

Genomic Survey of Heat Shock Proteins in Liriodendron chinense Provides Insight into Evolution, Characterization, and Functional Diversities

Abstract
:1. Introduction
2. Results
2.1. Identification of HSP Gene Members in L. chinense
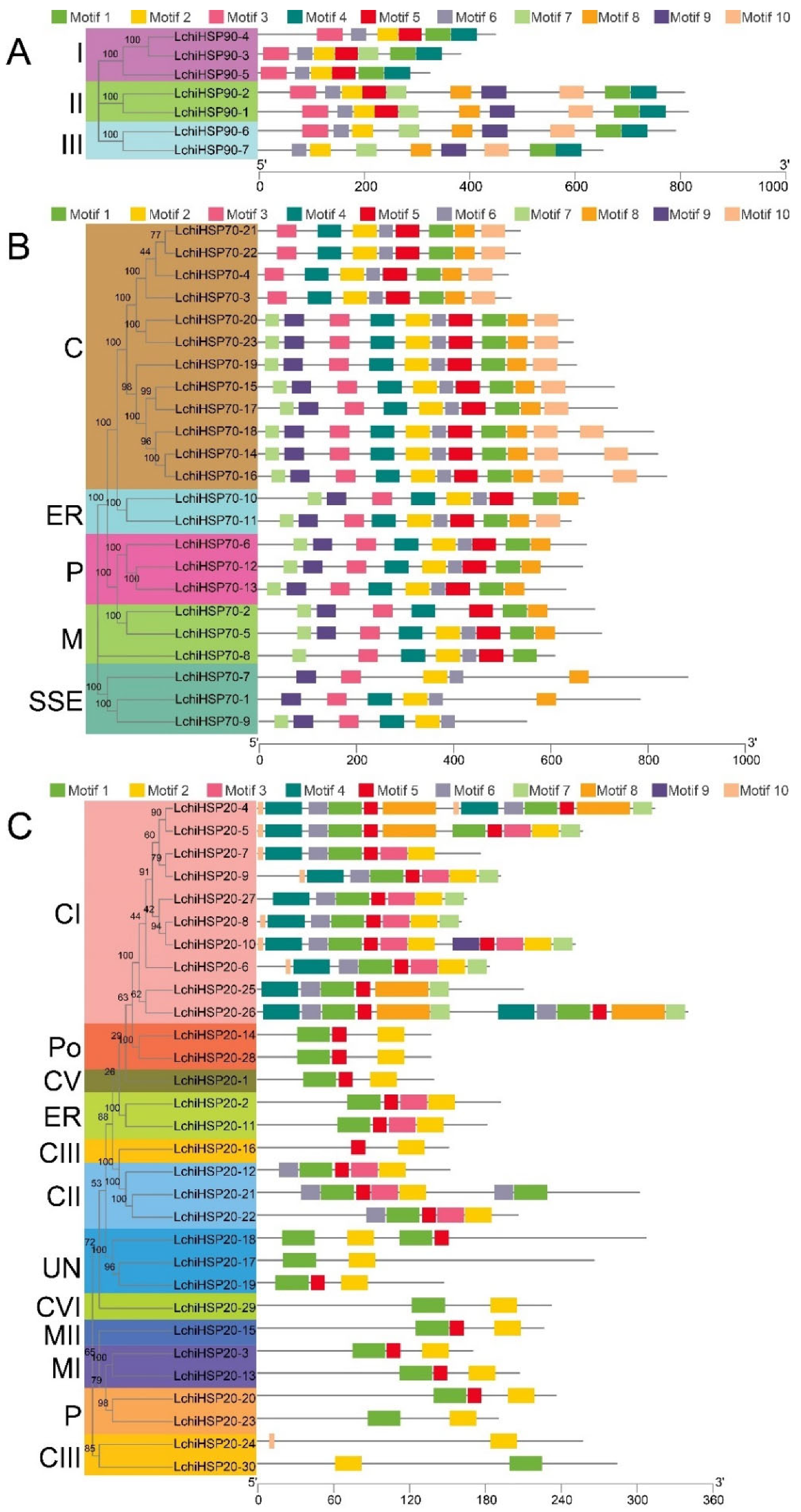
2.2. Phylogenetic Relationship of the LchiHSP Genes in L. chinense
2.3. Evolution of HSP Genes from Algae to Angiosperms
2.4. Gene Structure and Motif Analysis of the LchiHSP Genes
2.5. Gene Duplication and Collinearity Analysis of the LchiHSP Genes
2.6. Analysis of Cis-Element in LchiHSP Gene Promoters
2.7. Three-Dimensional Structures and Protein–Protein Network Analysis of LchiHSP Proteins
2.8. Expression Analysis of LchiHSPs under Heat and Cold Stress
3. Discussion
4. Materials and Methods
4.1. Identification and Characterization of HSP Genes in L. chinense
4.2. Systematic Evolution of HSP Gene Family
4.3. Gene Structure and Conserved Motif Analysis of LchiHSPs
4.4. Chromosome Distribution, Gene Duplication Events, and Collinearity Analysis
4.5. Cis-Regulatory Element Analysis of LchiHSP Gene Promoters
4.6. Three-Dimensional (3D) Protein Structures Prediction of LchiHSPs
4.7. Protein Interaction Network Analysis and Visualization
4.8. Expression Analysis of LchiHSP Genes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kidokoro, S.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant cold-stress responses. Trends Plant Sci. 2022, 27, 922–935. [Google Scholar] [CrossRef]
- Schulz, P.; Piepenburg, K.; Lintermann, R.; Herde, M.; Schottler, M.A.; Schmidt, L.K.; Ruf, S.; Kudla, J.; Romeis, T.; Bock, R. Improving plant drought tolerance and growth under water limitation through combinatorial engineering of signalling networks. Plant Biotechnol. J. 2021, 19, 74–86. [Google Scholar] [CrossRef]
- Shang, J.X.; Li, X.Y.; Li, C.L.; Zhao, L.Q. The role of nitric oxide in plant responses to salt stress. Int. J. Mol. Sci. 2022, 23, 6167. [Google Scholar] [CrossRef] [PubMed]
- Gill, R.A.; Ahmar, S.; Ali, B.; Saleem, M.H.; Khan, M.U.; Zhou, W.; Liu, S. The role of membrane transporters in plant growth and development, and abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 12792. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Vinocur, B.; Shoseyov, O.; Altman, A. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Sci. 2004, 9, 244–252. [Google Scholar] [CrossRef]
- Lindquist, S. The heat-shock response. Annu. Rev. Biochem. 1986, 55, 1151–1191. [Google Scholar] [CrossRef]
- Sun, X.; Huang, N.; Li, X.; Zhu, J.; Bian, X.; Li, H.; Wang, L.; Hu, Q.; Luo, H. A chloroplast heat shock protein modulates growth and abiotic stress response in creeping bentgrass. Plant Cell Environ. 2021, 44, 1769–1787. [Google Scholar] [CrossRef]
