
The Molecular Bases of the Interaction between a Saponin from the Roots of Gypsophila paniculata L. and Model Lipid Membranes

 , ,
, ,
Abstract
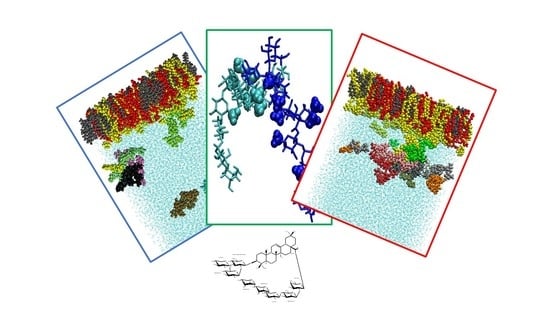
:
1. Introduction
2. Results and Discussion
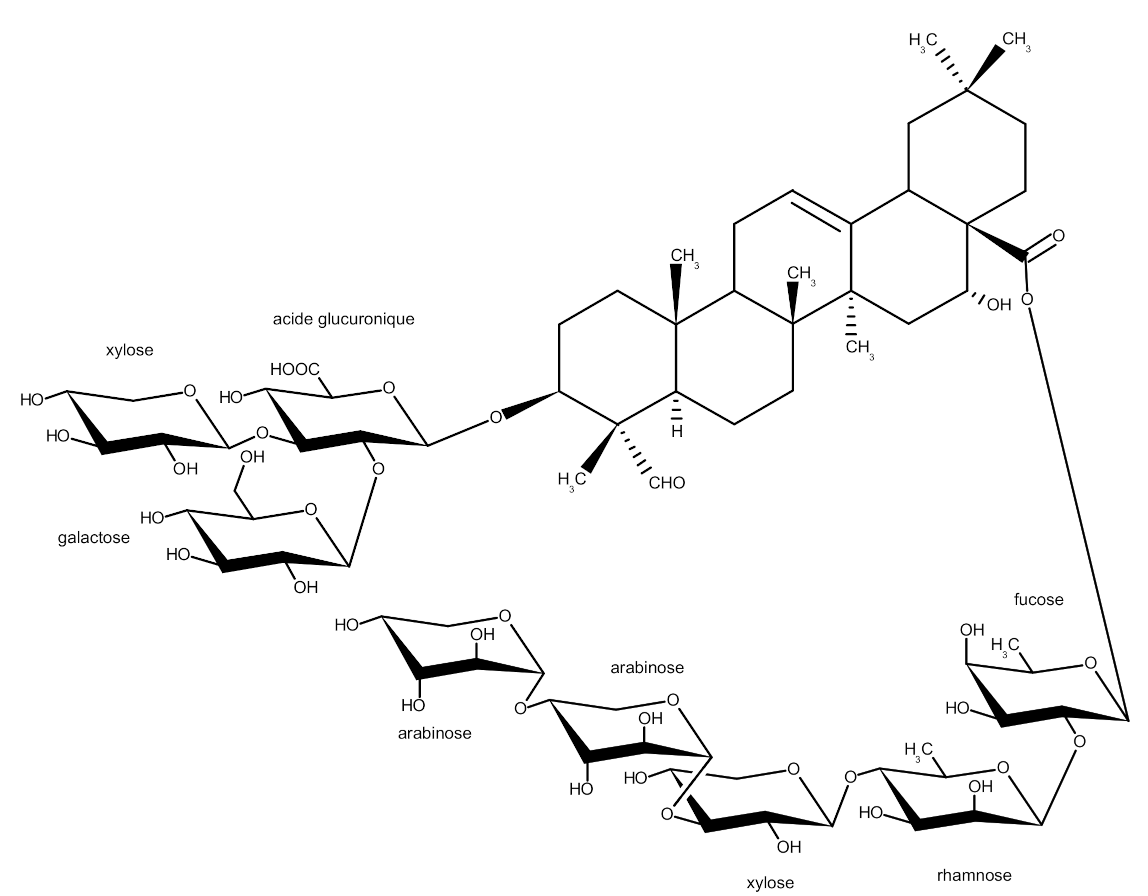
2.1. NMR Analysis
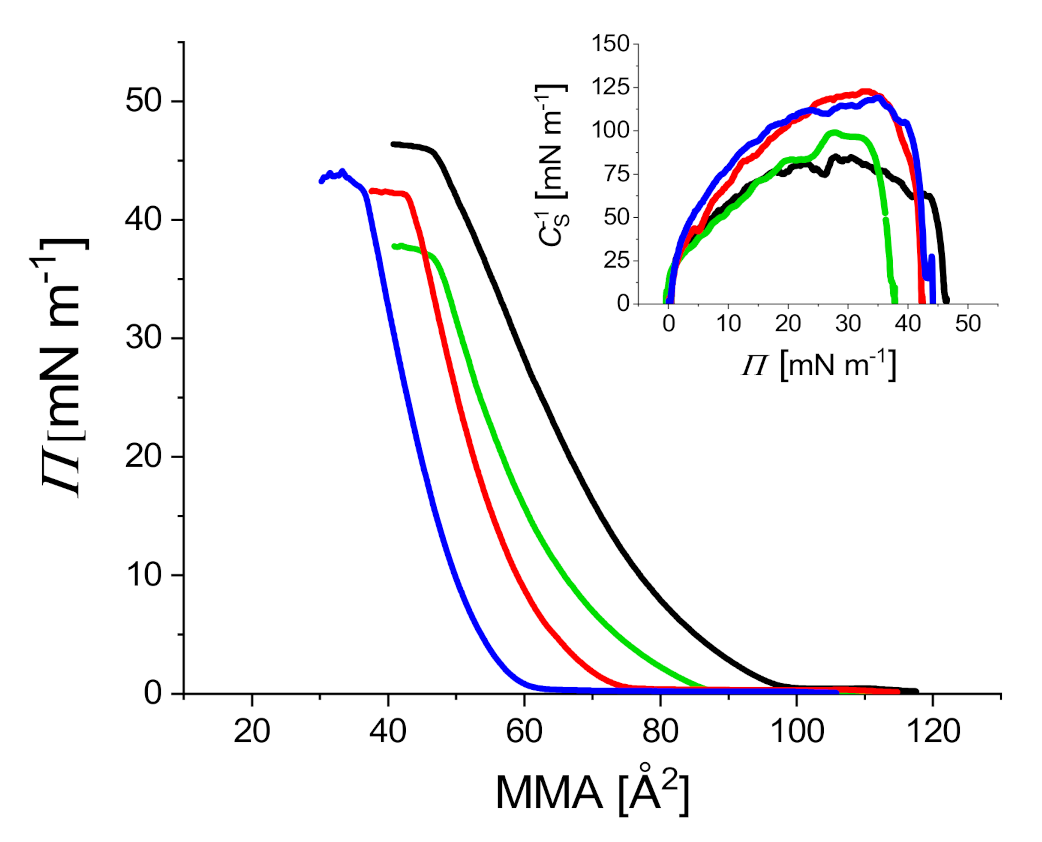
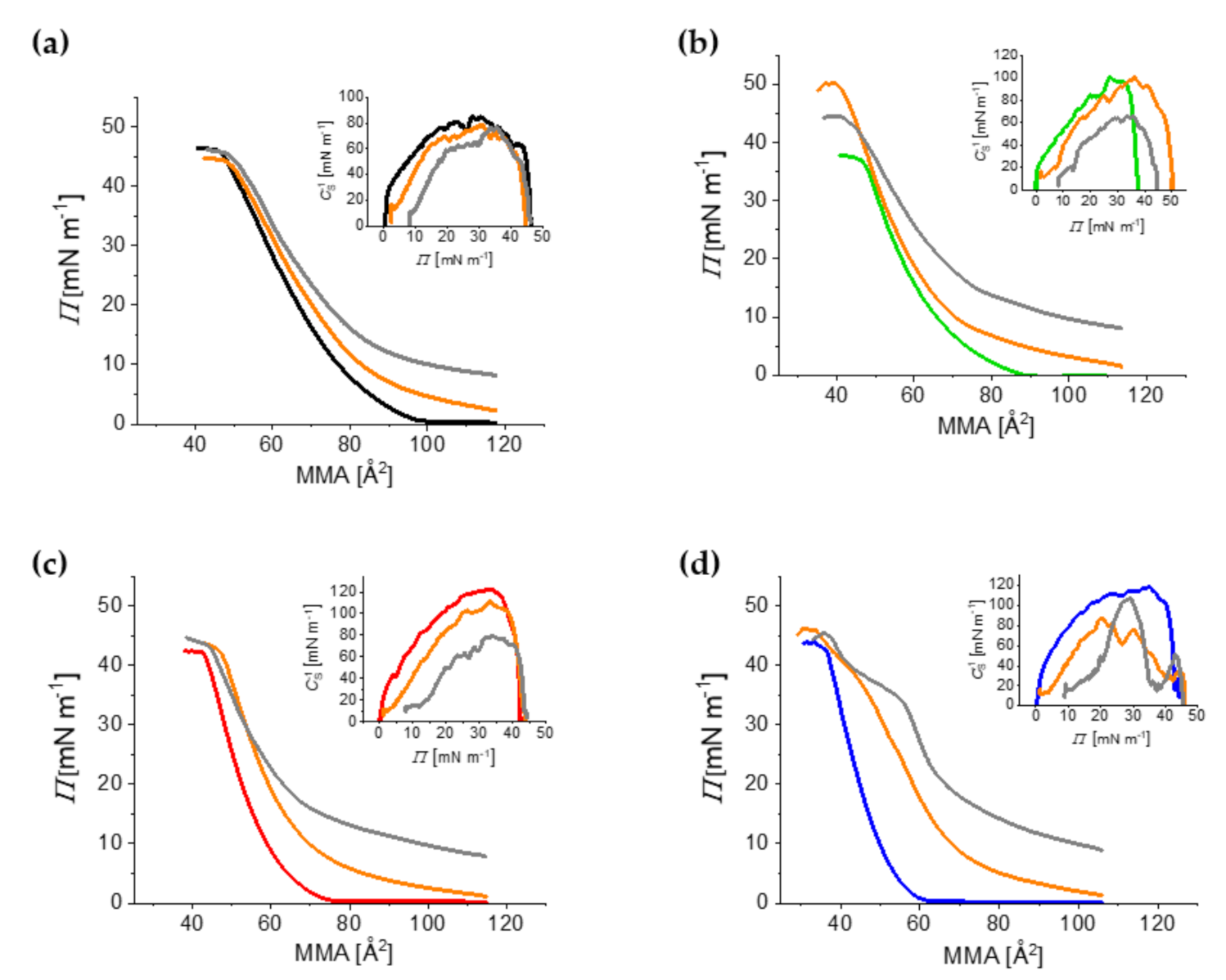
2.2. Compression Isotherms
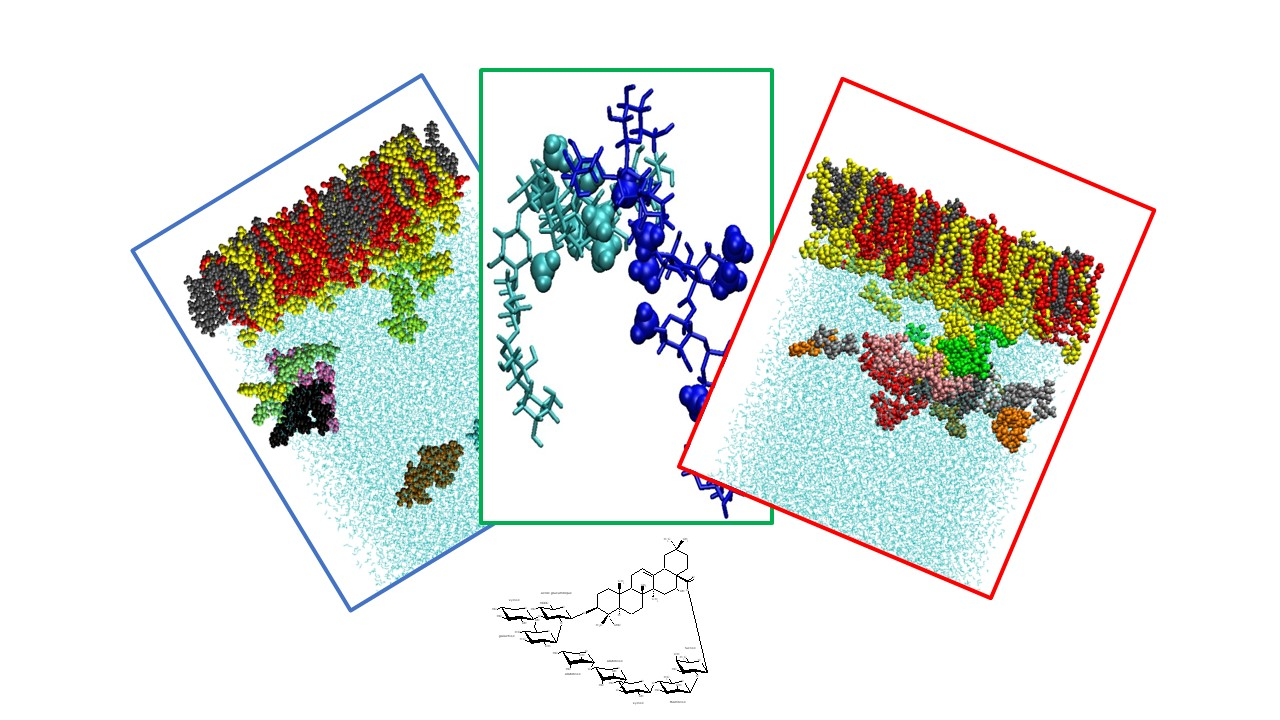
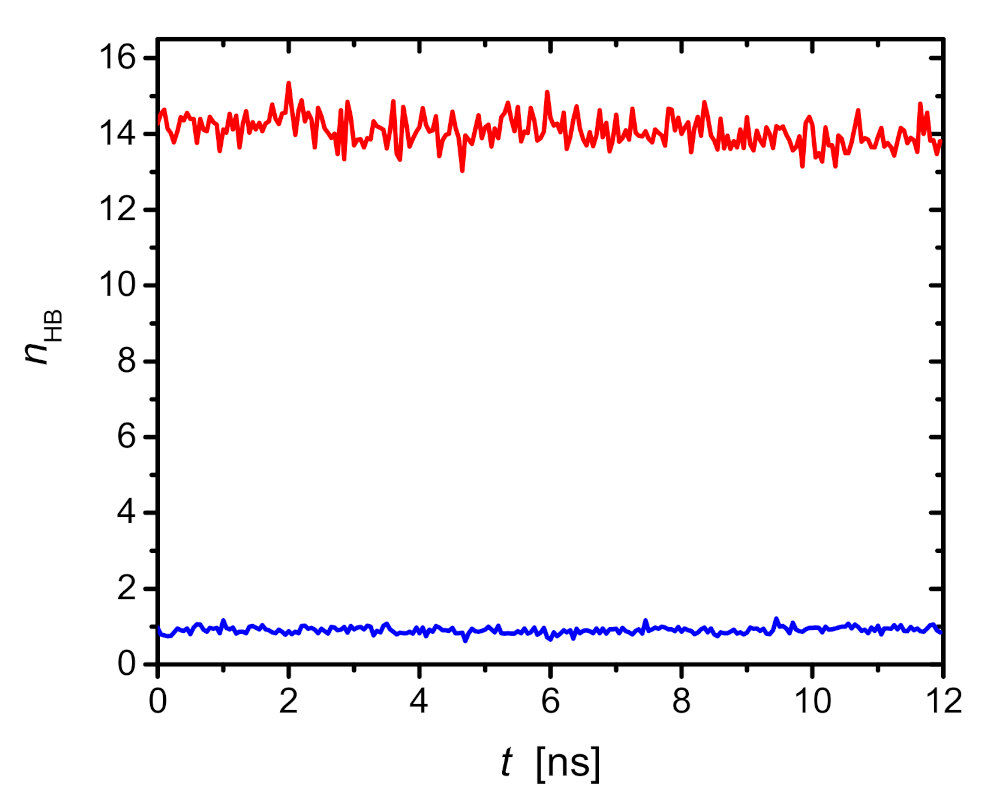
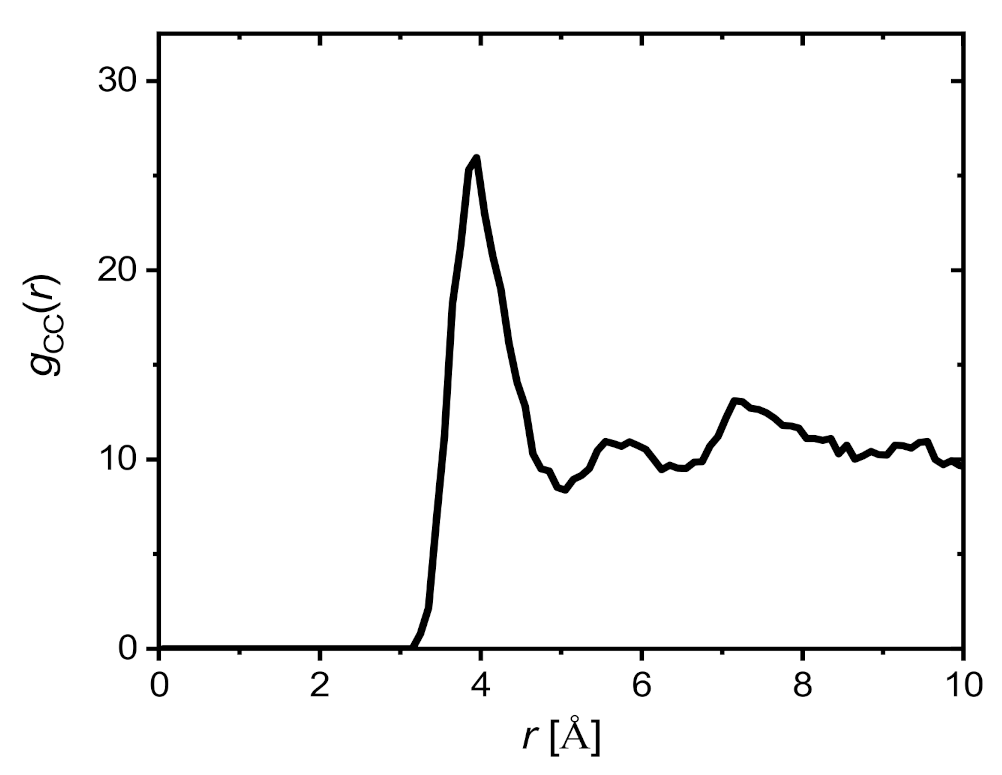
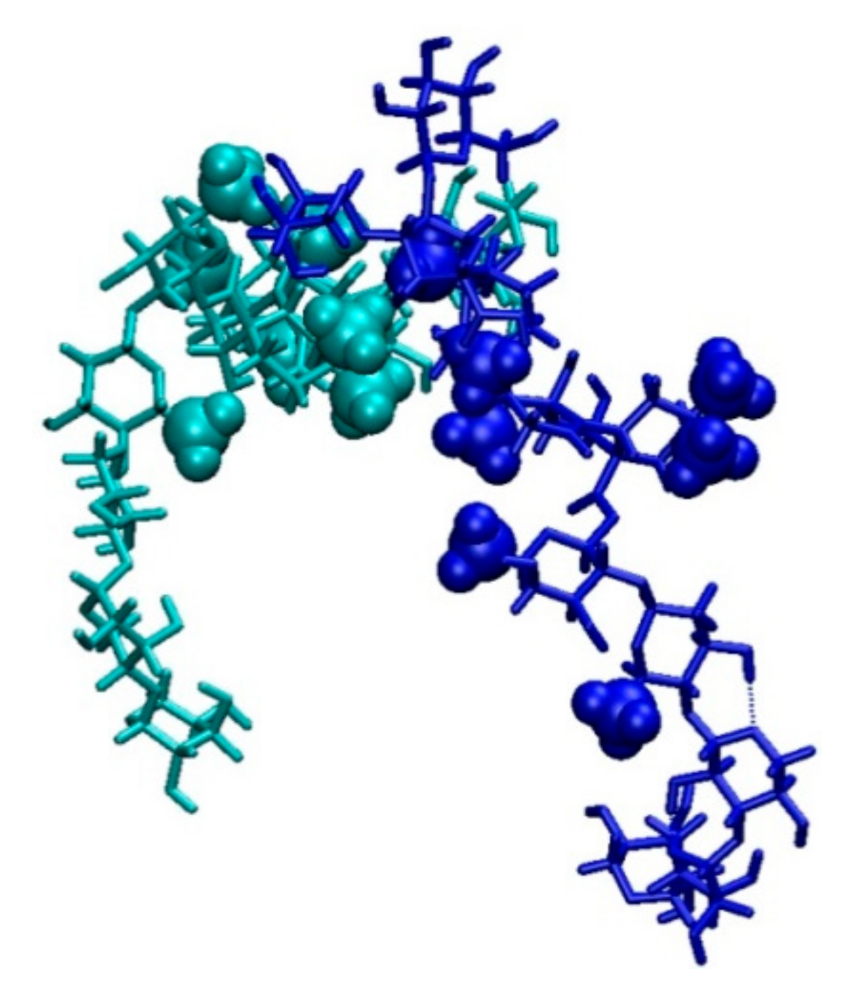
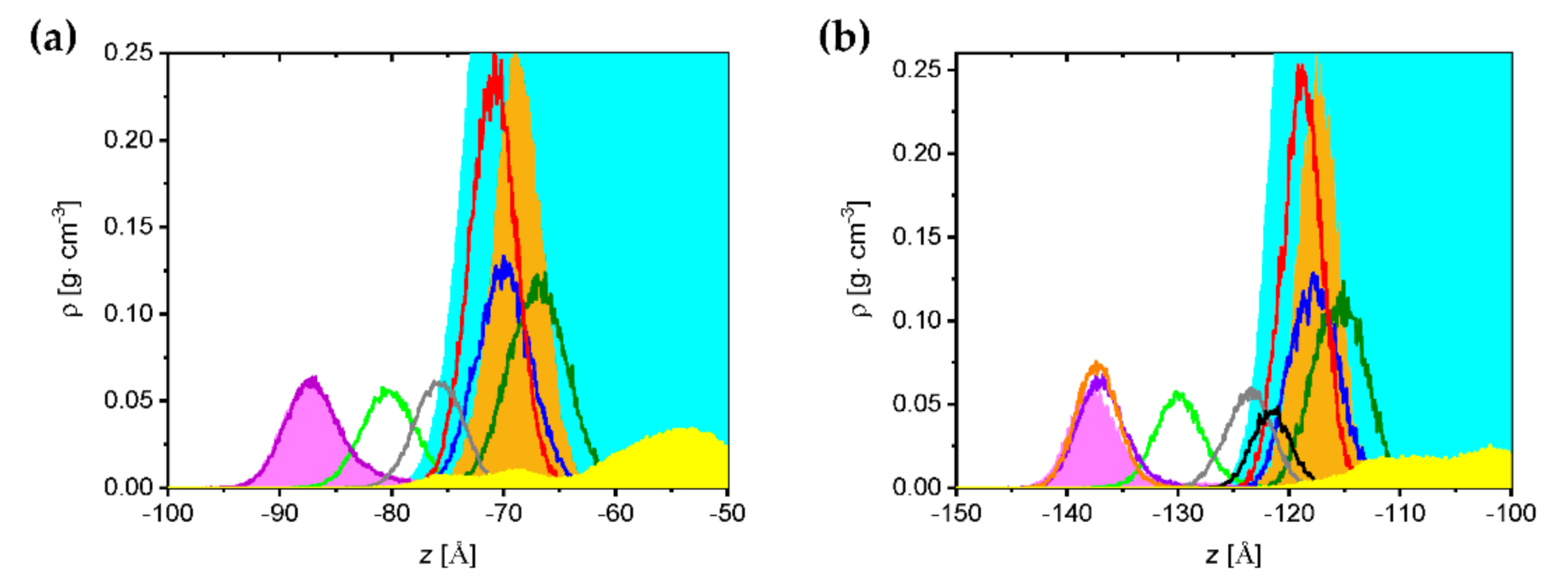
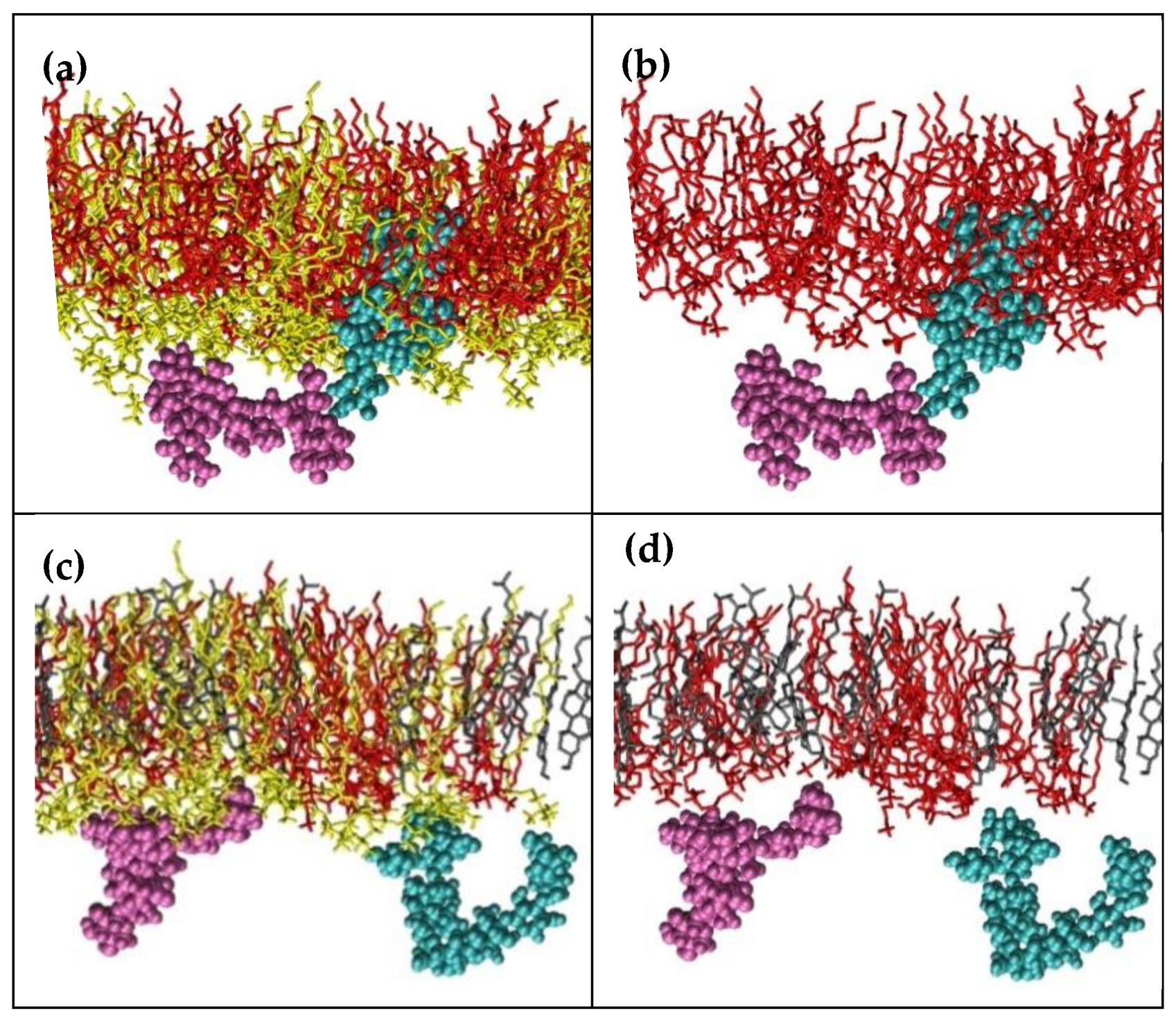
2.3. Molecular Dynamics Simulations
2.3.1. Saponin Molecules in a Box of Water
2.3.2. Saponin Molecules at the Air–Water Interface
3. Materials and Methods
3.1. Materials
3.2. NMR Experiments
3.3. Surface Pressure–Area Isotherms
3.4. Molecular Dynamics Simulations
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Thakur, M.; Weng, A.; Bachran, D.; Riese, S.B.; Boettger, S.; Melzig, M.F.; Fuchs, H. Electrophoretic isolation of saponin fractions from Saponinum album and their evaluation in synergistically enhancing the receptor-specific cytotoxicity of targeted toxins. Electrophoresis 2011, 32, 3085–3089. [Google Scholar] [CrossRef] [PubMed]
- Bachran, C.; Bachran, S.; Sutherland, M.; Bachran, D.; Fuchs, H. Saponins in tumor therapy. Mini-Rev. Med. Chem. 2008, 8, 575–584. [Google Scholar] [CrossRef]
- Thakur, M.; Melzig, M.F.; Fuchs, H.; Weng, A. Chemistry and pharmacology of saponins: Special focus on cytotoxic properties. Bot. Targets Ther. 2011, 1, 19. [Google Scholar]
- Skene, C.D.; Sutton, P. Saponin-adjuvanted particulate vaccines for clinical use. Methods 2006, 40, 53–59. [Google Scholar] [CrossRef]
- Sjolander, A.; Cox, J.C. Uptake and adjuvant activity of orally delivered saponin and ISCOM vaccines. Adv. Drug Deliv. Rev. 1998, 34, 321–338. [Google Scholar] [CrossRef]
- Hostettmann, K.; Marton, A. Saponins; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Hebestreit, P.; Weng, A.; Bachran, C.; Fuchs, H.; Melzig, M.F. Enhancement of cytotoxicity of lectins by Saponinum album. Toxicon 2006, 47, 330–335. [Google Scholar] [CrossRef]
