
Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats

 , , and
, , and 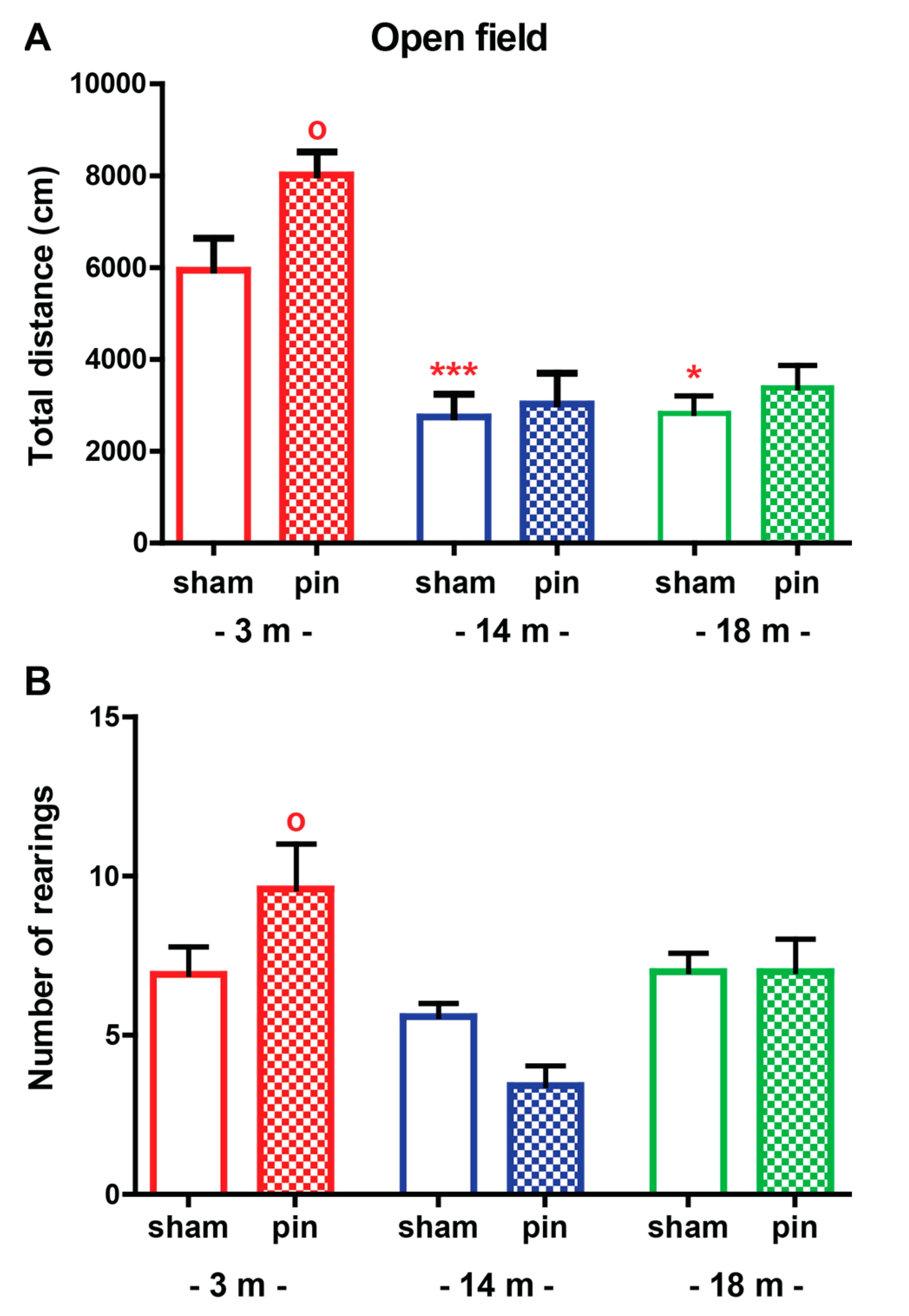{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effect of Melatonin Deficit on Emotional Behavior in Young Adult, Mature and Old Rats
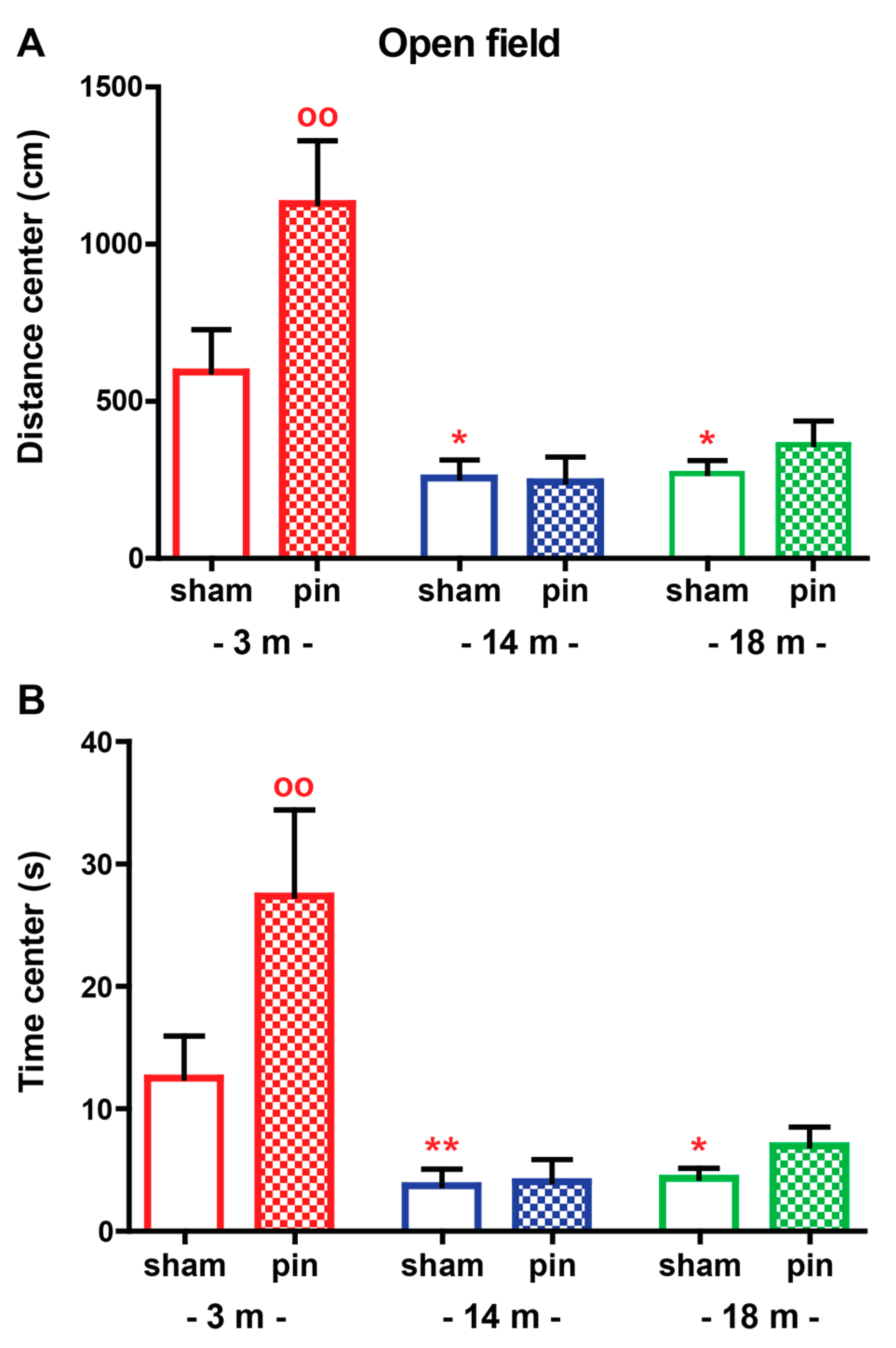
2.1.1. Open Field (OF) Test
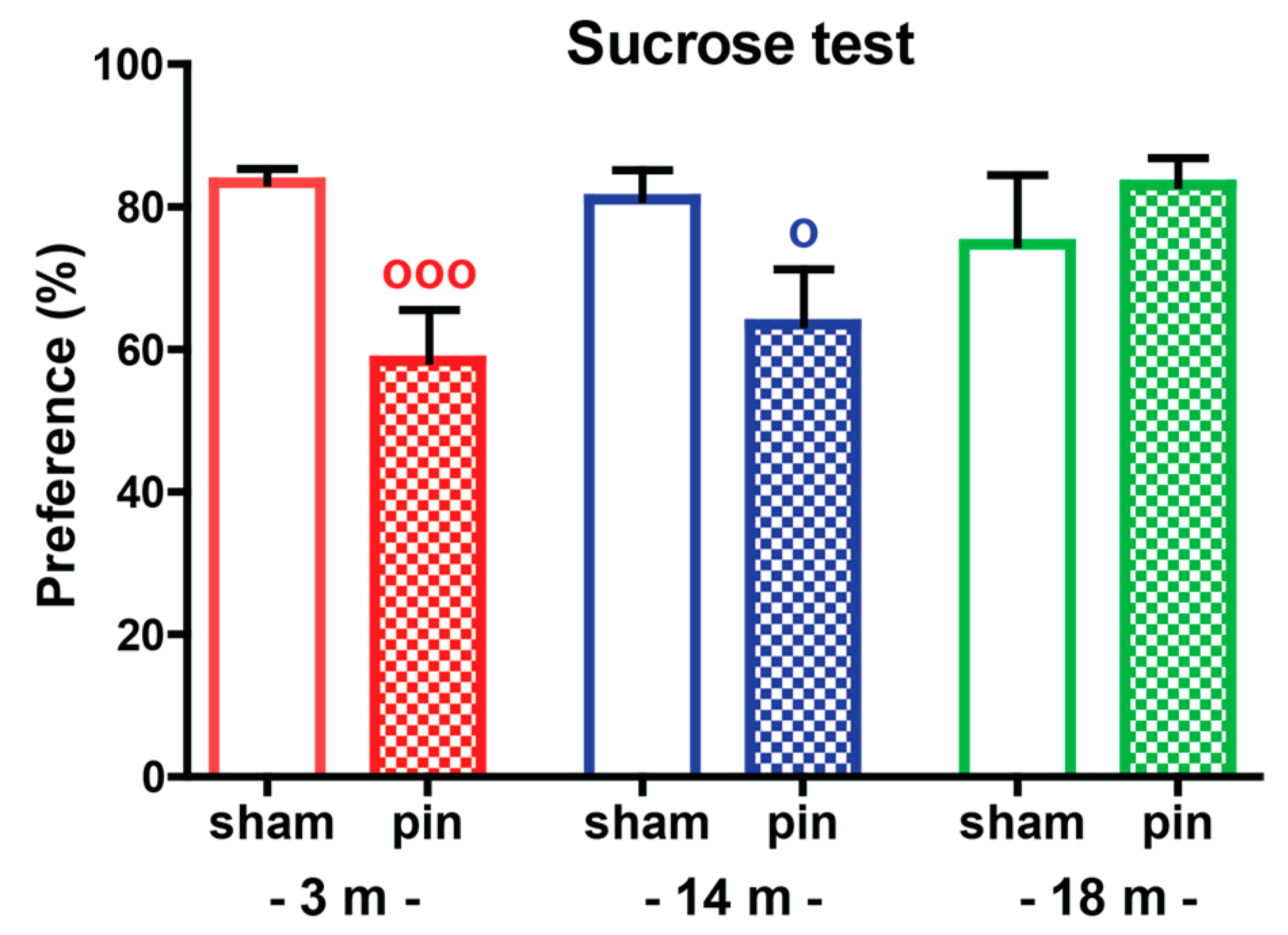
2.1.2. Sucrose Preference Test (SPT)
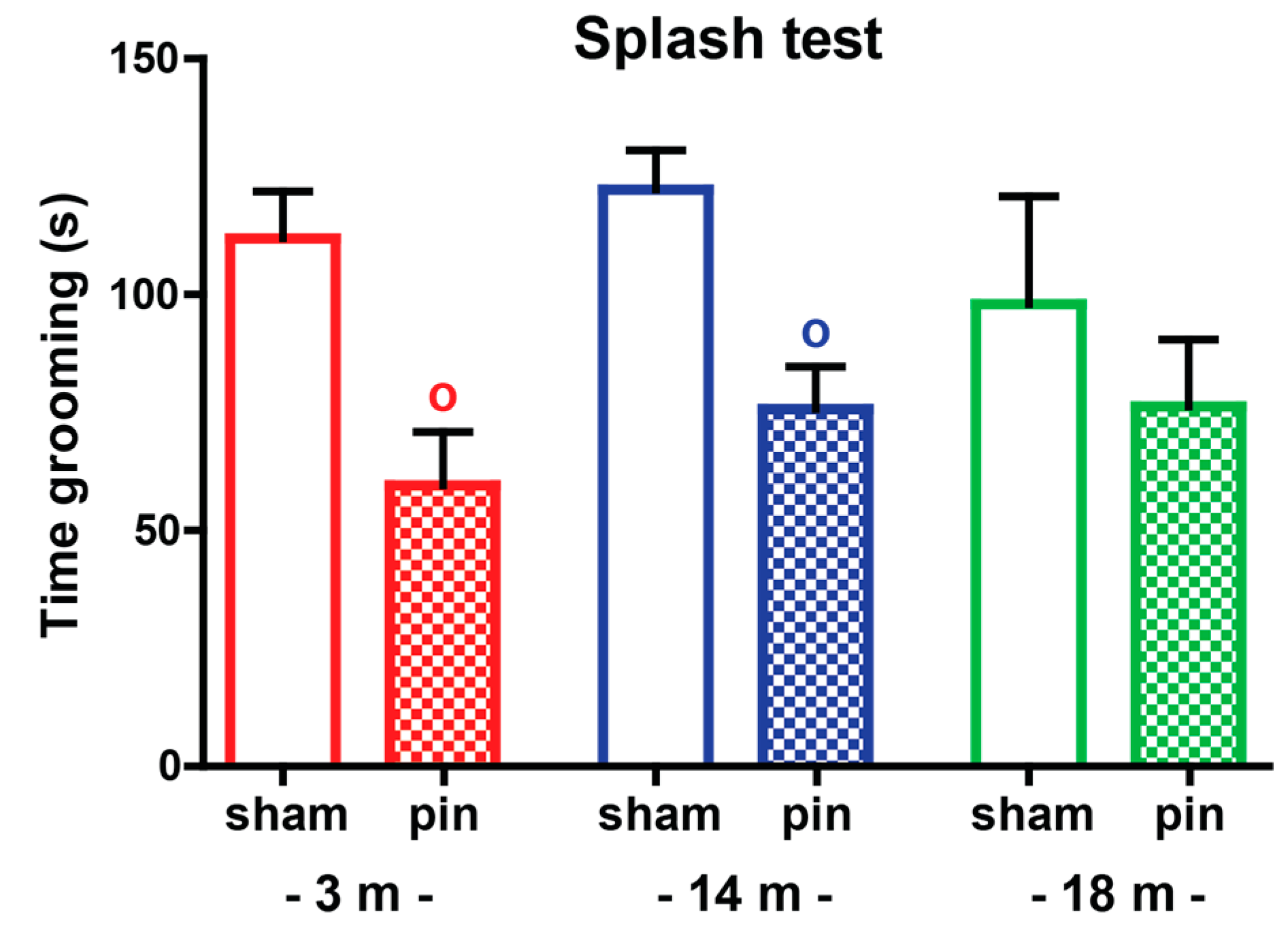
2.1.3. Splash Test
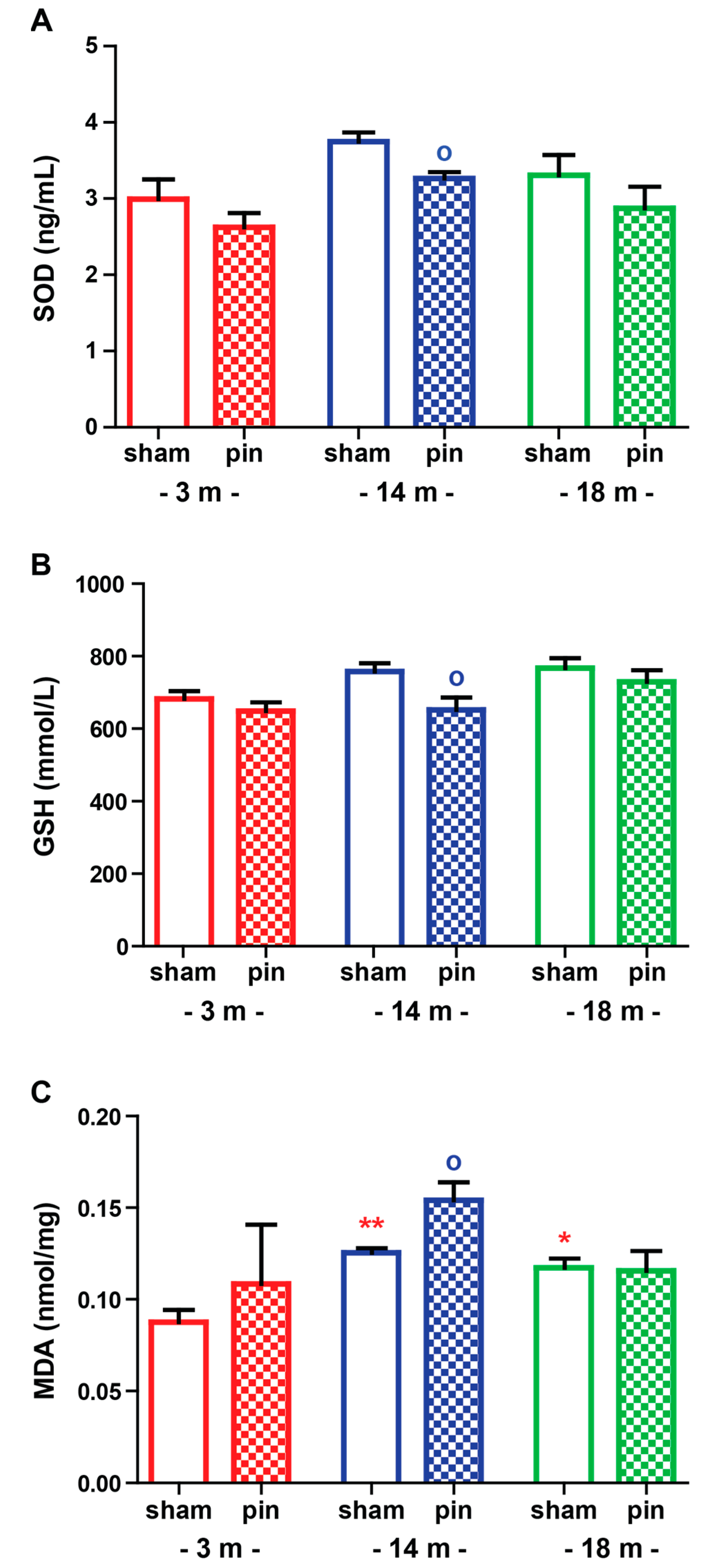
2.2. The Effect of Melatonin Deficit on Oxidative Stress Markers in the Hippocampus of Young Adult, Mature and Old Rats
2.2.1. Superoxide Dismutase (SOD)
2.2.2. Glutathione (GSH)
2.2.3. Malondialdehyde (MDA)
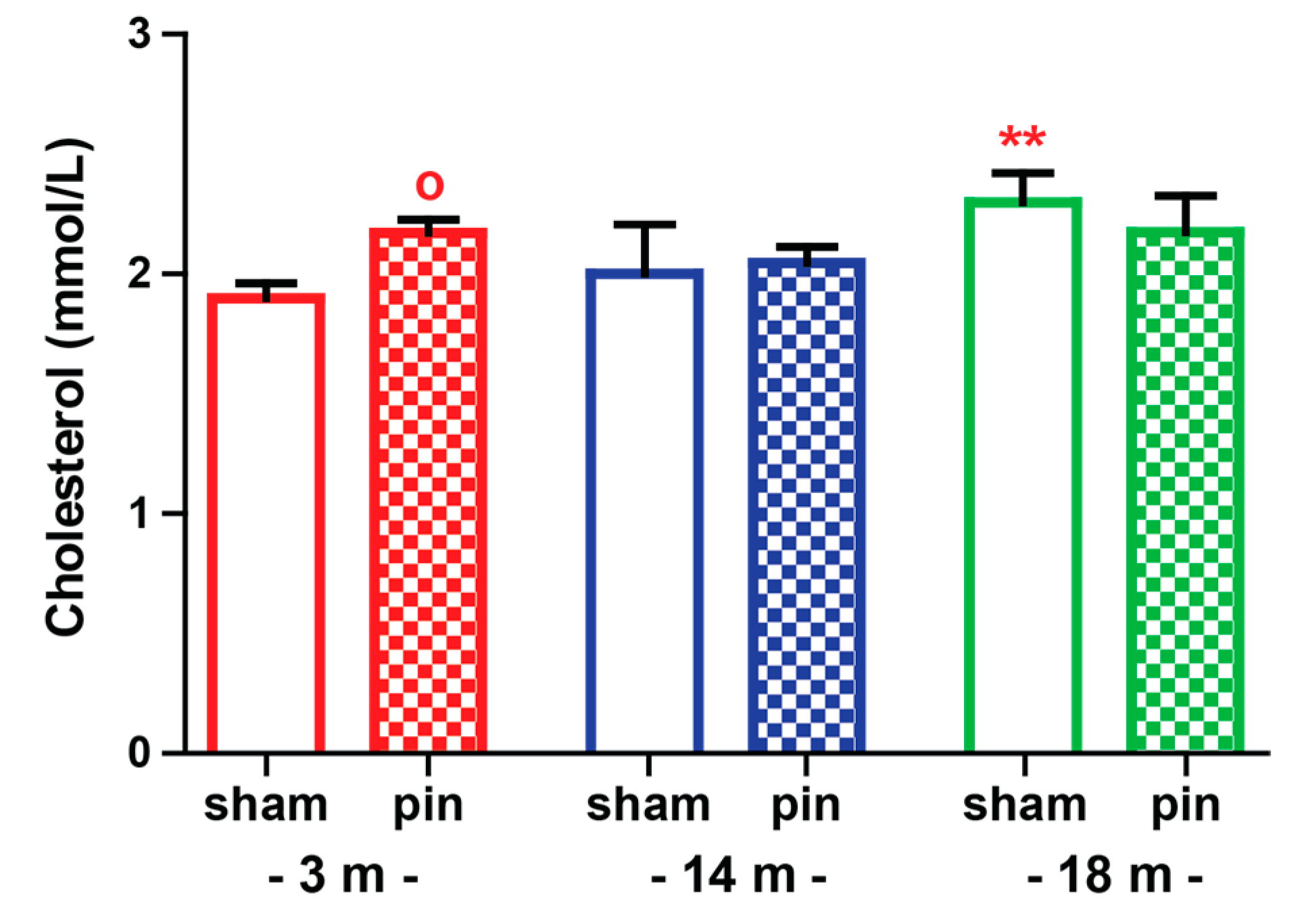
2.3. The Effect of Melatonin Deficit on Cholesterol Levels in Serum of Young Adult, Mature and Old Rats
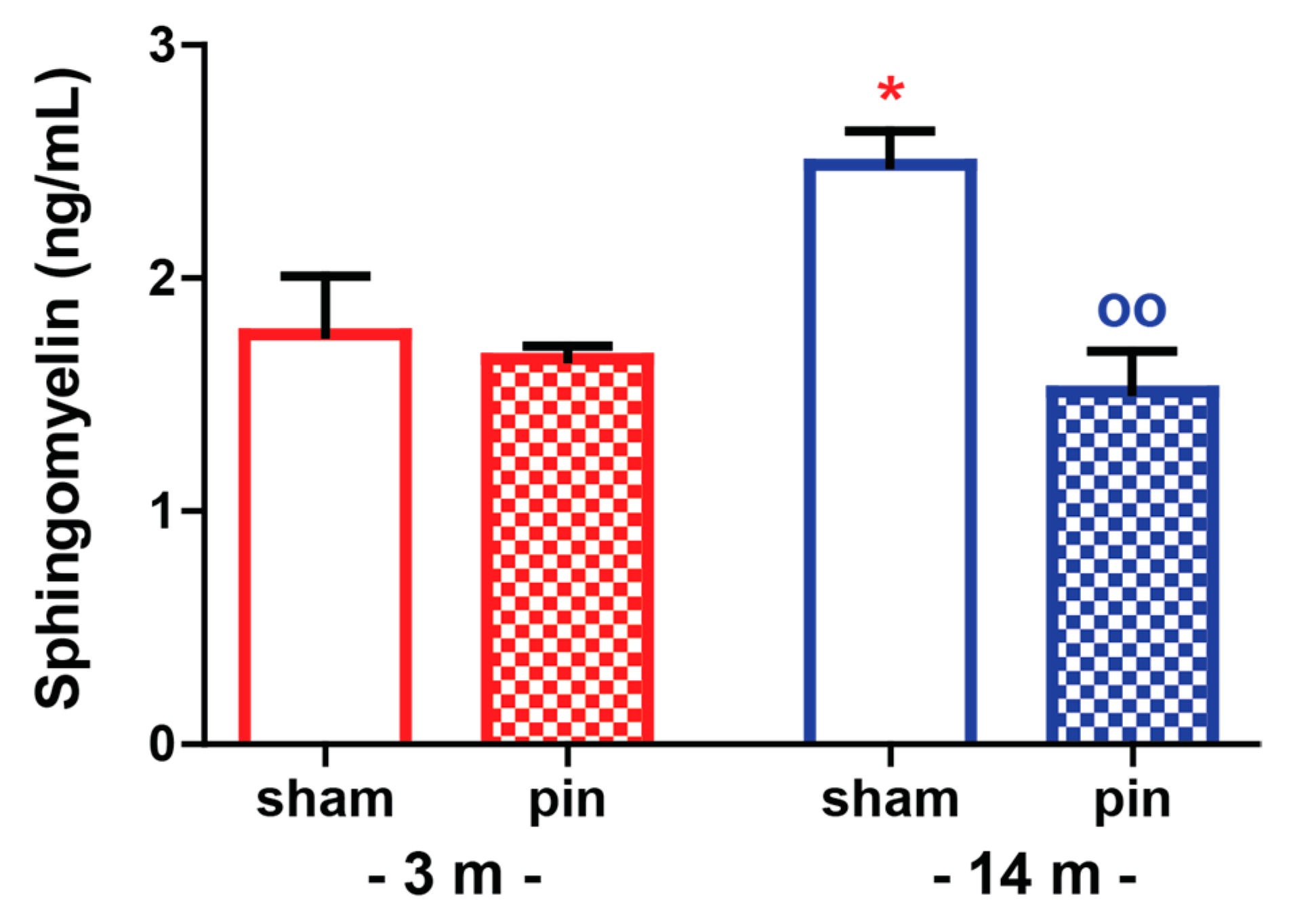
2.4. The Effect of Melatonin Deficit on Sphingomyelin Levels in the Hippocampus of Young Adult and Mature Rats
3. Discussion
4. Materials and Methods
4.1. Animals
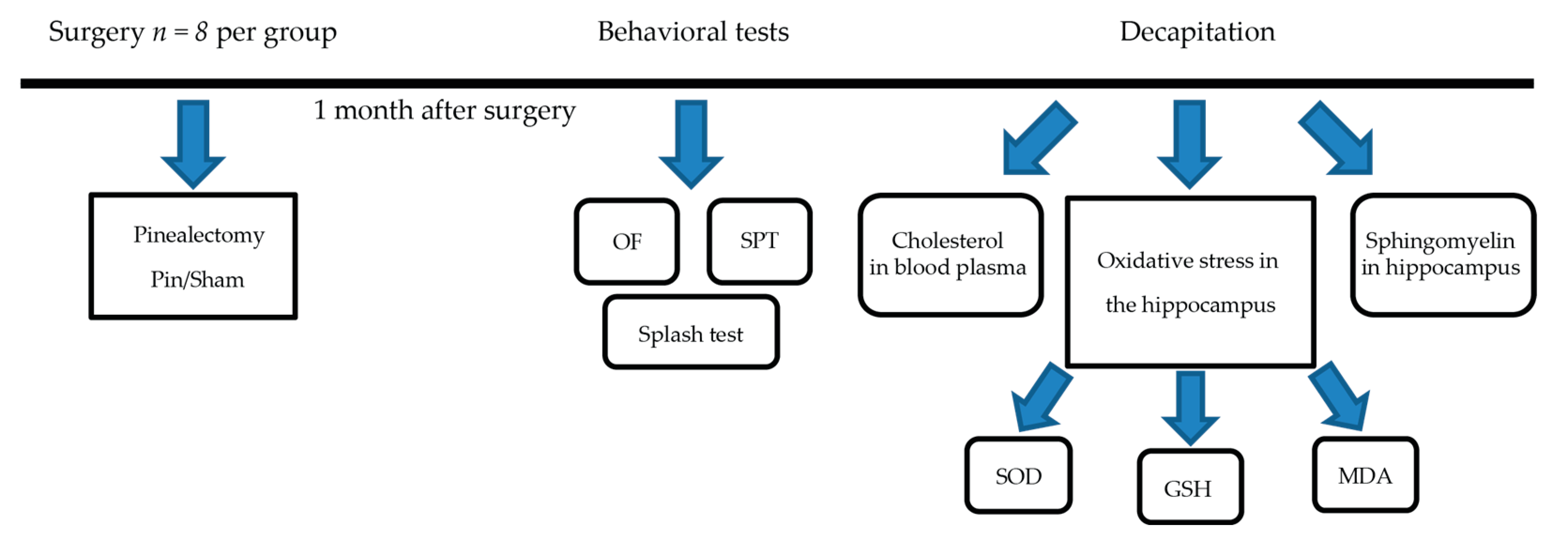
4.2. Experimental Schedule and Surgical Procedures
4.3. Behavioral Tests
4.3.1. Open Field Test
4.3.2. Sucrose Preference Test
4.3.3. Splash Test
4.4. Oxidative Stress in the Hippocampus
4.4.1. Superoxide Dismutase (SOD)
4.4.2. Glutathione
4.4.3. Malondialdehyde
4.5. Sphyngomielin Level in the Hippocampus and Cholesterol Level in Plasma