- Lindquist, S.; Craig, E.A. The heat-shock proteins. Annu. Rev. Genet. 1988, 22, 631–677. [Google Scholar] [CrossRef]
- Driedonks, N.; Xu, J.; Peters, J.L.; Park, S.; Rieu, I. Multi-level interactions between heat shock factors, heat shock proteins, and the redox system regulate acclimation to heat. Front. Plant Sci. 2015, 6, 999. [Google Scholar] [CrossRef]
- Wang, X.; Zheng, Y.; Chen, B.; Zhi, C.; Qiao, L.; Liu, C.; Pan, Y.; Cheng, Z. Genome-wide identification of small heat shock protein (HSP20) homologs in three cucurbit species and the expression profiles of CsHSP20s under several abiotic stresses. Int. J. Biol. Macromol. 2021, 190, 827–836. [Google Scholar] [CrossRef]
- Tabusam, J.; Shi, Q.; Feng, D.; Zulfiqar, S.; Shen, S.; Ma, W.; Zhao, J. HSP70 gene family in Brassica rapa: Genome-wide identification, characterization, and expression patterns in response to heat and cold stress. Cells 2022, 11, 2316. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Yang, Z.; Sui, M.; Cui, C.; Hu, Y.; Hou, X.; Xing, Q.; Huang, X.; Bao, Z. Identification and characterization of HSP90 gene family reveals involvement of HSP90, GRP94 and not TRAP1 in heat stress response in Chlamys farreri. Genes 2021, 12, 1592. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Pan, F.; Yang, C.; Jia, H.; Jiang, H.; He, F.; Li, N.; Lu, X.; Zhang, H. Genome-wide identification and expression analysis of HSP90 gene family in Nicotiana tabacum. BMC Genet. 2019, 20, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davoudi, M.; Chen, J.; Lou, Q. Genome-wide identification and expression analysis of heat shock protein 70 (HSP70) gene family in pumpkin (Cucurbita moschata) rootstock under drought stress suggested the potential role of these chaperones in stress tolerance. Int. J. Mol. Sci. 2022, 23, 1918. [Google Scholar] [CrossRef]
- Rehman, A.; Atif, R.M.; Qayyum, A.; Du, X.; Hinze, L.; Azhar, M.T. Genome-wide identification and characterization of HSP70 gene family in four species of cotton. Genomics 2020, 112, 4442–4453. [Google Scholar] [CrossRef]
- Waters, E.R.; Vierling, E. Plant small heat shock proteins—Evolutionary and functional diversity. New Phytol. 2020, 227, 24–37. [Google Scholar] [CrossRef] [Green Version]
- Gao, T.; Mo, Z.; Tang, L.; Yu, X.; Du, G.; Mao, Y. Heat shock protein 20 gene superfamilies in red algae: Evolutionary and functional diversities. Front. Plant Sci. 2022, 13, 817852. [Google Scholar] [CrossRef]
- Krishna, P.; Gloor, G. The Hsp90 family of proteins in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 238–246. [Google Scholar] [CrossRef]