- Bachran, C.; Duerkop, H.; Sutherland, M.; Bachran, D.; Mueller, C.; Weng, A.; Melzig, M.F.; Fuchs, H. Inhibition of Tumor Growth by Targeted Toxins in Mice is Dramatically Improved by Saponinum Album in a Synergistic Way. J. Immunother. 2009, 32, 713–725. [Google Scholar] [CrossRef] [PubMed]
- Weng, A.; Thakur, M.; Beceren-Braun, F.; Bachran, D.; Bachran, C.; Riese, S.B.; Jenett-Siems, K.; Gilabert-Oriol, R.; Melzig, M.F.; Fuchs, H. The toxin component of targeted anti-tumor toxins determines their efficacy increase by saponins. Mol. Oncol. 2012, 6, 323–332. [Google Scholar] [CrossRef] [Green Version]
- Boettger, S.; Westhof, E.; Siems, K.; Melzig, M.F. Structure-activity relationships of saponins enhancing the cytotoxicity of ribosome-inactivating proteins type I (RIP-I). Toxicon 2013, 73, 144–150. [Google Scholar] [CrossRef]
- Henry, M.; Rochd, M.; Bennini, B. Biosynthesis and accumulation of saponins in Gyposphila paniculata. Phytochemistry 1991, 30, 1819. [Google Scholar] [CrossRef]
- Frechet, D.; Christ, B.; Du Sorbier, B.M.; Fischer, H.; Vuilhorgne, M. Four triterpenoid saponins from dried roots of Gypsophila species. Phytochemistry 1991, 30, 927. [Google Scholar] [CrossRef]
- Kim, Y.C.; Higuchi, R.; Komori, T. Application of hydrothermolysis to the studies of the constituents of the Merck Saponin. Liebigs Ann. Chem. 1992, 1992, 941–946. [Google Scholar] [CrossRef]
- Delay, C.; Gavin, J.A.; Aumelas, A.; Bonnet, P.A.; Roumestand, C. Isolation and structure elucidation of a highly hemolytic saponin from the Merck saponin extract using high-field gradient-enhanced NMR techniques. Carbohydr. Res. 1997, 302, 67–78. [Google Scholar] [CrossRef]
- Yao, S.; Ma, L.; Luo, J.-G.; Wang, J.-S.; Kong, L.-Y. New Triterpenoid Saponins from the Roots of Gypsophila paniculata L. Helv. Chim. Acta 2010, 93, 361–374. [Google Scholar] [CrossRef]
