
TRPV2: A Key Player in Myelination Disorders of the Central Nervous System

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
2.1. Opalin Interacts In Vitro with the Ankyrin-Repeat Domain (ARD) of TRPV2
2.2. TRPV2 and Opalin Display a High Degree of Colocalization in Mixed Glial Cultures Enriched in Oligodendrocytes
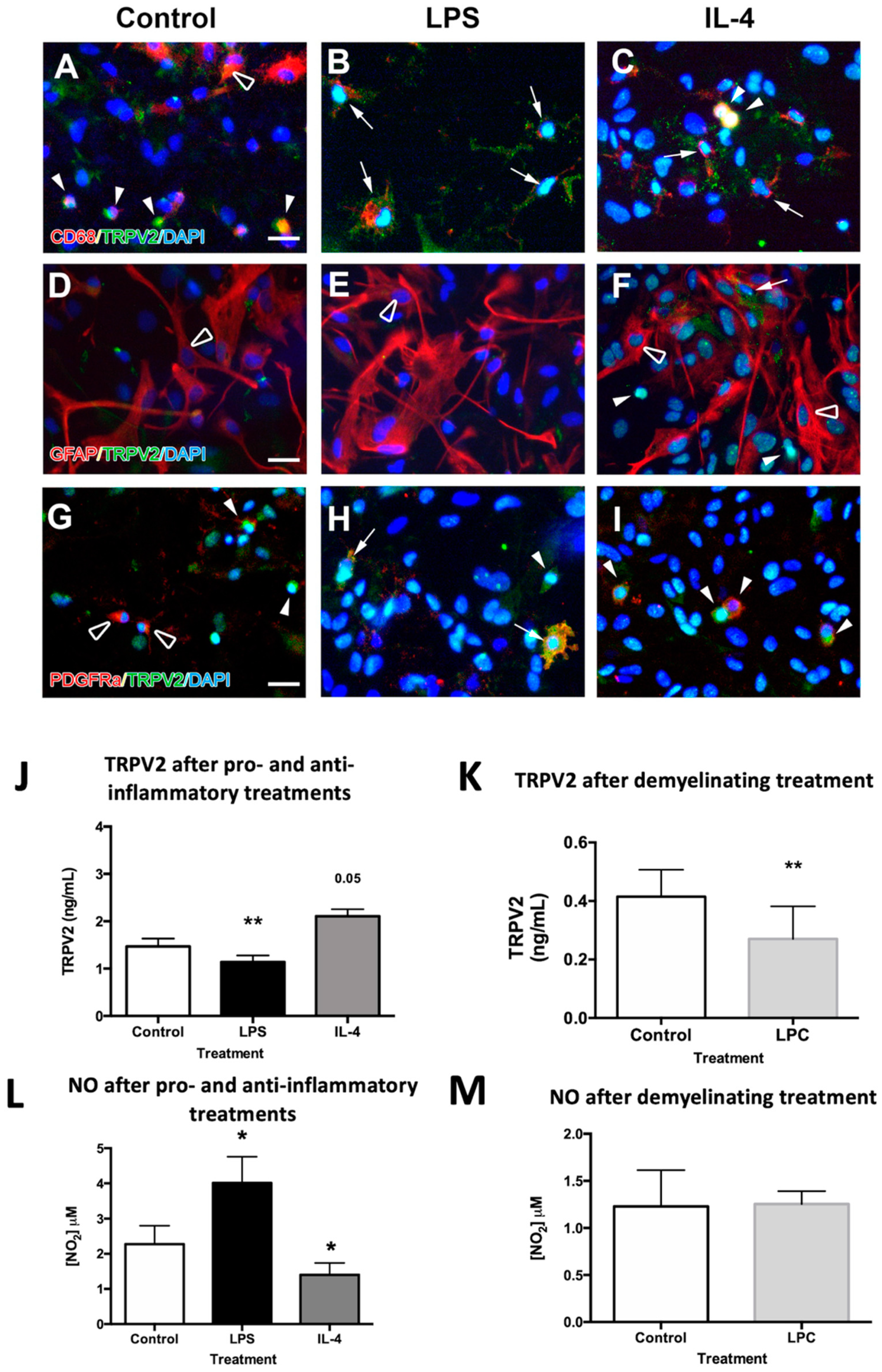
2.3. TRPV2 Expression in Microglia and Oligodendrocytes Is Regulated after Pro-Inflammatory, Anti-Inflammatory and Demyelinating Treatments
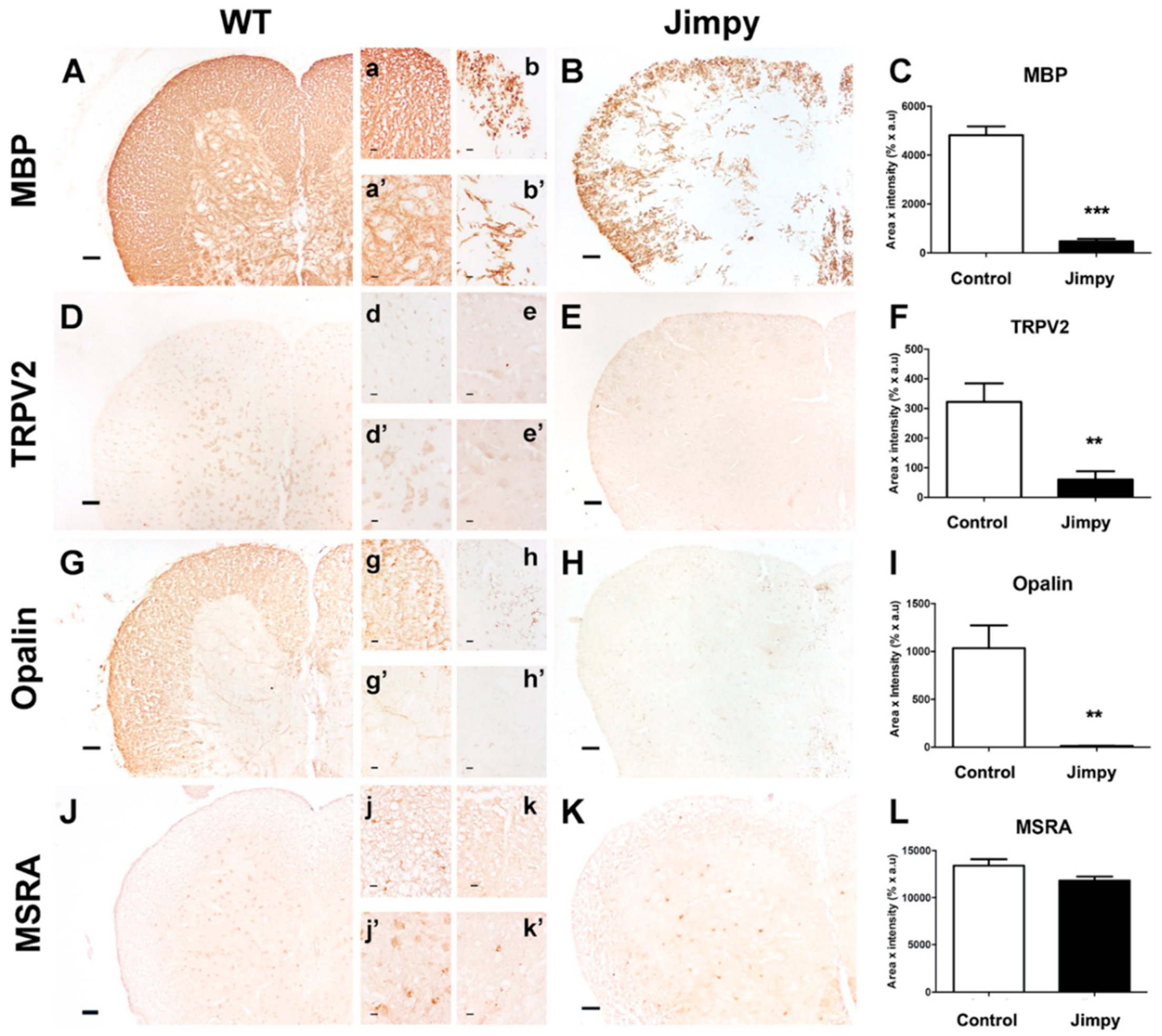
2.4. TRPV2 and Opalin Expression, but Not Methionine Sulfoxide Reductase A (MSRA) Levels, Are Importantly Reduced in the Hypomyelinating Jimpy Mutant Mice
2.5. TRPV2 and MSRA Expression Are Modulated in the Experimental Mouse Models of MS: The Cuprizone and the EAE Models
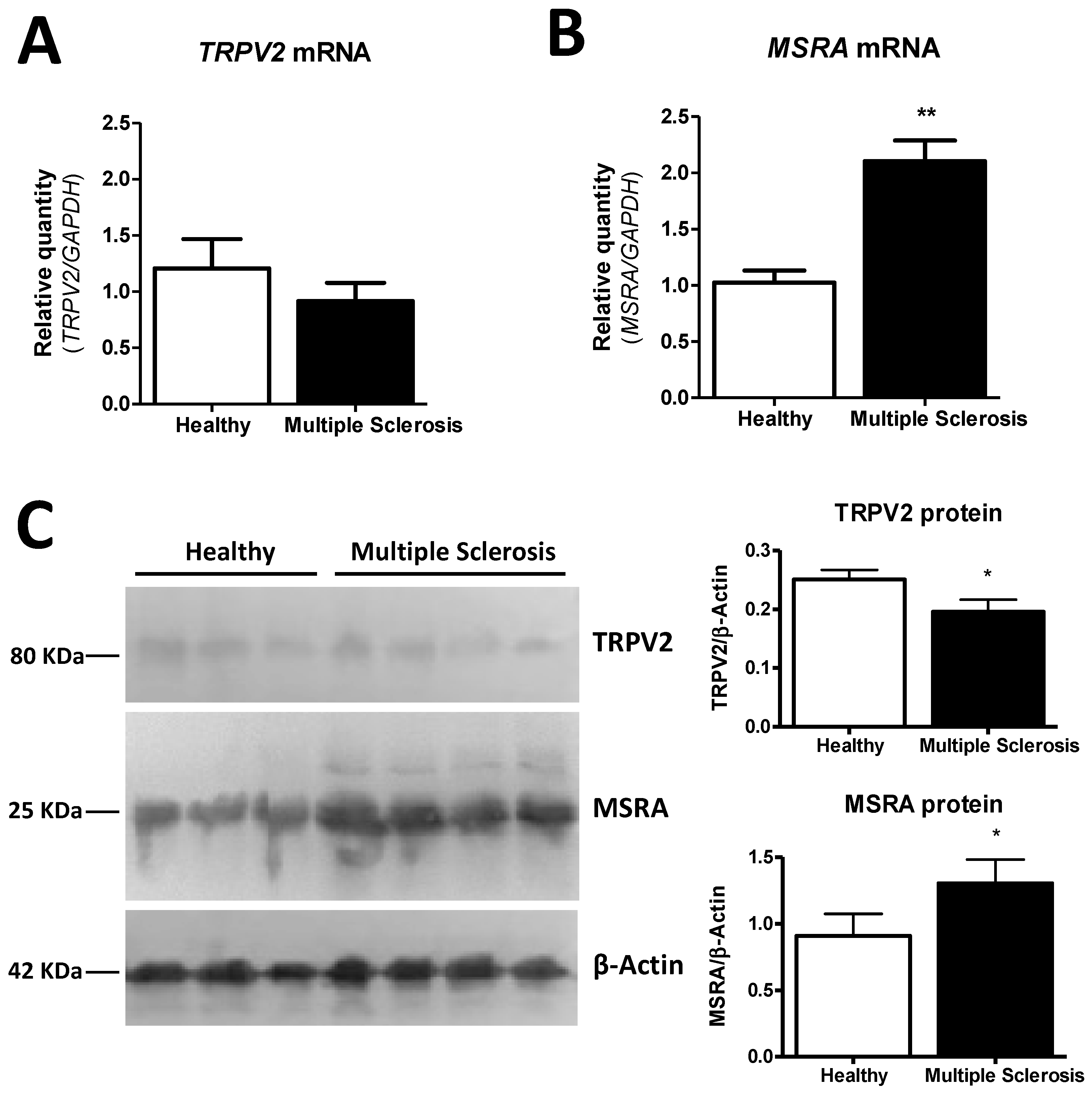
2.6. MSRA and TRPV2 Gene and Protein Expression Are Modified in Frontal Cortex Samples of MS Patients