- Jung, K.H.; Gho, H.J.; Nguyen, M.X.; Kim, S.R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, B.; Li, J.; Zhang, L.; Wang, Y.; Zheng, H.; Lu, M.; Chen, J. Hsf and Hsp gene families in Populus: Genome-wide identification, organization and correlated expression during development and in stress responses. BMC Genom. 2015, 16, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, L.; Hu, W.; Qian, Y.; Ren, Q.; Zhang, J. Genome-wide identification, classification and expression analysis of the Hsf and Hsp70 gene families in maize. Gene 2021, 770, 145348. [Google Scholar] [CrossRef]
- Sun, W.; Bernard, C.; van de Cotte, B.; Van Montagu, M.; Verbruggen, N. At-HSP17.6A, encoding a small heat-shock protein in Arabidopsis, can enhance osmotolerance upon overexpression. Plant J. 2001, 27, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.J.; Nou, I.S.; Kang, K.K. Overexpression of Oshsp16.9 gene encoding small heat shock protein enhances tolerance to abiotic stresses in rice. Plant Breed. Biotech. 2014, 2, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Liu, J.H.; Ma, X.; Zhai, Y.F.; Gong, Z.H.; Lu, M.H. Genome-wide analysis of the Hsp70 family genes in pepper (Capsicum annuum L.) and functional identification of CaHsp70-2 involvement in heat stress. Plant Sci. 2016, 252, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Yogendra, K.N.; Kumar, A.; Sarkar, K.; Li, Y.; Pushpa, D.; Mosa, K.A.; Duggavathi, R.; Kushalappa, A.C. Transcription factor StWRKY1 regulates phenylpropanoid metabolites conferring late blight resistance in potato. J. Exp. Bot. 2015, 66, 7377–7389. [Google Scholar] [CrossRef] [Green Version]
- Kallamadi, P.R.; Dandu, K.; Kirti, P.B.; Rao, C.M.; Thakur, S.S.; Mulpuri, S. An insight into powdery mildew-infected, susceptible, resistant, and immune sunflower genotypes. Proteomics 2018, 18, e1700418. [Google Scholar] [CrossRef]
- Farooq, M.A.; Zhang, X.; Zafar, M.M.; Ma, W.; Zhao, J. Roles of reactive oxygen species and mitochondria in seed germination. Front. Plant Sci. 2021, 12, 781734. [Google Scholar] [CrossRef]
- Ashraf, M.; Mao, Q.; Hong, J.; Shi, L.; Ran, X.; Liaquat, F.; Uzair, M.; Liang, W.; Fernie, A.R.; Shi, J. HSP70-16 and VDAC3 jointly inhibit seed germination under cold stress in Arabidopsis. Plant Cell Environ. 2021, 44, 3616–3627. [Google Scholar] [CrossRef]
- Samakovli, D.; Roka, L.; Dimopoulou, A.; Plitsi, P.K.; Zukauskait, A.; Georgopoulou, P.; Novak, O.; Milioni, D.; Hatzopoulos, P. HSP90 affects root growth in Arabidopsis by regulating the polar distribution of PIN1. New Phytol. 2021, 231, 1814–1831. [Google Scholar] [CrossRef]
- Cao, Y.; Feng, J.; Hwarari, D.; Ahmad, B.; Wu, H.; Chen, J.; Yang, L. Alterations in population distribution of Liriodendron chinense (Hemsl.) Sarg. and Liriodendron tulipifera Linn. caused by climate change. Forests 2022, 13, 488. [Google Scholar] [CrossRef]