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sun, N.; Youle, R.J.; Finkel, T. The Mitochondrial Basis of Aging. Mol. Cell. 2016, 61, 654–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardeland, R.; Fuhrberg, B. Ubiquitous melatonin—Presence and effects in unicells, plants and animals. Trends Comp. Biochem. Physiol. 1996, 2, 25–45. [Google Scholar]
- Cutler, R.; Kelly, G.; Storie, J.K.; Pedersen, W.A.; Tammara, A.; Hatanpa, K.; Troncoso, J.C.; Mattson, M.P. Involvement of oxidative stress-induced abnormalities in ceramide and cholesterol metabolism in brain aging and Alzheimer’s disease. Proc. Nat. Acad. Sci. USA 2004, 101, 2070–2075. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monette, S.; Gómez, L.A.; Moreau, R.; Dunn, K.; Butler, J.; Finlay, L.A.; Michels, A.J.; Shay, K.; Smith, E.; Hagen, T.M. (R)-α-Lipoic Acid Treatment Restores Ceramide Balance in Aging Rat Cardiac Mitochondria. Pharmacol. Res. 2011, 63, 23–29. [Google Scholar] [CrossRef] [Green Version]
- Brunkhorst, R.; Pfeilschifter, W.; Rajkovic, N.; Pfefferb, M.; Fischerb, C.; Werner, H.; Christoffersen, C.; Trautmann, S.; Thomas, D.; Pfeilschifter, J.; et al. Diurnal regulation of sphingolipids in blood. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2019, 1864, 304–311. [Google Scholar] [CrossRef]
- Hoehn, R.; Monse, M.; Pohl, E.; Wranik, S.; Wilker, B.; Keitsch, S.; Soddemann, M.; Kornhuber, J.; Kohnen, M.; Edwards, M.J.; et al. Melatonin acts as an antidepressant by inhibition of the acid sphingomyelinase/Ceramide System. Neurosignals 2016, 24, 48–58. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.; Dotti, C.G.; Ledesma, M.D. Brain cholesterol in normal and pathological aging. Biochim. Biophys. Acta 2010, 1801, 934–944. [Google Scholar] [CrossRef]