- Weng, A.; Bachran, D.; Goerick, C.; Bachran, C.; Fuchs, H.; Melzig, M.F. A simple method for isolation of Gypsophila saponins for the combined application of targeted toxins and saponins in tumor therapy. Planta Med. 2009, 75, 1421–1422. [Google Scholar] [CrossRef] [Green Version]
- Weng, A.; Jenett-Siems, K.; Schmieder, P.; Bachran, D.; Bachran, C.; Goerick, C.; Thakur, M.; Fuchs, H.; Melzig, M.F. A convenient method for saponin isolation in tumour therapy. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 713–718. [Google Scholar]
- Gilabert-Oriol, R.; Weng, A.; von Mallinckrodt, B.; Stoeshel, A.; Nissi, L.; Melzig, M.F.; Fuchs, H.; Thakur, M. Electrophoretic mobility as a tool to separate immune adjuvant saponins from Quillaja saponaria Molina. Int. J. Pharm. 2015, 487, 39–48. [Google Scholar] [CrossRef]
- Voutquenne-Nazabadioko, L.; Gevrenova, R.; Borie, N.; Harakat, D.; Sayagh, C.; Weng, A.; Thakur, M.; Zaharieva, M.; Henry, M. Triterpenoid saponins from the roots of Gypsophila trichotoma Wender. Phytochemistry 2013, 90, 114–127. [Google Scholar] [CrossRef]
- Gevrenova, R.; Joubert, O.; Mandova, T.; Zaiou, M.; Chapleur, Y.; Henry, M. Cytotoxic effects of four Caryophyllaceae species extracts on macrophage cell lines. Pharm. Biol. 2014, 52, 919–925. [Google Scholar] [CrossRef] [Green Version]
- Gevrenova, R.; Doytchinova, I.; Kolodziej, B.; Henry, M. In-depth characterization of the GOTCAB saponins in seven cultivated Gypsophila L. species (Caryophyllaceae) by liquid chromatography coupled with quadrupole-Orbitrap mass spectrometer. Biochem. Syst. Ecol. 2019, 83, 91–102. [Google Scholar] [CrossRef]
- Korchowiec, B.; Korchowiec, J.; Gorczyca, M.; Regnouf de Vains, J.-B.; Rogalska, E. Molecular Organization of Nalidixate Conjugated Calixarenes in Bacterial Model Membranes Probed by Molecular Dynamics Simulation and Langmuir Monolayer Studies. J. Phys. Chem. B 2015, 119, 2990–3000. [Google Scholar] [CrossRef] [PubMed]
- Korchowiec, B.M.; Baba, T.; Minamikawa, H.; Hato, M. Forces that Control pH-Dependent Aggregation of Nonionic Glycolipid Vesicles. Langmuir 2001, 17, 1853–1859. [Google Scholar] [CrossRef]