- Tu, Z.; Shen, Y.; Wen, S.; Zong, Y.; Li, H. Alternative splicing enhances the transcriptome complexity of Liriodendron chinense. Front. Plant Sci. 2020, 11, 578100. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L.; Long, Y.; Si, W.; Cheng, B.; Jiang, H. A novel heat shock transcription factor (ZmHsf08) negatively regulates salt and drought stress responses in Maize. Int. J. Mol. Sci. 2021, 22, 11922. [Google Scholar] [CrossRef]
- Zhao, Y.; Du, H.; Wang, Y.; Wang, H.; Yang, S.; Li, C.; Chen, N.; Yang, H.; Zhang, Y.; Zhu, Y.; et al. The calcium-dependent protein kinase ZmCDPK7 functions in heat-stress tolerance in maize. J. Integr. Plant Biol. 2021, 63, 510–527. [Google Scholar] [CrossRef]
- Chen, J.; Gao, T.; Wan, S.; Zhang, Y.; Yang, J.; Yu, Y.; Wang, W. Genome-wide identification, classification and expression analysis of the HSP gene superfamily in tea plant (Camellia sinensis). Int. J. Mol. Sci. 2018, 19, 2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Hao, Z.; Guang, X.; Zhao, C.; Wang, P.; Xue, L.; Zhu, Q.; Yang, L.; Sheng, Y.; Zhou, Y.; et al. Liriodendron genome sheds light on angiosperm phylogeny and species-pair differentiation. Nat. Plants 2019, 5, 18–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaw, S.M.; Liu, Y.C.; Wu, Y.W.; Wang, H.Y.; Lin, C.I.; Wu, C.S.; Ke, H.M.; Chang, L.Y.; Hsu, C.Y.; Yang, H.T.; et al. Stout camphor tree genome fills gaps in understanding of flowering plant genome evolution. Nat. Plants 2019, 5, 63–73. [Google Scholar] [CrossRef] [Green Version]
- Yao, F.; Song, C.; Wang, H.; Song, S.; Jiao, J.; Wang, M.; Zheng, X.; Bai, T. Genome-wide characterization of the HSP20 gene family identifies potential members involved in temperature stress response in apple. Front Genet. 2020, 11, 609184. [Google Scholar] [CrossRef]
- Zhao, P.; Wang, D.; Wang, R.; Kong, N.; Zhang, C.; Yang, C.; Wu, W.; Ma, H.; Chen, Q. Genome-wide analysis of the potato Hsp20 gene family: Identification, genomic organization and expression profiles in response to heat stress. BMC Genom. 2018, 19, 61. [Google Scholar] [CrossRef]
- Wang, L.; Liu, F.; Ju, L.; Xue, B.; Wang, Y.; Wang, D.; Hou, D. Genome structures and evolution analysis of Hsp90 gene family in Brassica napus reveal the possible roles of members in response to salt stress and the infection of Sclerotinia sclerotiorum. Front. Plant Sci. 2022, 13, 854034. [Google Scholar] [CrossRef]
- Lee, G.J.; Vierling, E. A small heat shock protein cooperates with heat shock protein 70 systems to reactivate a heat-denatured protein. Plant Physiol. 2000, 122, 189–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mogk, A.; Deuerling, E.; Vorderwülbecke, S.; Vierling, E.; Bukau, B. Small heat shock proteins, ClpB and the DnaK system form a functional triade in reversing protein aggregation. Mol. Microbiol. 2003, 50, 585–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Min, T.; Hwarari, D.; Li, D.; Movahedi, A.; Yang, L. CRISPR-based genome editing and its applications in woody plants. Int. J. Mol. Sci. 2022, 23, 10175. [Google Scholar] [CrossRef] [PubMed]