- Karolczak, K.; Watala, C. The Mystery behind the Pineal Gland: Melatonin Affects the Metabolism of Cholesterol. Oxidative Med. Cell. Longev. 2019, 2019, 4531865. [Google Scholar] [CrossRef]
- Pierpaoli, W. The pineal gland: A circadian or seasonal aging clock? Aging Clin. Exp. Res. 1991, 3, 99–101. [Google Scholar] [CrossRef]
- Pierpaoli, W. The Pineal Gland as Ontogenetic Scanner of Reproduction, Immunity, and Aging the Aging Clock. Ann. N. Y. Acad. Sci. 1994, 741, 46–49. [Google Scholar] [CrossRef]
- Pierpaoli, W.; Bulian, D. The Pineal Aging and Death Program. I. Grafting of Old Pineals in Young Mice Accelerates Their Aging. J. Anti-Aging Med. 2001, 4, 31–37. [Google Scholar] [CrossRef]
- Pierpaoli, W. The Pineal Aging and Death Program: Life Prolongation in Pre-aging Pinealectomized Mice. Ann. N. Y. Acad. Sci. 2005, 1057, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Emerich, D.F.; McDermott, P.; Krueger, P.; Banks, M.; Zhao, J.; Marszalowski, J.; Frydel, B.; Winn, S.R.; Sanberg, P.R. Locomotion of aged rats: Relationship to neurochemical but not morphological changes in nigrostriatal dopaminergic neurons. Brain Res. Bull. 1993, 32, 477–486. [Google Scholar] [CrossRef]
- Marshall, J.F.; Joyce, J.N. Basal ganglia dopamine receptor autoradiography and age-related movement disorders. Ann. N. Y. Acad. Sci. 1988, 515, 215–225. [Google Scholar] [CrossRef]
- Popoli, P.; Reggio, R.; Pèzzola, A.; Fuxe, K.; Ferré, S. Adenosine A1 and A2A receptorantagonists stimulate motor activity: Evidence for an increased effectiveness in aged rats. Neurosci. Lett. 1998, 251, 201–204. [Google Scholar] [CrossRef]
