
Maresin-1 and Inflammatory Disease

Department of Dermatology, University of Occupational and Environmental Health, 1-1, Iseigaoka, Yahatanishi-Ku, Kitakyushu 807-8555, Fukuoka, Japan
*
Authors to whom correspondence should be addressed.
Int. J. Mol. Sci. 2022, 23(3), 1367; https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031367
Submission received: 9 January 2022
/
Revised: 24 January 2022
/
Accepted: 24 January 2022
/
Published: 25 January 2022
(This article belongs to the Special Issue Fatty Acids and Inflammation: From Biochemistry to Human Evidence)
Abstract
:Inflammation is an essential action to protect the host human body from external, harmful antigens and microorganisms. However, an excessive inflammation reaction sometimes exceeds tissue damage and can disrupt organ functions. Therefore, anti-inflammatory action and resolution mechanisms need to be clarified. Dietary foods are an essential daily lifestyle that influences various human physiological processes and pathological conditions. Especially, omega-3 fatty acids in the diet ameliorate chronic inflammatory skin diseases. Recent studies have identified that omega-3 fatty acid derivatives, such as the resolvin series, showed strong anti-inflammatory actions in various inflammatory diseases. Maresin-1 is a derivative of one of the representative omega-3 fatty acids, i.e., docosahexaenoic acid (DHA), and has shown beneficial action in inflammatory disease models. In this review, we summarize the detailed actions of maresin-1 in immune cells and inflammatory diseases.
1. Introduction
The human body is exposed to various environmental stimuli which drive appropriate host defense reactions to protect against harmful antigens and/or microorganisms [1]. On the one hand, inflammatory responses are essential to drive host defense actions and the remodeling of organ tissues [2]. On the other hand, an inflammatory reaction sometimes exacerbates the inflammatory response, leading to tissue damage and the development of systemic inflammatory diseases [3]. Because inflammation has been implicated in many human diseases, anti-inflammatory agents should be applied to various inflammatory diseases. Steroid or non-steroidal anti-inflammatory drugs (NSAIDs) are currently available medications for the treatment of these inflammatory diseases; however, these medications have disadvantages due to the fact adverse reactions have been associated with them. Therefore, there is a need to develop some safe agents for the treatment of inflammatory diseases.
The daily diet is one of the essential daily lifestyles to sustain the lives of human beings, in which nutrition comprises their body elements and cell components. Among the various nutritional factors, fatty acids are components of cell membranes that regulate cellular signal transduction [4,5]. Recently, research has increased our knowledge of the beneficial actions of fatty acids on human health [6]. Omega-3 fatty acids are abundant in fish oil and are known to have potential benefits in various inflammatory diseases including asthma, psoriasis, inflammatory bowel disease, and rheumatoid arthritis [7]. Furthermore, lipid mediator metabolites derived from omega-3 fatty acids act on various physiological processes or pathological conditions by regulating lipid metabolism, cell signaling, and inflammation [8,9,10,11].
Maresin-1 is one of the highlighted metabolites from eicosapentaenoic acid (EPA) that shows various strong anti-inflammatory actions in inflammatory diseases. In this review, we summarize the actual impacts of maresin-1 on various diseases and discuss the detailed molecular mechanisms of maresin-1 focusing on its anti-inflammatory action.
1.1. The Anti-Inflammatory Actions of Maresin-1
Maresin-1 was first identified in human macrophages and was shown to be a lipid mediator that was active in the process of inflammatory resolution. Studies have shown that human macrophages produce maresin-1 mediated by 14-lipoxylation of DHA and enzymatic hydrolysis from 13S,14S-epoxymaresin [12,13]. Regarding its anti-inflammatory action, maresin-1 has been shown to suppress neutrophil migration [14] and cytokine production by activating CD8+ T cells, CD4+ T helper (Th1) cells, and Th17 cells. Maresin-1 has also been shown to negatively regulate the transcription factors T-bet and Rorc, prevent Th1 and Th17 differentiation, and simultaneously enhance Foxp3+ regulatory T (Treg) cells generation mediated by the GPR32 receptor [15].
Considering the action mechanism of maresin-1, a recent study identified one receptor, i.e., a leucine-rich repeat-containing G-protein-coupled receptor 6 (LGR6), which had the structure of GPCRs and was widely identified in various tissues [16]. Another receptor, i.e., retinoic acid-related orphan receptor α (RORα), has been located in the nucleus [17]. Although these two receptors play roles in the action mechanism of maresin-1, the total detailed molecular mechanisms of the resolution of inflammation, host defense, tissue homeostasis, and wound healing are still unclear.
1.2. DCs/Macrophages
External pathogens and apoptotic cells exacerbate tissue inflammation. However, DCs and macrophages dampen these inflammatory responses by phagocytosis of these pathogens and apoptotic cells for inflammatory resolution. Maresin-1 has been shown to enhance phagocytotic activity in macrophages and promote anti-inflammatory action [18].
Maresin-1 has been shown to promote polarization of CD11c−CD206+ (M2) macrophages and inhibit polarization of CD11c+CD206− (M1) macrophages [19]. The M2 macrophages secreted anti-inflammatory cytokines such as IL-10 and TGF-β, which accelerated tissue remodeling and the clearance of apoptotic debris by phagocytosis [20]. This action of maresin-1 negatively regulated the inflammatory response.
In addition, maresin-1 has been shown to impair inflammatory cytokine production in anti-inflammatory actions. Maresin-1 suppressed the production of IL-1β [21] and TNF-α [22]. In addition, maresin-1 suppressed the production of TNF-α and IL-6 through the suppression of the SIRT1/PGC-1α/PPARγ pathway [23].
1.3. T Cells
Studies have demonstrated that maresin-1 has anti-inflammatory effects on T cells. Maresin-1 suppressed the induction of CD4+ cells, CD8+ cells, and Th17 cells by downregulation of T-bet and Rorc expression [15]. On the contrary, maresin-1 enhanced the induction of Tregs and the production of anti-inflammatory cytokine IL-10. Maresin-1 negatively regulated IL-23 receptor expression on γδ T cells through downregulation of RORγ and internalization of IL-23 receptor [24]. Therefore, maresin-1 suppressed induction of effector cells and induced Treg expansion and anti-inflammatory cytokine production in T cells [15].
1.4. Neutrophils
Neutrophils are involved in the innate immune response and rapidly migrate to infection or injury sites to eliminate invading microorganisms by phagocytotic activity. Maresin-1 has been shown to suppress neutrophil infiltration and decrease the production of CXCL1, which is one of the major chemokines to recruit neutrophils. Apoptosis is a programmed cell death process that prevents the release of cytotoxic contents in cells, and neutrophils contain various abundant cytotoxic substances. Maresin-1 has also been shown to promote apoptosis of neutrophils to induce the resolution of inflammatory response [25].
2. The Inflammatory Actions of Maresin-1 in Inflammatory Diseases
In this section, we introduce the detailed action of maresin-1 in various diseases, especially, neurological disorders, pain, respiratory diseases, diabetes and obesity, kidney disease, liver disease, arthritis, colitis, infectious disease, cardiovascular diseases, and cutaneous diseases.