- Lin, B.L.; Wang, J.S.; Liu, H.C.; Chen, R.W.; Meyer, Y.; Barakat, A.; Delseny, M. Genomic analysis of the Hsp70 superfamily in Arabidopsis thaliana. Cell Stress Chaperones 2001, 6, 201–208. [Google Scholar] [CrossRef]
- Scharf, K.D.; Siddique, M.; Vierling, E. The expanding family of Arabidopsis thaliana small heat stress proteins and a new family of proteins containing alpha-crystallin domains (Acd proteins). Cell Stress Chaperones 2001, 6, 225–237. [Google Scholar] [CrossRef]
- Hwarari, D.; Guan, Y.; Li, R.; Movahedi, A.; Chen, J.; Yang, L. Comprehensive bioinformatics and expression analysis of TCP transcription factors in Liriodendron chinense reveals putative abiotic stress regulatory roles. Forests 2022, 13, 1401. [Google Scholar] [CrossRef]
- Ke, Y.; Xu, M.; Hwarari, D.; Ahmad, B.; Li, R.; Guan, Y.; Chen, J.; Yang, L. OSCA Genes in Liriodendron chinense: Characterization, evolution and response to abiotic stress. Forests 2022, 13, 1835. [Google Scholar] [CrossRef]
- Li, R.; Radani, Y.; Ahmad, B.; Movahedi, A.; Yang, L. Identification and characteristics of SnRK genes and cold stress-induced expression profiles in Liriodendron chinense. BMC Genom. 2022, 23, 708. [Google Scholar] [CrossRef]
- Wu, W.; Zhu, S.; Xu, L.; Zhu, L.; Wang, D.; Liu, Y.; Liu, S.; Hao, Z.; Lu, Y.; Yang, L.; et al. Genome-wide identification of the Liriodendron chinense WRKY gene family and its diverse roles in response to multiple abiotic stress. BMC Plant Biol. 2022, 22, 25. [Google Scholar] [CrossRef]
- Li, M.; Hwarari, D.; Li, Y.; Ahmad, B.; Min, T.; Zhang, W.; Wang, J.; Yang, L. The bZIP transcription factors in Liriodendron chinense: Genome-wide recognition, characteristics and cold stress response. Front. Plant Sci. 2022, 13, 1035627. [Google Scholar] [CrossRef]
- Li, R.; Ahmad, B.; Hwarari, D.; Li, D.; Lu, Y.; Gao, M.; Chen, J.; Yang, L. Genomic survey and cold-induced expression patterns of bHLH transcription factors in Liriodendron chinense (Hemsl) Sarg. Forests 2022, 13, 518. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Gene Name | Gene ID | Chr | AA | MW | PI | Instability Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| (KDa) | |||||||||
| HSP90 | LchiHSP90-1 | Lchi11721 | Chr4 | 816 | 93.26 | 4.94 | 35.4 | −0.68 | E.R. |
| LchiHSP90-2 | Lchi11722 | Chr4 | 809 | 92.36 | 4.91 | 32.9 | −0.73 | nucl | |
| LchiHSP90-3 | Lchi28127 | Chr8 | 384 | 43.31 | 4.5 | 40.7 | −0.49 | cyto | |
| LchiHSP90-4 | Lchi13166 | Chr11 | 450 | 50.66 | 4.98 | 33.5 | −0.29 | cyto | |
