.2 |
| Samples | Chlorophyll a | Chlorophyll b |
|---|---|---|
| Mean ± SD * | Mean ± SD * | |
| WT | 5670.1 ± 226.3 | 2302.4 ± 100.6 |
| sgr1 #1−6 | 7911.5 ± 113.2 | 3399.3 ± 63.1 |
| sgr1 #2−4 | 7491.1 ± 340.0 | 2934.9 ± 115.4 |
| Ontology * | GO Accession | GO Term | Gene Count | Gene IDs | Adjusted p-Value |
|---|---|---|---|---|---|
| BP | GO:0009835 | Fruit ripening | 15 | Solyc01g095080.3, Solyc02g091990.3, Solyc03g093610.1, Solyc03g118290.3, Solyc05g005560.3, Solyc05g050010.3, Solyc07g049530.3, Solyc07g049550.3, Solyc07g056570.1, Solyc07g064190.2, Solyc08g005610.3, Solyc09g089580.3, Solyc09g092480.2, Solyc10g080210.2, Solyc12g005940.2 | 1.10 × 10−7 |
| BP | GO:0016125 | Sterol metabolic process | 10 | Solyc01g109140.3, Solyc02g065750.2, Solyc02g070580.1, Solyc02g089160.3, Solyc04g078900.3, Solyc04g079730.1, Solyc07g049690.3, Solyc08g005610.3, Solyc10g007960.1, Solyc11g069800.1 | 6.70 × 10−5 |
| CC | GO:0005737 | Cytoplasm | 39 | Solyc01g099190.3, Solyc01g099760.3, Solyc01g099770.3, Solyc01g101170.3, Solyc01g101180.3, Solyc01g103390.3, Solyc01g111450.3, Solyc02g080790.3, Solyc03g111720.3, Solyc04g071260.3, Solyc04g082030.1, Solyc05g007940.3, Solyc05g023800.3, Solyc05g051750.3, Solyc06g005060.3, Solyc06g005260.3, Solyc06g009970.3, Solyc06g059885.1, Solyc06g073390.3, Solyc06g074350.3, Solyc06g076020.3, Solyc06g076570.2, Solyc06g083230.3, Solyc07g065840.2, Solyc08g014000.3, Solyc08g043170.3, Solyc08g080650.2, Solyc08g082820.3, Solyc09g007910.3, Solyc09g009390.3, Solyc09g092480.2, Solyc10g080500.2, Solyc10g085280.2, Solyc10g085870.1, Solyc10g086220.2, Solyc10g086410.3, Solyc12g006470.2, Solyc12g035890.2, Solyc12g098940.2 | 6.30 × 10−5 |
| BP | GO:0055114 | Oxidation–reduction process | 11 | Solyc01g109140.3, Solyc02g065750.2, Solyc02g070580.1, Solyc02g089160.3, Solyc04g078900.3, Solyc04g079730.1, Solyc07g049690.3, Solyc08g005610.3, Solyc10g007960.1, Solyc10g086500.1, Solyc11g069800.1 | 7.90 × 10−4 |
| CC | GO:0005576 | Extracellular region | 21 | Solyc01g008710.3, Solyc01g009590.3, Solyc01g067740.3, Solyc01g097240.3, Solyc02g079500.3, Solyc02g089350.3, Solyc02g093580.3, Solyc03g020060.3, Solyc03g123620.3, Solyc05g007940.3, Solyc05g007950.3, Solyc06g064520.3, Solyc06g068520.3, Solyc07g064190.2, Solyc08g080630.3, Solyc08g080650.2, Solyc09g084460.3, Solyc10g055810.2, Solyc11g011210.2, Solyc11g066130.1, Solyc11g066390.2 | 7.90 × 10−4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.Y.; Kim, J.H.; Jang, Y.H.; Yu, J.; Bae, S.; Kim, M.-S.; Cho, Y.-G.; Jung, Y.J.; Kang, K.K. Transcriptome and Metabolite Profiling of Tomato SGR-Knockout Null Lines Using the CRISPR/Cas9 System. Int. J. Mol. Sci. 2023, 24, 109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010109
Kim JY, Kim JH, Jang YH, Yu J, Bae S, Kim M-S, Cho Y-G, Jung YJ, Kang KK. Transcriptome and Metabolite Profiling of Tomato SGR-Knockout Null Lines Using the CRISPR/Cas9 System. International Journal of Molecular Sciences. 2023; 24(1):109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010109
Chicago/Turabian StyleKim, Jin Young, Jong Hee Kim, Young Hee Jang, Jihyeon Yu, Sangsu Bae, Me-Sun Kim, Yong-Gu Cho, Yu Jin Jung, and Kwon Kyoo Kang. 2023. "Transcriptome and Metabolite Profiling of Tomato SGR-Knockout Null Lines Using the CRISPR/Cas9 System" International Journal of Molecular Sciences 24, no. 1: 109. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms24010109