- Stefaniu, C.; Brezesinski, G.; Moehwald, H. Langmuir monolayers as models to study processes at membrane surfaces. Adv. Colloid Interface Sci. 2014, 208, 197–213. [Google Scholar] [CrossRef] [PubMed]
- Hilderson, H.J.; Ralston, G.B. Physicochemical Methods in the Study of Biomembranes; Springer Science & Business Media: Berlin/Heidelberg, Germany, 1994; Volume 23, p. 509. [Google Scholar]
- Yeagle, P.L. The Structure of Biological Membranes, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2012; p. 386. [Google Scholar]
- Korchowiec, B.; Gorczyca, M.; Wojszko, K.; Janikowska, M.; Henry, M.; Rogalska, E. Impact of two different saponins on the organization of model lipid membranes. Biochim. Biophys. Acta Biomembr. 2015, 1848, 1963–1973. [Google Scholar] [CrossRef]
- Korchowiec, B.; Janikowska-Sagan, M.; Kwiecinska, K.; Stachowicz-Kusnierz, A.; Korchowiec, J. The role of cholesterol in membrane activity of digitonin: Experimental and theoretical model studies. J. Mol. Liq. 2021, 323, 114598. [Google Scholar] [CrossRef]
- Lorent, J.H.; Leonard, C.; Abouzi, M.; Akabi, F.; Quetin-Leclercq, J.; Mingeot-Leclercq, M.-P. α-Hederin Induces Apoptosis, Membrane Permeabilization and Morphologic Changes in Two Cancer Cell Lines Through a Cholesterol-Dependent Mechanism. Planta Med. 2016, 82, 1532–1539. [Google Scholar] [CrossRef] [Green Version]
- Verstraeten, S.L.; Deleu, M.; Janikowska-Sagan, M.; Claereboudt, E.J.S.; Lins, L.; Tyteca, D.; Mingeot-Leclercq, M.-P. The activity of the saponin ginsenoside Rh2 is enhanced by the interaction with membrane sphingomyelin but depressed by cholesterol. Sci. Rep. 2019, 9, 7285. [Google Scholar] [CrossRef]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef]
- Simons, K.; Sampaio, J.L. Membrane organization and lipid rafts. Cold Spring Harb. Perspect. Biol. 2011, 3, 17. [Google Scholar] [CrossRef]
- Risselada, H.J.; Marrink, S.J. The molecular face of lipid rafts in model membranes. Proc. Natl. Acad. Sci. USA 2008, 105, 17367–17372. [Google Scholar] [CrossRef] [Green Version]
- Korchowiec, B.; Korchowiec, J.; Orlof-Naturalna, M.; Regnouf de Vains, J.-B.; Rogalska, E. Two antibacterial nalidixate calixarene derivatives in cholesterol monolayers: Molecular dynamics and physicochemical effects. Colloids Surf. B 2016, 145, 777–784. [Google Scholar] [CrossRef]