2.1. Neurology
Neurological actions are essential for human beings to sustain life and the dysregulation of neurological action leads to various severe disorders, such as Alzheimer’s disease. Because there is no radical treatment for these neurological diseases, novel therapeutic or preventive options are desired. Consistently, several studies have identified that maresin-1 had beneficial effects against these neurological diseases. It has been shown, in mouse models, that maresin-1 showed an inhibitory effect on acute neurological damage such as spinal cord injury and cerebral ischemia. In addition, maresin-1 has been shown to impair the disease progress of a chronic degenerative disease, Alzheimer’s disease, by the action mechanism of an inflammatory resolution against the deposition of amyloid-β protein-mediated inflammation [26].
A spinal cord injury accelerates the inflammatory response in neurological tissue and causes delayed remodeling of neurological function. Therefore, early resolution of inflammation associated with a spinal cord injury is required to avoid an excessive inflammatory response in the inflammatory site. Maresin-1 has been shown to be active in the resolution of inflammatory after a spinal cord injury. In an animal model of spinal cord injury, maresin-1 accelerated the resolution of neutrophils and decreased macrophage infiltration at the lesion, which contributed to neurological recovery after a spinal cord injury [27].
In an animal model of brain infarction, maresin-1 also impaired inflammatory reactions in lesions and reduced neurological defects [28]. Epigenetic modification is a powerful gene regulatory mechanism through DNA and/or DNA-binding proteins, such as histone, which modulate open chromatin sites to enhance transcriptional activation [29,30]. Interestingly, maresin-1 has been shown to activate epigenetic modification mediated by silent information regulator 1 (SIRT1 signaling), which is one of the histone deacetylases, to negatively regulate acetylation of nuclear factor kappa B and Bax expression, as well as to reduce downstream proinflammatory cytokines, such as TNF-α and IL-1, leading to a reduction in infarction size and the neurological defects after cerebral ischemia/reperfusion [31].
Epidemiological studies have shown the benefits of DHA intake to reduce the risk of Alzheimer’s disease [32]. Furthermore, oral intake of DHA has been shown to impair disease activity in an animal model of Alzheimer’s disease [33]. These findings suggest that maresin-1, which is a derivative of DHA, should show therapeutic efficacy against Alzheimer’s disease. Consistently, maresin-1 has also shown potential benefits associated with neurological degenerative diseases. For instance, Alzheimer’s disease is a representative neurological degenerative disease, and its incidence is increasing worldwide. Because there are few therapeutic options that have obtained a satisfactory level of clinical use, various research approaches are currently being conducted. In an animal model of Alzheimer’s disease, amyloid-β42 protein was believed to play a central role in the pathogenesis of Alzheimer’s disease by causing an inflammatory response in the amyloid-β42 proteins deposited in the brain. Maresin-1 has been shown to decrease the production of proinflammatory cytokines, such as TNF-α and IL-6, while increasing the secretion of the anti-inflammatory cytokines, IL-2 and IL-10, by the regulation of the amyloid-β42 protein [34,35,36].
2.2. Pain
The therapeutic potential of maresin-1 against pain has been investigated in various animal models. Dorsal root ganglion (DRG) neurons were used to cause neurological pain by capsaicin and vincristine sulfate injection into the hind paw plantar surface, and these stimuli enhanced pains mediated by the transient receptor potential V1 (TRPV1), which was suppressed by maresin-1 treatment [37]. Consistently, maresin-1 inhibited the TRPV1 agonist-induced activation of pain [38].
The beneficial effect of maresin-1 has also been shown in allodynia and thermal hyperalgesia, which relate to nerve hypersensitivity to pain [39]. Maresin-1 has also been shown to be effective in chronic pain, when administered between the L4 and L6 vertebrae of the spinal cord and the analgesic effect was observed for 5 days [40]. Regarding the mechanisms, maresin-1 decreased proinflammatory cytokines (IL-1β, IL-18, and TNFα) and reduced NLRP3 inflammasome, leading to the impairment of cell death and positive activation of NF-κB/p65-mediated inflammation and pain [41]; the mechanical and thermal hypersensitivity enhanced the IL-1β and IL-18 levels and the expression of NLRP3 inflammasome components, which were markedly suppressed by maresin-1 treatment [42].
Polyunsaturated linoleic acid decreases in Ca2+ ions in the cytosol of neurons and astrocytes in an ischemia model [43]. Because maresin-1 is a derivative of polyunsaturated linoleic, maresin-1 might also show therapeutic potential against brain stroke as another possible action mechanism.
2.3. Respiratory Diseases
The anti-inflammatory effects of maresin-1 have been confirmed in various respiratory diseases. Organic dust becomes the cause of chronic airway inflammation, especially obstructive pulmonary disease, due to increased neutrophil infiltration and the production of TNF-α and IL-6. Maresin-1 suppressed proinflammatory cytokine production and intracellular adhesion molecule-1 (ICAM-1) expression in bronchial epithelial cells under organic dust exposure [44]. Maresin-1 significantly decreased bronchoalveolar lavage neutrophil infiltration, and the secretion levels of IL-6, TNF-α, and chemokine C-X-C motif ligand 1 (CXCL1) [45].
High-dose maresin-1 treatment has been shown to impair lung inflammation in an LPS-induced acute lung injury (ALI) mouse model [46]. Infiltrating neutrophils enhance the inflammatory response and release proteolytic enzymes and reactive oxygen species to cause excessive tissue damages. Neutrophil apoptosis was accelerated under LPS-induced ALI, which was impaired by maresin-1 due to the enhancement of caspase-dependent neutrophil apoptosis [25].
Pulmonary fibrosis is a progressive, chronic lung epithelial injury that results in an uncontrolled fibrotic response. Epithelial-to-mesenchymal transition (EMT) is thought to have a pathogenetic role in pulmonary fibrosis by causing epithelial cells to irreversibly shift to a mesenchymal phenotype. TGF-β1 plays a role as a positive driver for EMT in pulmonary fibrosis and enhances collagen synthesis and fibroblast proliferation. The concentration of TGF-β1 in bronchoalveolar lavage fluid and fibrosis markers, such as fibronectin and α-SMA, were suppressed by maresin-1 administration [47]. Maresin-1 has also been shown to inhibit proliferation, migration, and differentiation in fibroblast by suppressing phosphorylation of decapentaplegic homolog 2/3 (Smad2/3) and extracellular-signal-related kinase 1 and 2 (ERK1/2) in a dependent manner [48].
Bronchial asthma is a chronic inflammatory disease of the lower respiratory tract mediated by Th2 allergic reactions, which are essential to exacerbate an inflammatory response by NF-κB-induced ICAM-1 expression in vascular endothelial cells and lung epithelial cells to increase eosinophil adhesion to endothelial cells and to promote Th2 differentiation. Epidemiological studies have suggested that increased dietary intake of fish oil containing omega-3 fatty acids was associated with a reduced risk of asthma [49], suggesting a possible therapeutic efficacy of maresin-1 for asthma. In a mouse model of OVA-induced asthma, maresin-1 markedly suppressed activation of the NF-κB signaling pathway and its downstream cascades, such as COX-2 and ICAM-1 [50]. Maresin-1 enhanced the suppression of innate lymphoid cell type 2 (ILC2) in a TGF-β-dependent manner [51].
Pulmonary ischemia/reperfusion injury causes obstructive bronchiolitis, such as pulmonary thrombolysis and oxidative stress generation plays some part in its pathogenesis. A study has shown that maresin-1 impaired oxidative and antioxidant production leading to protection of the lung tissues [52].