| LchiHSP90-5 | Lchi19077 | Chr14 | 325 | 36.14 | 4.46 | 33.1 | −0.32 | plas | |
| LchiHSP90-6 | Lchi20964 | Chr14 | 792 | 90.22 | 4.99 | 49.9 | −0.49 | chlo | |
| LchiHSP90-7 | Lchi07189 | Chr16 | 654 | 74.29 | 5.06 | 34.7 | −0.59 | chlo | |
| HSP70 | LchiHSP70-1 | Lchi21749 | Chr1 | 786 | 86.84 | 5.23 | 47.0 | −0.33 | cyto |
| LchiHSP70-2 | Lchi01584 | Chr2 | 694 | 74.45 | 5.52 | 35.4 | −0.25 | chlo | |
| LchiHSP70-3 | Lchi18956 | Chr4 | 520 | 57.22 | 5.31 | 34.0 | −0.44 | chlo | |
| LchiHSP70-4 | Lchi18963 | Chr4 | 514 | 56.63 | 5.06 | 37.6 | −0.49 | cyto | |
| LchiHSP70-5 | Lchi16202 | Chr4 | 708 | 75.26 | 5.49 | 31.5 | −0.27 | chlo | |
| LchiHSP70-6 | Lchi06739 | Chr6 | 676 | 73.17 | 5.31 | 35.9 | −0.17 | E.R. | |
| LchiHSP70-7 | Lchi06856 | Chr6 | 884 | 98.26 | 5.42 | 37.7 | −0.41 | E.R. | |
| LchiHSP70-8 | Lchi12038 | Chr6 | 612 | 66.35 | 5.13 | 39.3 | 0.04 | cyto | |
| LchiHSP70-9 | Lchi17471 | Chr6 | 550 | 60.67 | 7.88 | 45.4 | −0.06 | cyto | |
| LchiHSP70-10 | Lchi28754 | Chr10 | 671 | 74.65 | 8.9 | 31.1 | −0.40 | chlo | |
| LchiHSP70-11 | Lchi28755 | Chr10 | 644 | 71.43 | 5.12 | 29.7 | −0.51 | E.R. | |
| LchiHSP70-12 | Lchi18241 | Chr13 | 669 | 71.67 | 5.79 | 37.6 | −0.31 | mito | |
| LchiHSP70-13 | Lchi17994 | Chr16 | 634 | 68.03 | 5.33 | 38.6 | −0.30 | cyto | |
| LchiHSP70-14 | Lchi28433 | Chr17 | 822 | 88.99 | 4.79 | 35.8 | −0.37 | cyto | |
| LchiHSP70-15 | Lchi29184 | Chr17 | 732 | 80.11 | 5.2 | 33.6 | −0.41 | cyto | |
| LchiHSP70-16 | Lchi29183 | Chr17 | 840 | 91.01 | 4.74 | 38.5 | −0.41 | cyto | |
| LchiHSP70-17 | Lchi29180 | Chr17 | 739 | 80.76 | 5.26 | 33.3 | −0.34 | mito | |
| LchiHSP70-18 | Lchi29179 | Chr17 | 813 | 88.72 | 5.05 | 33.3 | −0.28 | cyto | |
| LchiHSP70-19 | Lchi03740 | Chr17 | 655 | 71.64 | 5.19 | 35.3 | −0.38 | cyto | |
| LchiHSP70-20 | Lchi21178 | Chr18 | 648 | 71.05 | 5.09 | 34.0 | −0.43 | cyto | |
| LchiHSP70-21 | Lchi21185 | Chr18 | 539 | 59.06 | 5.32 | 34.6 | −0.46 | cyto | |
| LchiHSP70-22 | Lchi24313 | Chr18 | 539 | 59.64 | 5.13 | 36.9 | −0.51 | cyto | |
| LchiHSP70-23 | Lchi34042 | Scaffold674 | 648 | 71.05 | 5.09 | 34.0 | −0.43 | cyto | |
| HSP20 | LchiHSP20-1 | Lchi21756 | Chr1 | 139 | 16.13 | 5.2 | 48.3 | −0.30 | cyto |
| LchiHSP20-2 | Lchi01684 | Chr2 | 192 | 21.97 | 5.78 | 32.5 | −0.40 | vacu | |
| LchiHSP20-3 | Lchi13753 | Chr2 | 170 | 19.11 | 6.77 | 63.8 | −0.70 | mito | |
| LchiHSP20-4 | Lchi04244 | Chr4 | 314 | 36.03 | 6.23 | 59.1 | −0.66 | cyto | |
| LchiHSP20-5 | Lchi04241 | Chr4 | 257 | 29.61 | 7.86 | 51.4 | −0.76 | cyto | |
| LchiHSP20-6 | Lchi04239 | Chr4 | 183 | 20.27 | 6.19 | 54.3 | −0.45 | cyto | |
| LchiHSP20-7 | Lchi04235 | Chr4 | 176 | 20.12 | 5.4 | 45.6 | −0.68 | cyto | |
| LchiHSP20-8 | Lchi04233 | Chr4 | 161 | 18.50 | 6.78 | 44.4 | −0.70 | cyto | |
| LchiHSP20-9 | Lchi04232 | Chr4 | 192 | 21.80 | 9.2 | 53.6 | −0.62 | cyto | |