- Zhao, L.; Feng, S.-S. Effects of cholesterol component on molecular interactions between paclitaxel and phospholipid within the lipid monolayer at the air-water interface. J. Colloid Interface Sci. 2006, 300, 314–326. [Google Scholar] [CrossRef]
- Martinez-Seara, H.; Rog, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M.; Reigada, R. Interplay of unsaturated phospholipids and cholesterol in membranes: Effect of the double-bond position. Biophys. J. 2008, 95, 3295–3305. [Google Scholar] [CrossRef] [Green Version]
- Rog, T.; Pasenkiewicz-Gierula, M.; Vattulainen, I.; Karttunen, M. What happens if cholesterol is made smoother: Importance of methyl substituents in cholesterol ring structure on phosphatidylcholine-sterol interaction. Biophys. J. 2007, 92, 3346–3357. [Google Scholar] [CrossRef] [Green Version]
- Subczynski, W.K.; Pasenkiewicz-Gierula, M.; Widomska, J.; Mainali, L.; Raguz, M. High Cholesterol/Low Cholesterol: Effects in Biological Membranes Review. Cell Biochem. Biophys. 2017, 75, 369–385. [Google Scholar] [CrossRef]
- Gorczyca, M.; Korchowiec, B.; Korchowiec, J.; Trojan, S.; Rubio-Magnieto, J.; Luis, S.V.; Rogalska, E. A Study of the Interaction between a Family of Gemini Amphiphilic Pseudopeptides and Model Monomolecular Film Membranes Formed with a Cardiolipin. J. Phys. Chem. B 2015, 119, 6668–6679. [Google Scholar] [CrossRef]
- Korchowiec, B.; Gorczyca, M.; Rogalska, E.; Regnouf-de-Vains, J.-B.; Mourer, M.; Korchowiec, J. The selective interactions of cationic tetra-p-guanidinoethylcalix[4] arene with lipid membranes: Theoretical and experimental model studies. Soft Matter 2016, 12, 181–190. [Google Scholar] [CrossRef]
- Korchowiec, J.; Korchowiec, B.; Priebe, W.; Rogalska, E. DFT Study on the Selectivity of Complexation of Metal Cations with a Dioxadithia Crown Ether Ligand. J. Phys. Chem. A 2008, 112, 13633–13640. [Google Scholar] [CrossRef]