2.4. Diabetes and Obesity
The role of inflammation in the development of type 2 diabetes mellitus and its complications has received increasing attention because the incidence is currently increasing in the world. IL-1β, IL-6, and CRP are considered to be prognostic factors in diabetes [53], and anti-inflammatory lipid mediators such as maresin-1 should prevent the worsening of diabetic retinopathy by converging such adipose inflammation and altering insulin resistance and adipokine secretion [54]. Diet-induced obese mice treated with maresin-1 also exhibited decreased proinflammatory cytokines such as TNF-α and IL-1β [55]. Furthermore, high-fat diet-induced hyperglycemia has been improved by maresin-1 [56] and maresin-1 has enhanced the repair function of macrophages and has also promoted the diabetic wound repair ability [57].
Nonalcoholic fatty liver disease (NAFLD) associated with obesity is a pathological condition caused by endoplasmic reticulum stress and activation of unfolded protein responses. Maresin-1 has been shown to inhibit the endoplasmic reticulum stress of hepatocytes and to enhance the phagocytic activity of Kupffer cells, leading to the protection of hepatocytes from apoptosis [58]. DHA supplementation has been shown to impair metabolic abnormalities in children with NAFLD [59], suggesting a possible therapeutic potential of maresin-1 against NAFLD. Studies have shown that maresin-1 ameliorated hepatic steatosis by inhibiting AMPK/SERCA2b-mediated endoplasmic reticulum stress [60]; impaired hepatic lipidosis by inhibiting AMPK activation and inducing autophagy [61]; and suppressed liver injury by increasing the expression and transcriptional activity of RORα [62].
2.5. Kidney Disease
Leukocyte-mediated inflammation also plays a pathogenetic role in acute kidney injury. One study reported that maresin-1 influenced the survival of neutrophils and subsequently prevented kidney injury. The macrophages produced 14S,21R-dihydroxydocosahexaenoic acid (14S,21R-diHDHA), which repaired the vascular endothelium and contributed to the protection of kidney function [63].
Inflammation and fibrosis are also important pathologies in diabetic nephropathy. In a mouse model of diabetic nephropathy, maresin-1 exerted a protective effect on glomerular mesangial cells by decreasing the expression of ROS, NLPR3, caspase-1, and IL-1β, which are responsible for the development of diabetic nephropathy [64].
Inflammation and oxidative stresses are also involved in ischemia/reperfusion-induced renal injury; TLR4-mediated inflammatory response and the signal pathway mediated by ERK, JNK, and P38 MAPK play an important role. Maresin-1 treatment has been shown to decrease the expression levels of ERK, JNK, and P38 MAPK [65].
2.6. Liver
The effects of maresin-1 in hepatic injury have also been investigated in a mouse model of acute hepatic injury. Maresin-1 inhibited reactive oxygen species and inflammatory cytokines and chemokines, such as IL-6, IL-1β, and monocyte chemotaxis protein-1 (MCP-1), and suppressed carbon tetrachloride-induced liver injury [66]. In another acute liver injury mouse model induced by concanavalin A, maresin-1 also impaired liver injury by reducing hepatocytes apoptosis while increasing apoptosis of mouse macrophages, in addition to reducing ROS in macrophages [67].
Hepatic ischemia-reperfusion injury causes liver dysfunction after liver surgery. In a rat model, maresin-1 impaired hepatic injury by activating hepatocyte and promoting nuclear localization of Nrf-2, leading to a decrease in NF-κB activity [68].
In another study, the hepatoprotective effect of maresin-1 was abrogated by pretreatment with Boc2 (lipoxin A4 receptor antagonist), and the hepatoprotective effect of maresin-1 was further reversed by inhibition of Akt. Thus, maresin-1 protected the liver from hepatic ischemia-reperfusion injury mediated by the ALXR/Akt signaling pathway [69].
2.7. Arthritis
Several studies have shown the therapeutic potential of maresin-1 against arthritis. The concentration of maresin-1 in synovial fluid of rheumatoid arthritis patients has been shown to be related to disease activity [70], suggesting that maresin-1 might have a protective role in the development of rheumatoid arthritis. The serum level of maresin-1 was also lower in inactive rheumatoid arthritis than active rheumatoid arthritis. An inverse correlation was observed between the FoxP3/RORc ratio and the disease activity score 28, which is a measure of disease activity in rheumatoid arthritis. Furthermore, maresin-1 has been shown to suppress inflammatory response, in a rheumatoid arthritis animal model [71]. Maresin-1 also showed therapeutic potential by elevating intra-articular lavage fluid in a treadmill-loaded mouse model of osteoarthritis. Maresin-1 treatment has been reported to enhance type II collagen in cartilage and to decrease MMP13 in the synovium, mediated by the PI3k/Akt and NF-κB p65 pathways [72].
2.8. Colitis
The beneficial effects of EPA and DHA on inflammatory bowel disease have been reported. In a colitis animal model induced by dextran sulfate sodium (DSS) and 2,4,6-trinitrobenzene sulfonic acid, maresin-1 suppressed disease activity in colitis and improved weight loss by decreasing IL-1β, TNF-α, IL-6, and IFN-γ in the acute phase. Maresin-1 also decreased neutrophil migration and ROS production mediated by the NF-κB pathway [73]. Maresin-1 negatively regulated the toll-like receptor 4 (TLR4)-mediated NF-κB pathway [74].
2.9. Infectious Diseases
Acute inflammation induced by infections can cause excessive tissue damage, and therefore, the resolution of an acute inflammatory response during the early phase is important for infectious disease regulation. Maresin-1 has also been shown to regulate the inflammatory immune response during an Escherichia coli infection [75]. Furthermore, in a sepsis animal model induced by intestinal ligation and puncture, maresin-1 decreased the serum concentration of LPS, promoted bacterial clearance, and protected critical organ functions in addition to improved survival [76]. Furthermore, mitochondrial dysfunction leads to increased ROS production in sepsis, and maresin-1 has been reported to increase mitochondrial membrane integrity by retaining adenosine triphosphate content and decreasing ROS production [77].
During severe sepsis, acute kidney injury is the most severe complication, which has been shown to be impaired by maresin-1 treatment. Neutrophil infiltration is inhibited by maresin-1 via the NF-κB/STAT3/MAPK pathway and negatively modulated proinflammatory cytokine levels [78]. Maresin-1 can also impair myocardial infarction during sepsis and reduce the levels of LDH and CK, leading to the improvement of cardiac function through promoting M2 macrophages differentiation [79].
2.10. Cardiovascular Diseases
Cerebrovascular diseases are life-threatening diseases with inflammation responses that can result in vascular damage. Maresin-1 treatment could impair these responses through anti-inflammatory effects in vascular endothelial cells. Maresin-1 has been shown to inhibit TNF-α-induced monocyte adhesion and ROS generation in vascular endothelial cells and smooth muscle cells by causing upregulation of cAMP and downregulation of the transcription factor NF-κβ [80]. Vascular injury activates remodeling of vascular endothelial cells by inflammation and sometimes causes neointima formation leading to re-occlusion in blood vessels. Systemic administration of maresin-1 has been shown to reduce neointima formation, and therefore, should suppress the re-occlusion of blood vessels [81].