- Hofecker, G.; Kment, A.; Niedermüller, H. Die motorische Aktivität als Altersparameter der Ratte [The motor activity as an age parameter of the rat (authors transl)]. Aktuelle Gerontol. 1978, 8, 271–279. [Google Scholar]
- FIood, D.G.; Coleman, P.D. Neuron numbers and sizes in aging brain: Comparisons of Human, monkey, and rodent data. Neurobiof. Aging 1988, 9, 453–463. [Google Scholar] [CrossRef]
- Murray, A.M.; Waddington, J.L. Age-related changes in the regulation of behavior by D-l: D-2 dopamine receptor interactions. Neurobiol. Aging 1991, 12, 431–435. [Google Scholar] [CrossRef]
- Sudakov, S.K.; Alekseeva, E.V.; Nazarova, G.A.; Bashkatova, V.G. Age-Related Individual Behavioural Characteristics of Adult Wistar Rats. Animals 2021, 11, 2282. [Google Scholar] [CrossRef]
- Boguszewski, P.; Zagrodzka, J. Emotional changes related to age in rats—A behavioral analysis. Behav. Brain Res. 2002, 133, 323–332. [Google Scholar] [CrossRef]
- Meyza, K.Z.; Boguszewski, P.M.; Nikolaev, E.; Zagrodzka, J. Age increases anxiety and reactivity of the fear/anxiety circuit in Lewis rats. Behav. Brain Res. 2011, 225, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Singhal, G.; Morgan, J.; Jawahar, M.C.; Corrigan, F.; Jaehne, E.J.; Toben, C.; Breen, J.; Pederson, S.M.; Manavis, J.; Hannan, A.J.; et al. Effects of aging on the motor, cognitive and affective behaviors, neuroimmune responses and hippocampal gene expression. Behav. Brain Res. 2020, 383, 112501. [Google Scholar] [CrossRef] [PubMed]