- Stachowicz-Kusnierz, A.; Trojan, S.; Cwiklik, L.; Korchowiec, B.; Korchowiec, J. Modeling Lung Surfactant Interactions with Benzo[a]pyrene. Chem. Eur. J. 2017, 23, 5307–5316. [Google Scholar] [CrossRef]
- Korchowiec, B.; Orlof-Naturalna, M.; Korchowiec, J.; de Vains, J.B.R.; Mourer, M.; Rogalska, E. The hydrophobic core effect in model bacterial membranes upon interaction with tetra-p-guanidinoethylcalix 4 arene. J. Mol. Liq. 2021, 343, 117636. [Google Scholar] [CrossRef]
- Slotte, J.P. Sphingomyelin-cholesterol interactions in biological and model membranes. Chem. Phys. Lipids 1999, 102, 13–27. [Google Scholar] [CrossRef]
- Davies, J.T.; Rideal, E.K. Interfacial Phenomena, 2nd ed.; Academic Press: Cambridge, MA, USA, 1963; p. 480. [Google Scholar]
- Freudenthal, O.; Quiles, F.; Francius, G.; Wojszko, K.; Gorczyca, M.; Korchowiec, B.; Rogalska, E. Nanoscale investigation of the interaction of colistin with model phospholipid membranes by Langmuir technique, and combined infrared and force spectroscopies. Biochim. Biophys. Acta Biomembr. 2016, 1858, 2592–2602. [Google Scholar] [CrossRef] [PubMed]
- Gravier, J.; Korchowiec, B.; Schneider, R.; Rogalska, E. Interaction of amphiphilic chlorin-based photosensitizers with 1,2-dipalmitoyl-sn-glycero-3-phosphocholine monolayers. Chem. Phys. Lipids 2009, 158, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Vanommeslaeghe, K.; Hatcher, E.; Acharya, C.; Kundu, S.; Zhong, S.; Shim, J.; Darian, E.; Guvench, O.; Lopes, P.; Vorobyov, I.; et al. CHARMM general force field: A force field for drug-like molecules compatible with the CHARMM all-atom additive biological force fields. J. Comput. Chem. 2010, 31, 671–690. [Google Scholar] [CrossRef] [Green Version]
- Guvench, O.; Hatcher, E.; Venable, R.M.; Pastor, R.W.; MacKerell, A.D., Jr. CHARMM Additive All-Atom Force Field for Glycosidic Linkages between Hexopyranoses. J. Chem. Theory Comput. 2009, 5, 2353–2370. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VDM: Visual molecular dynamics. J. Mol. Graph. 1996, 14, 27–28. [Google Scholar] [CrossRef]