There is agreement that chronic inflammation is one of the causes of atherosclerosis. Profiling of the aortic qualities of mediators in Apoe-deficient mice fed a high-fat diet showed increased inflammatory lipid mediators leukotriene B4 and prostaglandin E2, and decreased omega-3 polyunsaturated fatty acid lipid mediators, such as resolvin D2 (RvD2) and maresin-1. Maresin-1 inhibited the progression of atherosclerosis by suppressing necrosis in the atherosclerosis site and macrophage accumulation, and increasing the fibrous coat thickness of smooth muscle cells [82].
Smooth muscle cell-specific TGF-β2 receptor-deficient mice were used to induce localized abdominal aortic aneurysms to confirm the therapeutic potential of maresin-1; maresin-1 treatment suppressed the growth of aortic aneurysms mediated by LGR6 receptor signaling which was responsible for TGF-β2 and MMP2 activity in macrophage-apoptotic smooth muscle cell crosstalk [83].
2.11. Cutaneous Diseases
Psoriasis is an inflammatory skin disease that is characterized by scaly erythematous plaques, which can also drive the inflammatory immune reaction to systemic organs. Patients with psoriasis have been shown to have a lower amount of omega-3 fatty acids as compared with healthy controls [84], indicating the importance of a diet with regular intake of foods that provide omega-3 fatty acids. A study reported that consistent fish oil supplement intake impaired psoriatic skin inflammation in approximately 80% of patients with psoriasis [85]. There are several benefits of omega-3 polyunsaturated fatty acid metabolites in inflammatory skin diseases. Maresin-1 has also been known to have anti-inflammatory action in psoriasis [24]. A topical application of maresin-1 showed anti-inflammatory effects in a mouse model of psoriasis induced by imiquimod. Maresin-1 inhibited the production of IL-17A by γδTCRmid+ and CD4+ cells in the skin by downmodulation of IL-23 receptor (IL-23R) expression in clathrin-dependent internalization mechanisms in γδTCRmid+ and CD4+ cells [24]. Therefore, topical maresin-1 could become a therapeutic option for the treatment of IL-17-mediated other inflammatory diseases.
There are various types of skin inflammations. For instance, Th1 is involved in the pathogenesis of contact dermatitis and Th2 mediates atopic dermatitis. The pathogenesis of alopecia areata is associated with changes in Th1, Th2, and Th17. Therefore, maresin-1 should show anti-inflammatory actions in various skin diseases.
3. Conclusions
In this review, we summarized the therapeutic potential of maresin-1 in various inflammatory diseases. Maresin-1 could be an alternative therapeutic option to overcome the disadvantages of current anti-inflammatory agents. Furthermore, by discovering the effects on other types of cells, such as keratinocytes, the actions of maresin-1 could be clarified. Because the incidence of inflammatory diseases is currently increasing, future basic research and clinical trials for maresin-1 should provide beneficial information on its use for the daily clinical treatment of inflammatory diseases.
Author Contributions
N.S.-S. and Y.S. wrote and revised manuscript; M.N. conducted a critical review of this manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
Y.S. received a basic research grant from the Japanese Dermatology Association (Novartis Pharma donated) and a research grant from the Lydia O’Leary Memorial Pias Dermatological Foundation.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef] [PubMed]
- Reilkoff, R.A.; Bucala, R.; Herzog, E.L. Fibrocytes: Emerging effector cells in chronic inflammation. Nat. Rev. Immunol. 2011, 11, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Hawkes, J.E.; Chan, T.C.; Krueger, J.G. Psoriasis pathogenesis and the development of novel targeted immune therapies. J. Allergy Clin. Immunol. 2017, 140, 645–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueharaguchi, Y.; Honda, T.; Kusuba, N.; Hanakawa, S.; Adachi, A.; Sawada, Y.; Otsuka, A.; Kitoh, A.; Dainichi, T.; Egawa, G.; et al. Thromboxane A2 facilitates IL-17A production from Vγ4+ γδ T cells and promotes psoriatic dermatitis in mice. J. Allergy Clin. Immunol. 2018, 142, 680–683.e2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sawada, Y.; Honda, T.; Nakamizo, S.; Nakajima, S.; Nonomura, Y.; Otsuka, A.; Egawa, G.; Yoshimoto, T.; Nakamura, M.; Narumiya, S.; et al. Prostaglandin E(2) (PGE(2))-EP2 signaling negatively regulates murine atopic dermatitis-like skin inflammation by suppressing thymic stromal lymphopoietin expression. J. Allergy Clin. Immunol. 2019, 144, 1265–1273.e9. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Saito-Sasaki, N.; Nakamura, M. Omega 3 Fatty Acid and Skin Diseases. Front. Immunol. 2020, 11, 623052. [Google Scholar] [CrossRef]
- Horrobin, D.F. Low prevalences of coronary heart disease (CHD), psoriasis, asthma and rheumatoid arthritis in Eskimos: Are they caused by high dietary intake of eicosapentaenoic acid (EPA), a genetic variation of essential fatty acid (EFA) metabolism or a combination of both? Med. Hypotheses 1987, 22, 421–428. [Google Scholar]
- Serhan, C.N.; Haeggström, J.Z.; Leslie, C.C. Lipid mediator networks in cell signaling: Update and impact of cytokines. FASEB J. 1996, 10, 1147–1158. [Google Scholar] [CrossRef]
- Levy, B.D.; Clish, C.B.; Schmidt, B.; Gronert, K.; Serhan, C.N. Lipid mediator class switching during acute inflammation: Signals in resolution. Nat. Immunol. 2001, 2, 612–619. [Google Scholar] [CrossRef]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [Green Version]