| LchiHSP20-10 | Lchi04231 | Chr4 | 251 | 28.97 | 9.24 | 52.9 | −0.78 | cyto | |
| LchiHSP20-11 | Lchi01989 | Chr4 | 181 | 20.53 | 6.87 | 46.1 | −0.49 | extr | |
| LchiHSP20-12 | Lchi02197 | Chr4 | 152 | 17.13 | 5.08 | 41.0 | −0.71 | chlo | |
| LchiHSP20-13 | Lchi09721 | Chr4 | 207 | 23.46 | 5.49 | 66.8 | −0.61 | mito | |
| LchiHSP20-14 | Lchi25013 | Chr5 | 137 | 15.57 | 6.85 | 37.8 | −0.39 | pero | |
| LchiHSP20-15 | Lchi05522 | Chr6 | 226 | 25.99 | 9.01 | 52.8 | −0.75 | mito | |
| LchiHSP20-16 | Lchi11461 | Chr7 | 151 | 16.73 | 6.1 | 52.4 | −0.42 | cyto | |
| LchiHSP20-17 | Lchi20267 | Chr8 | 266 | 29.12 | 9.34 | 37.9 | −0.65 | cyto | |
| LchiHSP20-18 | Lchi20268 | Chr8 | 307 | 35.25 | 9.15 | 35.6 | −0.59 | chlo | |
| LchiHSP20-19 | Lchi20269 | Chr8 | 147 | 16.88 | 9.3 | 43.5 | −0.66 | golg | |
| LchiHSP20-20 | Lchi06582 | Chr8 | 236 | 26.51 | 8.24 | 53.0 | −0.55 | chlo | |
| LchiHSP20-21 | Lchi24852 | Chr11 | 302 | 34.14 | 4.96 | 45.6 | −0.77 | cyto | |
| LchiHSP20-22 | Lchi24857 | Chr11 | 206 | 23.64 | 6.87 | 53.1 | −0.43 | chlo | |
| LchiHSP20-23 | Lchi19505 | Chr15 | 190 | 21.49 | 9 | 44.8 | −0.81 | nucl | |
| LchiHSP20-24 | Lchi03985 | Chr15 | 257 | 28.99 | 6.32 | 58.9 | −0.49 | cyto | |
| LchiHSP20-25 | Lchi03949 | Chr15 | 210 | 23.85 | 9.57 | 57.4 | −0.53 | cyto | |
| LchiHSP20-26 | Lchi03941 | Chr15 | 340 | 38.71 | 6.45 | 58.3 | −0.63 | chlo | |
| LchiHSP20-27 | Lchi28031 | Chr18 | 165 | 18.87 | 6.21 | 43.0 | −0.66 | cyto | |
| LchiHSP20-28 | Lchi31103 | Scaffold97 | 137 | 15.59 | 6.85 | 37.8 | −0.39 | pero | |
| LchiHSP20-29 | Lchi26427 | Scaffold941 | 232 | 26.84 | 5.06 | 65.9 | −0.38 | chlo | |
| LchiHSP20-30 | Lchi31495 | Scaffold1647 | 284 | 31.74 | 9.16 | 42.5 | −0.40 | cyto |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ke, Y.; Xu, M.; Hwarari, D.; Chen, J.; Yang, L. Genomic Survey of Heat Shock Proteins in Liriodendron chinense Provides Insight into Evolution, Characterization, and Functional Diversities. Int. J. Mol. Sci. 2022, 23, 15051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315051
Ke Y, Xu M, Hwarari D, Chen J, Yang L. Genomic Survey of Heat Shock Proteins in Liriodendron chinense Provides Insight into Evolution, Characterization, and Functional Diversities. International Journal of Molecular Sciences. 2022; 23(23):15051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315051
Chicago/Turabian StyleKe, Yongchao, Mingyue Xu, Delight Hwarari, Jinhui Chen, and Liming Yang. 2022. "Genomic Survey of Heat Shock Proteins in Liriodendron chinense Provides Insight into Evolution, Characterization, and Functional Diversities" International Journal of Molecular Sciences 23, no. 23: 15051. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms232315051