- Mayne, C.G.; Saam, J.; Schulten, K.; Tajkhorshid, E.; Gumbart, J.C. Rapid parameterization of small molecules using the Force Field Toolkit. J. Comput. Chem. 2013, 34, 2757–2770. [Google Scholar] [CrossRef] [Green Version]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Darden, T.; York, D.; Pedersen, L. Particle mesh Ewald: An N⋅log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Jorgensen, W.L.; Chandrasekhar, J.; Madura, J.D.; Impey, R.W.; Klein, M.L. Comparison of Simple Potential Functions for Simulating Liquid Water. J. Chem. Phys. 1983, 79, 926–935. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Atom Number | 1H (ppm) (J/Hz) | 13C (ppm) |
|---|---|---|
| 1 | 1.39; 0.78 | 37.4 |
| 2 | 1.83; 2.13 | 23.8 |
| 3 | 3.90 | 84.4 |
| 4 | – | 54.8 |
| 5 | 1.22 | 43.7 |
| 6 | 0.85; 1.22 | 19.7 |
| 7 | 1.31; 1.46 | 31.7 |
| 8 | – | 39.2 |
| 9 | 1.60 | 46.0 |
| 10 | – | 35.3 |
| 11 | 1.60; 1.74 | 22.7 |
| 12 | 5.38 | 121.7 |
| 13 | – | 143.2 |
| 14 | – | 41.0 |
| 15 | 1.72; 1.89 | 35.2 |
| 16 | 4.85 | 72.9 |
| 17 | – | 48.3 |
| 18 | 3.12 | 40.9 |
| 19 | 1.07; 2.41 | 46.3 |
| 20 | – | 29.7 |
| 21 | 2.02; 1.12 | 34.7 |
| 22 | 2.13; 1.90 | 30.3 |
| 23 | 9.55 | 210.5 |
| 24 | 1.20 | 9.5 |
| 25 | 0.72 | 14.8 |
| 26 | 0.83 | 16.5 |
| 27 | 1.50 | 26.2 |
| 28 | – | 175.8 |
| 29 | 0.78 | 32.1 |
| 30 | 0.78 | 23.6 |
| Atom Number | 1H (ppm) (J/Hz) | 13C (ppm) |
|---|---|---|
| Fucose (Fuc) | ||
| 1 | 5.59 (d, 8.4) | 93.7 |
| 2 | 4.25 (t, 8.8) | 73.4 |
| 3 | 3.90 (m) | 74.6 |
| 4 | 3.90 (m) | 71.5 |
| 5 | 3.75 (m) | 71.5 |
| 6 | 1.31 (d, 6.3) | 15.5 |
| Arabinose (Ara) | ||
| 1 | 4.89 (d, 7.7) | 103.6 |
| 2 | 4.13 (t, 8.3) | 71.5 |
| 3 | 4.01 (m) | 72.4 |
| 4 | 4.17 (m) | 77.8 |
| 5 | 4.33 (dd, 12.7, 2.3) 3.73 (m) | 65.3 |
| terminal Arabinose (Ara’) | ||
| 1 | 4.70 (d, 7.7) | 105.1 |
| 2 | 4.07 (t, 7.6) | 71.1 |
| 3 | 3.90 (m) | 72.4 |
| 4 | 4.09 (m) | 68.0 |
| 5 | 4.05 (dd, 9.6, -) 3.66 (dd, 9.6, -) | 65.9 |
| terminal Xylose (Xyl) | ||
| 1 | 5.04 (d, 7.7) | 103.0 |
| 2 | 3.67 (t, 8.2) | 73.7 |
| 3 | 3.86 (m) | 76.2 |
| 4 | 3.89 (m) | 69.4 |
| 5 | 4.17 (dd, 10.1, 5.0) 3.55 (t, 10.1) | 65.0 |
| Xylose (Xyl’) | ||
| 1 | 4.85 (d, 7.7) | 105.1 |
| 2 | 3.72 (t, 8.4) | 73.9 |
| 3 | 3.87 (m) | 84.6 |