- Sawada, Y.; Honda, T.; Hanakawa, S.; Nakamizo, S.; Murata, T.; Ueharaguchi-Tanada, Y.; Ono, S.; Amano, W.; Nakajima, S.; Egawa, G.; et al. Resolvin E1 inhibits dendritic cell migration in the skin and attenuates contact hypersensitivity responses. J. Exp. Med. 2015, 212, 1921–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalli, J.; Zhu, M.; Vlasenko, N.A.; Deng, B.; Haeggström, J.Z.; Petasis, N.A.; Serhan, C.N. The novel 13S,14S-epoxy-maresin is converted by human macrophages to maresin 1 (MaR1), inhibits leukotriene A4 hydrolase (LTA4H), and shifts macrophage phenotype. FASEB J. 2013, 27, 2573–2583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, B.; Wang, C.W.; Arnardottir, H.H.; Li, Y.; Cheng, C.Y.; Dalli, J.; Serhan, C.N. Maresin biosynthesis and identification of maresin 2, a new anti-inflammatory and pro-resolving mediator from human macrophages. PLoS ONE 2014, 9, e102362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdulnour, R.E.; Dalli, J.; Colby, J.K.; Krishnamoorthy, N.; Timmons, J.Y.; Tan, S.H.; Colas, R.A.; Petasis, N.A.; Serhan, C.N.; Levy, B.D. Maresin 1 biosynthesis during platelet-neutrophil interactions is organ-protective. Proc. Natl. Acad. Sci. USA 2014, 111, 16526–16531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiurchiù, V.; Leuti, A.; Dalli, J.; Jacobsson, A.; Battistini, L.; Maccarrone, M.; Serhan, C.N. Proresolving lipid mediators resolvin D1, resolvin D2, and maresin 1 are critical in modulating T cell responses. Sci. Transl. Med. 2016, 8, 353ra111. [Google Scholar] [CrossRef] [Green Version]
- Chiang, N.; Libreros, S.; Norris, P.C.; de la Rosa, X.; Serhan, C.N. Maresin 1 activates LGR6 receptor promoting phagocyte immunoresolvent functions. J. Clin. Investig. 2019, 129, 5294–5311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Im, D.S. Maresin-1 resolution with RORα and LGR6. Prog. Lipid Res. 2020, 78, 101034. [Google Scholar] [CrossRef]
- Zhu, M.; Wang, X.; Hjorth, E.; Colas, R.A.; Schroeder, L.; Granholm, A.C.; Serhan, C.N.; Schultzberg, M. Pro-Resolving Lipid Mediators Improve Neuronal Survival and Increase Aβ42 Phagocytosis. Mol. Neurobiol. 2016, 53, 2733–2749. [Google Scholar] [CrossRef]
- Qiao, N.; Lin, Y.; Wang, Z.; Chen, J.Y.; Ge, Y.Y.; Yao, S.L.; Gong, J. Maresin1 Promotes M2 Macrophage Polarization Through Peroxisome Proliferator-Activated Receptor-γ Activation to Expedite Resolution of Acute Lung Injury. J. Surg. Res. 2020, 256, 584–594. [Google Scholar] [CrossRef]
- Bi, Y.; Chen, J.; Hu, F.; Liu, J.; Li, M.; Zhao, L. M2 Macrophages as a Potential Target for Antiatherosclerosis Treatment. Neural Plast. 2019, 2019, 6724903. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Tao, K.; Zhang, P.; Chen, X.; Sun, X.; Li, R. Maresin 1 protects against lipopolysaccharide/d-galactosamine-induced acute liver injury by inhibiting macrophage pyroptosis and inflammatory response. Biochem. Pharmacol. 2022, 195, 114863. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, A.; Sarabia, C.; Torres, M.; Juárez, E. Resolvin D1 (RvD1) and maresin 1 (Mar1) contribute to human macrophage control of M. tuberculosis infection while resolving inflammation. Int. Immunopharmacol. 2019, 74, 105694. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xu, R.L.; He, P.; Chen, R. MAR1 suppresses inflammatory response in LPS-induced RAW 264.7 macrophages and human primary peripheral blood mononuclear cells via the SIRT1/PGC-1α/PPAR-γ pathway. J. Inflamm. 2021, 18, 8. [Google Scholar] [CrossRef] [PubMed]
- Saito-Sasaki, N.; Sawada, Y.; Mashima, E.; Yamaguchi, T.; Ohmori, S.; Yoshioka, H.; Haruyama, S.; Okada, E.; Nakamura, M. Maresin-1 suppresses imiquimod-induced skin inflammation by regulating IL-23 receptor expression. Sci. Rep. 2018, 8, 5522. [Google Scholar] [CrossRef]
- Gong, J.; Liu, H.; Wu, J.; Qi, H.; Wu, Z.Y.; Shu, H.Q.; Li, H.B.; Chen, L.; Wang, Y.X.; Li, B.; et al. Maresin 1 prevents lipopolysaccharide-induced neutrophil survival and accelerates resolution of acute lung injury. Shock 2015, 44, 371–380. [Google Scholar] [CrossRef]
- Wang, Y.; Leppert, A.; Tan, S.; van der Gaag, B.; Li, N.; Schultzberg, M.; Hjorth, E. Maresin 1 attenuates pro-inflammatory activation induced by β-amyloid and stimulates its uptake. J. Cell. Mol. Med. 2021, 25, 434–447. [Google Scholar] [CrossRef]
- Francos-Quijorna, I.; Santos-Nogueira, E.; Gronert, K.; Sullivan, A.B.; Kopp, M.A.; Brommer, B.; David, S.; Schwab, J.M.; López-Vales, R. Maresin 1 Promotes Inflammatory Resolution, Neuroprotection, and Functional Neurological Recovery after Spinal Cord Injury. J. Neurosci. 2017, 37, 11731–11743. [Google Scholar] [CrossRef] [Green Version]
- Xian, W.; Li, T.; Li, L.; Hu, L.; Cao, J. Maresin 1 attenuates the inflammatory response and mitochondrial damage in mice with cerebral ischemia/reperfusion in a SIRT1-dependent manner. Brain Res. 2019, 1711, 83–90. [Google Scholar] [CrossRef]
- Sawada, Y.; Gallo, R.L. Role of Epigenetics in the Regulation of Immune Functions of the Skin. J. Investig. Dermatol. 2021, 141, 1157–1166. [Google Scholar] [CrossRef]
- Sawada, Y.; Nakatsuji, T.; Dokoshi, T.; Kulkarni, N.N.; Liggins, M.C.; Sen, G.; Gallo, R.L. Cutaneous innate immune tolerance is mediated by epigenetic control of MAP2K3 by HDAC8/9. Sci. Immunol. 2021, 6, eabe1935. [Google Scholar] [CrossRef]