- Turner, R.C.; Seminerio, M.J.; Naser, Z.J.; Martin, S.J.; Matsumoto, R.R.; Rosen, C.L.; Hube, J.D. Effects of aging on behavioral assessment performance: Implications for clinically relevant models of neurological disease. J. Neurosurg. 2012, 117, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Torras-Garcia, M.; Costa-Miserachs, D.; Coll-Andreu, M.; Portell-Cortés, I. Decreased anxiety levels related to aging. Exp. Brain Res. 2005, 164, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Gokdemir, O.; Cetinkaya, C.; Gumus, H.; Aksu, I.; Kiray, M.; Ates, M.; Kiray, A.; Baykara, B.; Baykara, B.; Sisman, A.R.; et al. The effect of exercise on anxiety- and depression-like behavior of aged rats. Biotech. Histochem. 2020, 95, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Nenchovska, Z.; Atanasova, D.; Atanasova, M.; Kortenska, L.; Stefanova, M.; Alova, L.; Lazarov, N. Consequences of long-term treatment with agomelatine on depressive-like behavior and neurobiological abnormalities in pinealectomized rats. Behav. Brain Res. 2016, 302, 11–28. [Google Scholar] [CrossRef]
- Parker, G. A case for reprising and redefining melancholia. Can. J. Psychiatry 2013, 58, 183–189. [Google Scholar] [CrossRef] [Green Version]
- Appenrodt, E.; Schwarzberg, H. Central vasopressin administration failed to influence anxiety behavior after pinealectomy in rats. Physiol. Behav. 2000, 68, 735–739. [Google Scholar] [CrossRef]
- Karakaş, A.; Coşkun, H.; Kaya, A.; Kücük, A.; Gündüz, B. The effects of theintraamygdalar melatonin injections on the anxiety like behavior and thespatial memory performance in male wistar rats. Behav. Brain Res. 2011, 222, 141–150. [Google Scholar] [CrossRef]