| 4 | 3.81 (m) | 67.7 |
| 5 | 3.98 (dd, 11.1, 5.6) 3.38 (t, 11.1) | 65.2 |
| Rhamnose (Rha) | ||
| 1 | 5.82 (d, 1.1) | 100.1 |
| 2 | 4.46 (d, 3.3) | 70.3 |
| 3 | 4.26 (dd, 9.1, 3.3) | 70.8 |
| 4 | 3.98 (m) | 82.7 |
| 5 | 4.10 (m) | 67.6 |
| 6 | 1.49 (d, 6.2) | 17.3 |
| terminal Galactose (Gal) | ||
| 1 | 5.16 (d, 7.5) | 102.3 |
| 2 | 3.97 (dd, 7.5, 9.3) | 71.7 |
| 3 | 3.92 (m) | 73.2 |
| 4 | 4.18 (m) | 69.1 |
| 5 | 3.8 (m) | 75.4 |
| 6 | 4.16 (m) 4.02 (m) | 61.1 |
| Glucuronic acid (GlcA) | ||
| 1 | 4.66 (d, 7.11) | 102.4 |
| 2 | 4.10 (m) | 76.6 |
| 3 | 4.10 (m) | 84.2 |
| 4 | 4.00 (m) | 70.5 |
| 5 | 4.08 (m) | 76.8 |
| 6 | - | 174.7 |
| n = 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | |
|---|---|---|---|---|---|---|---|---|
| System I | 7.39 | 4.69 | 1.82 | 0.26 | 0.44 | 0.16 | 0.08 | 0.00 |
| System II | 9.58 | 2.04 | 1.52 | 0.32 | 0.47 | 0.15 | 0.58 | 0.03 |
| System III | 8.69 | 2.40 | 1.07 | 1.75 | 0.35 | 0.20 | 0.05 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korchowiec, B.; Korchowiec, J.; Kwiecińska, K.; Gevrenova, R.; Bouguet-Bonnet, S.; Deng, C.; Henry, M.; Rogalska, E. The Molecular Bases of the Interaction between a Saponin from the Roots of Gypsophila paniculata L. and Model Lipid Membranes. Int. J. Mol. Sci. 2022, 23, 3397. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063397
Korchowiec B, Korchowiec J, Kwiecińska K, Gevrenova R, Bouguet-Bonnet S, Deng C, Henry M, Rogalska E. The Molecular Bases of the Interaction between a Saponin from the Roots of Gypsophila paniculata L. and Model Lipid Membranes. International Journal of Molecular Sciences. 2022; 23(6):3397. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063397
Chicago/Turabian StyleKorchowiec, Beata, Jacek Korchowiec, Klaudia Kwiecińska, Reneta Gevrenova, Sabine Bouguet-Bonnet, Cheng Deng, Max Henry, and Ewa Rogalska. 2022. "The Molecular Bases of the Interaction between a Saponin from the Roots of Gypsophila paniculata L. and Model Lipid Membranes" International Journal of Molecular Sciences 23, no. 6: 3397. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23063397