- Xian, W.; Wu, Y.; Xiong, W.; Li, L.; Li, T.; Pan, S.; Song, L.; Hu, L.; Pei, L.; Yao, S.; et al. The pro-resolving lipid mediator Maresin 1 protects against cerebral ischemia/reperfusion injury by attenuating the pro-inflammatory response. Biochem. Biophys. Res. Commun. 2016, 472, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Barberger-Gateau, P.; Letenneur, L.; Deschamps, V.; Pérès, K.; Dartigues, J.F.; Renaud, S. Fish, meat, and risk of dementia: Cohort study. BMJ 2002, 325, 932–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, G.P.; Calon, F.; Morihara, T.; Yang, F.; Teter, B.; Ubeda, O.; Salem, N., Jr.; Frautschy, S.A.; Cole, G.M. A diet enriched with the omega-3 fatty acid docosahexaenoic acid reduces amyloid burden in an aged Alzheimer mouse model. J. Neurosci. 2005, 25, 3032–3040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.; Xu, G.; Newton, P.T.; Chagin, A.S.; Mkrtchian, S.; Carlström, M.; Zhang, X.M.; Harris, R.A.; Cooter, M.; Berger, M.; et al. Maresin 1 attenuates neuroinflammation in a mouse model of perioperative neurocognitive disorders. Br. J. Anaesth. 2019, 122, 350–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, P.; Wang, X.; Wang, S.; Wei, Y.; Feng, J.; Zhu, M. Maresin 1 Improves Cognitive Decline and Ameliorates Inflammation in a Mouse Model of Alzheimer’s Disease. Front. Cell. Neurosci. 2019, 13, 466. [Google Scholar] [CrossRef] [Green Version]
- Miyazawa, K.; Fukunaga, H.; Tatewaki, Y.; Takano, Y.; Yamamoto, S.; Mutoh, T.; Taki, Y. Alzheimer’s Disease and Specialized Pro-Resolving Lipid Mediators: Do MaR1, RvD1, and NPD1 Show Promise for Prevention and Treatment? Int. J. Mol. Sci. 2020, 21, 5783. [Google Scholar] [CrossRef]
- Serhan, C.N.; Dalli, J.; Karamnov, S.; Choi, A.; Park, C.K.; Xu, Z.Z.; Ji, R.R.; Zhu, M.; Petasis, N.A. Macrophage proresolving mediator maresin 1 stimulates tissue regeneration and controls pain. FASEB J. 2012, 26, 1755–1765. [Google Scholar] [CrossRef] [Green Version]
- Park, C.K. Maresin 1 Inhibits TRPV1 in Temporomandibular Joint-Related Trigeminal Nociceptive Neurons and TMJ Inflammation-Induced Synaptic Plasticity in the Trigeminal Nucleus. Mediat. Inflamm. 2015, 2015, 275126. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Tang, C.; Tai, L.W.; Ouyang, Y.; Li, N.; Hu, Z.; Chen, X. Pro-resolving mediator maresin 1 ameliorates pain hypersensitivity in a rat spinal nerve ligation model of neuropathic pain. J. Pain Res. 2018, 11, 1511–1519. [Google Scholar] [CrossRef] [Green Version]
- Fattori, V.; Pinho-Ribeiro, F.A.; Staurengo-Ferrari, L.; Borghi, S.M.; Rossaneis, A.C.; Casagrande, R.; Verri, W.A., Jr. The specialised pro-resolving lipid mediator maresin 1 reduces inflammatory pain with a long-lasting analgesic effect. Br. J. Pharmacol. 2019, 176, 1728–1744. [Google Scholar] [CrossRef]
- Wang, Y.H.; Li, Y.; Wang, J.N.; Zhao, Q.X.; Jin, J.; Wen, S.; Wang, S.C.; Sun, T. Maresin 1 Attenuates Radicular Pain Through the Inhibition of NLRP3 Inflammasome-Induced Pyroptosis via NF-κB Signaling. Front. Neurosci. 2020, 14, 831. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Li, Y.; Wang, J.N.; Zhao, Q.X.; Wen, S.; Wang, S.C.; Sun, T. A Novel Mechanism of Specialized Proresolving Lipid Mediators Mitigating Radicular Pain: The Negative Interaction with NLRP3 Inflammasome. Neurochem. Res. 2020, 45, 1860–1869. [Google Scholar] [CrossRef] [PubMed]
- Turovsky, E.A.; Varlamova, E.G.; Gudkov, S.V.; Plotnikov, E.Y. The Protective Mechanism of Deuterated Linoleic Acid Involves the Activation of the Ca2+ Signaling System of Astrocytes in Ischemia In Vitro. Int. J. Mol. Sci. 2021, 22, 13216. [Google Scholar] [CrossRef] [PubMed]
- Nordgren, T.M.; Heires, A.J.; Wyatt, T.A.; Poole, J.A.; LeVan, T.D.; Cerutis, D.R.; Romberger, D.J. Maresin-1 reduces the pro-inflammatory response of bronchial epithelial cells to organic dust. Respir. Res. 2013, 14, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordgren, T.M.; Bauer, C.D.; Heires, A.J.; Poole, J.A.; Wyatt, T.A.; West, W.W.; Romberger, D.J. Maresin-1 reduces airway inflammation associated with acute and repetitive exposures to organic dust. Transl. Res. 2015, 166, 57–69. [Google Scholar] [CrossRef] [Green Version]
- Gong, J.; Wu, Z.Y.; Qi, H.; Chen, L.; Li, H.B.; Li, B.; Yao, C.Y.; Wang, Y.X.; Wu, J.; Yuan, S.Y.; et al. Maresin 1 mitigates LPS-induced acute lung injury in mice. Br. J. Pharmacol. 2014, 171, 3539–3550. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, R.; Chen, L.; Tan, W.; Sun, Z.; Xia, H.; Li, B.; Yu, Y.; Gong, J.; Tang, M.; et al. Maresin 1 Inhibits Epithelial-to-Mesenchymal Transition In Vitro and Attenuates Bleomycin Induced Lung Fibrosis In Vivo. Shock 2015, 44, 496–502. [Google Scholar] [CrossRef]
- Sun, Q.; Wu, Y.; Zhao, F.; Wang, J. Maresin 1 inhibits transforming growth factor-β1-induced proliferation, migration and differentiation in human lung fibroblasts. Mol. Med. Rep. 2017, 16, 1523–1529. [Google Scholar] [CrossRef] [Green Version]
- Mihrshahi, S.; Peat, J.K.; Webb, K.; Oddy, W.; Marks, G.B.; Mellis, C.M. Effect of omega-3 fatty acid concentrations in plasma on symptoms of asthma at 18 months of age. Pediatric Allergy Immunol. 2004, 15, 517–522. [Google Scholar] [CrossRef]
- Ou, G.; Liu, Q.; Yu, C.; Chen, X.; Zhang, W.; Chen, Y.; Wang, T.; Luo, Y.; Jiang, G.; Zhu, M.; et al. The Protective Effects of Maresin 1 in the OVA-Induced Asthma Mouse Model. Mediat. Inflamm. 2021, 2021, 4131420. [Google Scholar] [CrossRef]