- Zahra, E.; Siham, O.; Abdelhalim, M.; Aboubakr, E.; Ali, O. Pinealectomy and exogenous melatonin regulate anxiety-like and depressive-like behaviors in male and female Wistar rats. Neurosci. Med. 2012, 3, 394–403. [Google Scholar] [CrossRef] [Green Version]
- Tchekalarova, J.; Atanasova, D.; Nenchovska, Z.; Atanasova, M.; Kortenska, L.; Gesheva, R.; Lazarov, N. Agomelatine protects against neuronal damage without preventing epileptogenesis in the kainate model of temporal lobe epilepsy. Neurobiol. Dis. 2017, 104, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Kaya, A.; Karakaş, A.; Coşkun, H. The effects of the time of the day and the pinealectomy on anxiety-like behaviour in male Wistar rats. Biol. Rhythm. 2011, 42, 367–383. [Google Scholar] [CrossRef]
- Moyanova, S.; De Fusco, A.; Santolini, I.; Celli, R.; Bucci, D.; Mastroiacovo, F.; Battaglia, G.; Nicoletti, F.; Tchekalarova, J. Abnormal Hippocampal Melatoninergic System: A Potential Link between Absence Epilepsy and Depression-Like Behavior in WAG/Rij Rats? Int. J. Mol. Sci. 2018, 19, 1973. [Google Scholar] [CrossRef] [Green Version]
- Medel-Matus, J.S.; Shin, D.; Sankar, R.; Mazarati, A. Inherent vulnerabilities in monoaminergic pathways predict the emergence of depressive impairments in an animal model of chronic epilepsy. Epilepsia 2017, 58, e116–e121. [Google Scholar] [CrossRef] [PubMed]
- Gaffori, O.; Van Ree, J.M. Serotonin and antidepressant drugs antagonize melatonin-induced behavioural changes after injection into the nucleus accumbens of rats. Neuropharmacology 1985, 24, 237–244. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Stoynova, T.; Ilieva, K.; Mitreva, R.; Atanasova, M. Agomelatine treatment corrects symptoms of depression and anxiety by restoring the disrupted melatonin circadian rhythms of rats exposed to chronic constant light. Pharmacol. Biochem. Behav. 2018, 171, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Tchekalarova, J.; Atanasova, M.; Ivanova, N.; Boyadjiev, N.; Mitreva, R.; Georgieva, K. Endurance training exerts time-dependent modulation on depressive responses and circadian rhythms of corticosterone and BDNF in the rats with pinealectomy. Brain Res. Bull. 2020, 162, 40–48. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Kortenska, L.; Ivanova, N.; Atanasova, M.; Marinov, P. Agomelatine treatment corrects impaired sleep-wake cycle and sleep architecture and increases MT1 receptor as well as BDNF expression in the hippocampus during the subjective light phase of rats exposed to chronic constant light. Psychopharmacology 2020, 237, 503–518. [Google Scholar] [CrossRef]
- Kregel, K.C.; Zhang, H.J. An integrated view of oxidative stress in aging: Basic mechanisms, functional effects, and pathological considerations. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R18–R36. [Google Scholar] [CrossRef]
- Fukui, K.; Omoi, N.O.; Hayasaka, T.; Shinnkai, T.; Suzuki, S.; Abe, K.; Urano, S. Cognitive impairment of rats caused by oxidative stress and aging, and its prevention by vitamin E. Ann. N. Y. Acad Sci. 2002, 959, 275–284. [Google Scholar] [CrossRef]