- Krishnamoorthy, N.; Burkett, P.R.; Dalli, J.; Abdulnour, R.E.; Colas, R.; Ramon, S.; Phipps, R.P.; Petasis, N.A.; Kuchroo, V.K.; Serhan, C.N.; et al. Cutting edge: Maresin-1 engages regulatory T cells to limit type 2 innate lymphoid cell activation and promote resolution of lung inflammation. J. Immunol. 2015, 194, 863–867. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Wu, Y.; Zhao, F.; Wang, J. Maresin 1 Ameliorates Lung Ischemia/Reperfusion Injury by Suppressing Oxidative Stress via Activation of the Nrf-2-Mediated HO-1 Signaling Pathway. Oxidative Med. Cell. Longev. 2017, 2017, 9634803. [Google Scholar] [CrossRef] [PubMed]
- Navarro, J.F.; Mora, C. Diabetes, inflammation, proinflammatory cytokines, and diabetic nephropathy. Sci. World J. 2006, 6, 908–917. [Google Scholar] [CrossRef] [PubMed]
- Clària, J.; Nguyen, B.T.; Madenci, A.L.; Ozaki, C.K.; Serhan, C.N. Diversity of lipid mediators in human adipose tissue depots. Am. J. Physiol. Cell Physiol. 2013, 304, C1141–C1149. [Google Scholar] [CrossRef]
- León, I.C.; Quesada-Vázquez, S.; Sáinz, N.; Guruceaga, E.; Escoté, X.; Moreno-Aliaga, M.J. Effects of Maresin 1 (MaR1) on Colonic Inflammation and Gut Dysbiosis in Diet-Induced Obese Mice. Microorganisms 2020, 8, 1156. [Google Scholar] [CrossRef]
- Martínez-Fernández, L.; González-Muniesa, P.; Sáinz, N.; Escoté, X.; Martínez, J.A.; Arbones-Mainar, J.M.; Moreno-Aliaga, M.J. Maresin 1 regulates insulin signaling in human adipocytes as well as in adipose tissue and muscle of lean and obese mice. J. Physiol. Biochem. 2021, 77, 167–173. [Google Scholar] [CrossRef]
- Hong, S.; Lu, Y.; Tian, H.; Alapure, B.V.; Wang, Q.; Bunnell, B.A.; Laborde, J.M. Maresin-like lipid mediators are produced by leukocytes and platelets and rescue reparative function of diabetes-impaired macrophages. Chem. Biol. 2014, 21, 1318–1329. [Google Scholar] [CrossRef] [Green Version]
- Rius, B.; Duran-Güell, M.; Flores-Costa, R.; López-Vicario, C.; Lopategi, A.; Alcaraz-Quiles, J.; Casulleras, M.; Lozano, J.J.; Titos, E.; Clària, J. The specialized proresolving lipid mediator maresin 1 protects hepatocytes from lipotoxic and hypoxia-induced endoplasmic reticulum stress. FASEB J. 2017, 31, 5384–5398. [Google Scholar] [CrossRef] [Green Version]
- Pacifico, L.; Bonci, E.; Di Martino, M.; Versacci, P.; Andreoli, G.; Silvestri, L.M.; Chiesa, C. A double-blind, placebo-controlled randomized trial to evaluate the efficacy of docosahexaenoic acid supplementation on hepatic fat and associated cardiovascular risk factors in overweight children with nonalcoholic fatty liver disease. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 734–741. [Google Scholar] [CrossRef]
- Jung, T.W.; Kim, H.C.; Abd El-Aty, A.M.; Jeong, J.H. Maresin 1 attenuates NAFLD by suppression of endoplasmic reticulum stress via AMPK-SERCA2b pathway. J. Biol. Chem. 2018, 293, 3981–3988. [Google Scholar] [CrossRef] [Green Version]
- Laiglesia, L.M.; Lorente-Cebrián, S.; Martínez-Fernández, L.; Sáinz, N.; Prieto-Hontoria, P.L.; Burrell, M.A.; Rodríguez-Ortigosa, C.M.; Martínez, J.A.; Moreno-Aliaga, M.J. Maresin 1 mitigates liver steatosis in ob/ob and diet-induced obese mice. Int. J. Obes. 2018, 42, 572–579. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.H.; Shin, K.O.; Kim, J.Y.; Khadka, D.B.; Kim, H.J.; Lee, Y.M.; Cho, W.J.; Cha, J.Y.; Lee, B.J.; Lee, M.O. A maresin 1/RORα/12-lipoxygenase autoregulatory circuit prevents inflammation and progression of nonalcoholic steatohepatitis. J. Clin. Investig. 2019, 129, 1684–1698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.; Lu, Y. Omega-3 fatty acid-derived resolvins and protectins in inflammation resolution and leukocyte functions: Targeting novel lipid mediator pathways in mitigation of acute kidney injury. Front. Immunol. 2013, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, S.; Gao, C.; Long, Y.; Huang, W.; Chen, J.; Fan, F.; Jiang, C.; Xu, Y. Maresin 1 Mitigates High Glucose-Induced Mouse Glomerular Mesangial Cell Injury by Inhibiting Inflammation and Fibrosis. Mediat. Inflamm. 2017, 2017, 2438247. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Y.; Wu, Y.; Zhao, H.; Sun, H.; Gao, S. Maresin 1 mitigates renal ischemia/reperfusion injury in mice via inhibition of the TLR4/MAPK/NF-κB pathways and activation of the Nrf2 pathway. Drug Des. Dev. Ther. 2019, 13, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Wang, Y.; Zhao, E.; Wu, K.; Li, W.; Shi, L.; Wang, D.; Xie, G.; Yin, Y.; Deng, M.; et al. Maresin 1, a Proresolving Lipid Mediator, Mitigates Carbon Tetrachloride-Induced Liver Injury in Mice. Oxidative Med. Cell. Longev. 2016, 2016, 9203716. [Google Scholar] [CrossRef]
- Zhang, P.; Yin, Y.; Wang, T.; Li, W.; Li, C.; Zeng, X.; Yang, W.; Zhang, R.; Tang, Y.; Shi, L.; et al. Maresin 1 mitigates concanavalin A-induced acute liver injury in mice by inhibiting ROS-mediated activation of NF-κB signaling. Free Radic. Biol. Med. 2020, 147, 23–36. [Google Scholar] [CrossRef]
- Soto, G.; Rodríguez, M.J.; Fuentealba, R.; Treuer, A.V.; Castillo, I.; González, D.R.; Zúñiga-Hernández, J. Maresin 1, a Proresolving Lipid Mediator, Ameliorates Liver Ischemia-Reperfusion Injury and Stimulates Hepatocyte Proliferation in Sprague-Dawley Rats. Int. J. Mol. Sci. 2020, 21, 540. [Google Scholar] [CrossRef] [Green Version]
- Tang, D.; Fu, G.; Li, W.; Sun, P.; Loughran, P.A.; Deng, M.; Scott, M.J.; Billiar, T.R. Maresin 1 protects the liver against ischemia/reperfusion injury via the ALXR/Akt signaling pathway. Mol. Med. 2021, 27, 18. [Google Scholar] [CrossRef]
- Giera, M.; Ioan-Facsinay, A.; Toes, R.; Gao, F.; Dalli, J.; Deelder, A.M.; Serhan, C.N.; Mayboroda, O.A. Lipid and lipid mediator profiling of human synovial fluid in rheumatoid arthritis patients by means of LC-MS/MS. Biochim. Biophys. Acta 2012, 1821, 1415–1424. [Google Scholar] [CrossRef] [Green Version]
- Jin, S.; Chen, H.; Li, Y.; Zhong, H.; Sun, W.; Wang, J.; Zhang, T.; Ma, J.; Yan, S.; Zhang, J.; et al. Maresin 1 improves the Treg/Th17 imbalance in rheumatoid arthritis through miR-21. Ann. Rheum. Dis. 2018, 77, 1644–1652. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Feng, X.; Zhang, H.; Wei, Y.; Yang, Y.; Tian, Y.; Bai, L. Maresin-1 suppresses IL-1β-induced MMP-13 secretion by activating the PI3K/AKT pathway and inhibiting the NF-κB pathway in synovioblasts of an osteoarthritis rat model with treadmill exercise. Connect. Tissue Res. 2021, 62, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Inflammatory bowel disease as a disorder of an imbalance between pro- and anti-inflammatory molecules and deficiency of resolution bioactive lipids. Lipids Health Dis. 2016, 15, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, S.; Li, P.; Zhao, H.; Li, X. Maresin 1 alleviates dextran sulfate sodium-induced ulcerative colitis by regulating NRF2 and TLR4/NF-kB signaling pathway. Int. Immunopharmacol. 2020, 78, 106018. [Google Scholar] [CrossRef] [PubMed]
- Colas, R.A.; Dalli, J.; Chiang, N.; Vlasakov, I.; Sanger, J.M.; Riley, I.R.; Serhan, C.N. Identification and Actions of the Maresin 1 Metabolome in Infectious Inflammation. J. Immunol. 2016, 197, 4444–4452. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wang, Y.; Ma, Z.; Ma, M.; Wang, D.; Xie, G.; Yin, Y.; Zhang, P.; Tao, K. Maresin 1 Mitigates Inflammatory Response and Protects Mice from Sepsis. Mediat. Inflamm. 2016, 2016, 3798465. [Google Scholar] [CrossRef]
- Gu, J.; Luo, L.; Wang, Q.; Yan, S.; Lin, J.; Li, D.; Cao, B.; Mei, H.; Ying, B.; Bin, L.; et al. Maresin 1 attenuates mitochondrial dysfunction through the ALX/cAMP/ROS pathway in the cecal ligation and puncture mouse model and sepsis patients. Lab. Investig. 2018, 98, 715–733. [Google Scholar] [CrossRef]
- Sun, S.; Wang, J.; Wang, J.; Wang, F.; Yao, S.; Xia, H. Maresin 1 Mitigates Sepsis-Associated Acute Kidney Injury in Mice via Inhibition of the NF-κB/STAT3/MAPK Pathways. Front. Pharmacol. 2019, 10, 1323. [Google Scholar] [CrossRef] [Green Version]
- Li, D.; Wang, M.; Ye, J.; Zhang, J.; Xu, Y.; Wang, Z.; Zhao, M.; Ye, D.; Wan, J. Maresin 1 alleviates the inflammatory response, reduces oxidative stress and protects against cardiac injury in LPS-induced mice. Life Sci. 2021, 277, 119467. [Google Scholar] [CrossRef]
- Chatterjee, A.; Sharma, A.; Chen, M.; Toy, R.; Mottola, G.; Conte, M.S. The pro-resolving lipid mediator maresin 1 (MaR1) attenuates inflammatory signaling pathways in vascular smooth muscle and endothelial cells. PLoS ONE 2014, 9, e113480. [Google Scholar] [CrossRef] [Green Version]
- Akagi, D.; Chen, M.; Toy, R.; Chatterjee, A.; Conte, M.S. Systemic delivery of proresolving lipid mediators resolvin D2 and maresin 1 attenuates intimal hyperplasia in mice. FASEB J. 2015, 29, 2504–2513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viola, J.R.; Lemnitzer, P.; Jansen, Y.; Csaba, G.; Winter, C.; Neideck, C.; Silvestre-Roig, C.; Dittmar, G.; Döring, Y.; Drechsler, M.; et al. Resolving Lipid Mediators Maresin 1 and Resolvin D2 Prevent Atheroprogression in Mice. Circ. Res. 2016, 119, 1030–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elder, C.T.; Filiberto, A.C.; Su, G.; Ladd, Z.; Leroy, V.; Pruitt, E.Y.; Lu, G.; Jiang, Z.; Sharma, A.K.; Upchurch, G.R., Jr. Maresin 1 activates LGR6 signaling to inhibit smooth muscle cell activation and attenuate murine abdominal aortic aneurysm formation. FASEB J. 2021, 35, e21780. [Google Scholar] [CrossRef] [PubMed]
- Barrea, L.; Macchia, P.E.; Tarantino, G.; Di Somma, C.; Pane, E.; Balato, N.; Napolitano, M.; Colao, A.; Savastano, S. Nutrition: A key environmental dietary factor in clinical severity and cardio-metabolic risk in psoriatic male patients evaluated by 7-day food-frequency questionnaire. J. Transl. Med. 2015, 13, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kragballe, K. Dietary supplementation with a combination of n-3 and n-6 fatty acids (super gamma-oil marine) improves psoriasis. Acta Derm. Venereol. 1989, 69, 265–268. [Google Scholar] [PubMed]
Figure 1.
The detailed action of maresin-1 in immune cells. ↑ upregulated, ↓ downregulated.


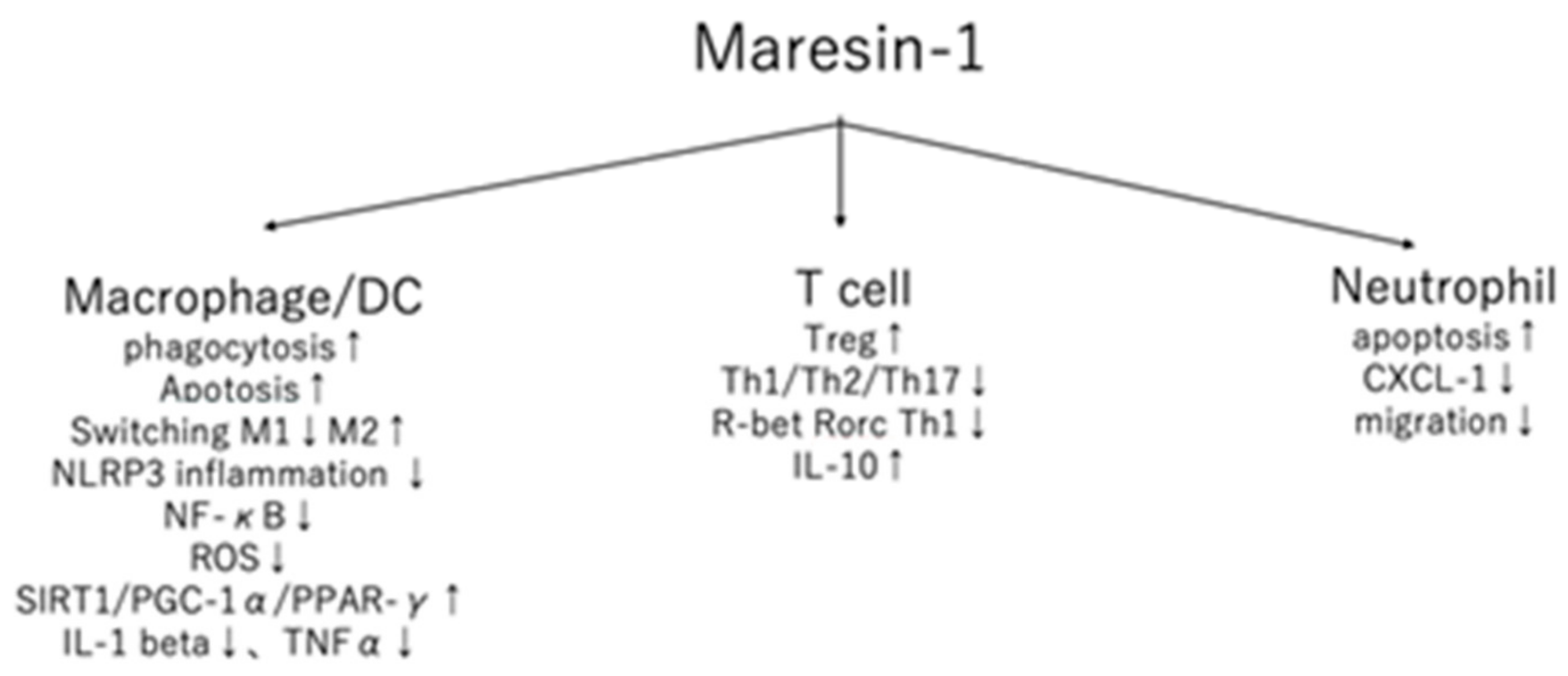{kind=link}
Table 1.
The detailed action of maresin-1 in immune cells. ↑ upregulated, ↓ downregulated.
| Macrophage DC | Phagocytosis ↑ M2 Polarization IL-1β ↓, TNF-α ↓, IL-6 ↓, ROS ↓ |
| T cells | Th1, Th2, Th17 induction ↓ Treg ↑ |
| Neutrophil | Apoptosis ↑ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Saito-Sasaki, N.; Sawada, Y.; Nakamura, M. Maresin-1 and Inflammatory Disease. Int. J. Mol. Sci. 2022, 23, 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031367
AMA Style
Saito-Sasaki N, Sawada Y, Nakamura M. Maresin-1 and Inflammatory Disease. International Journal of Molecular Sciences. 2022; 23(3):1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031367
Chicago/Turabian StyleSaito-Sasaki, Natsuko, Yu Sawada, and Motonobu Nakamura. 2022. "Maresin-1 and Inflammatory Disease" International Journal of Molecular Sciences 23, no. 3: 1367. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23031367
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.