- Tchekalarova, J.; Stoyanova, T.; Nenchovska, Z.; Ivanova, N.; Atanasova, D.; Atanasova, M.; Georgieva, K. Effect of endurance training on diurnal rhythms of superoxide dismutase activity, glutathione and lipid peroxidation in plasma of pinealectomized rats. Neurosci. Lett. 2020, 716, 134637. [Google Scholar] [CrossRef]
- Reiter, R.J. Oxidative processes and antioxidative defense mechanisms in the aging brain. FASEB J. 1995, 9, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Antolín, I.; Rodríguez, C.; Saínz, R.M.; Mayo, J.C.; Uría, H.; Kotler, M.L.; Rodríguez-Colunga, M.J.; Tolivia, D.; Menéndez-Peláez, A. Neurohormone melatonin prevents cell damage: Effect on gene expression for antioxidant enzymes. FASEB J. 1996, 10, 882–890. [Google Scholar] [CrossRef] [PubMed]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tchekalarova, J.; Nenchovska, Z.; Kortenska, L.; Uzunova, V.; Georgieva, I.; Tzoneva, R. Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats. Int. J. Mol. Sci. 2022, 23, 2809. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052809
Tchekalarova J, Nenchovska Z, Kortenska L, Uzunova V, Georgieva I, Tzoneva R. Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats. International Journal of Molecular Sciences. 2022; 23(5):2809. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052809
Chicago/Turabian StyleTchekalarova, Jana, Zlatina Nenchovska, Lidia Kortenska, Veselina Uzunova, Irina Georgieva, and Rumiana Tzoneva. 2022. "Impact of Melatonin Deficit on Emotional Status and Oxidative Stress-Induced Changes in Sphingomyelin and Cholesterol Level in Young Adult, Mature, and Aged Rats" International Journal of Molecular Sciences 23, no. 5: 2809. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23052809