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Cuprizone-Induced Demyelination and Remyelination in Mice
4.3. Experimental Autoimmune Encephalomyelitis in Mice
4.4. Human Samples
4.5. FL and ARD TRPV2 Fragment Purification
4.6. Slot Blot
4.7. Cell Cultures
4.8. Cell Culture Treatments
4.9. Nitrite Assay
4.10. Immunocytochemistry
4.11. Colocalization Studies of Opalin and TRPV2 in Cell Cultures
4.12. Quantification of TRPV2 in Glial Cell Cultures by Enzyme-Linked Immunosorbent Assay
4.13. Tissue Fixation and Processing for Immunohistochemistry
4.14. Immunohistochemistry
4.15. Quantification of Immunohistochemical Stainings
4.16. Double Immunohistochemistry
4.17. Isolation of Total Proteins
4.18. Quantification of TRPV2 and MSRA in EAE Mouse and Human Samples by Western Blot
4.19. RNA Extraction, cDNA Synthesis and Expression Analyses in Human Samples
4.20. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Perálvarez-Marín, A.; Doñate-Macian, P.; Gaudet, R. What do we know about the transient receptor potential vanilloid 2 (TRPV2) ion channel? FEBS J. 2013, 280, 5471–5487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fricke, T.C.; Echtermeyer, F.; Zielke, J.; de la Roche, J.; Filipovic, M.R.; Claverol, S.; Herzog, C.; Tominaga, M.; Pumroy, R.A.; Moiseenkova-Bell, V.Y.; et al. Oxidation of methionine residues activates the high-threshold heat-sensitive ion channel TRPV2. Proc. Natl. Acad. Sci. USA 2019, 116, 24359–24365. [Google Scholar] [CrossRef] [PubMed]
- Kojima, I.; Nagasawa, M. TRPV2. Handb. Exp. Pharmacol. 2014, 222, 247–272. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Rosen, T.A.; Tominaga, M.; Brake, A.J.; Julius, D. A capsaicin-receptor homologue with a high threshold for noxious heat. Nature 1999, 398, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Maksoud, M.J.E.; Tellios, V.; An, D.; Xiang, Y.; Lu, W. Nitric oxide upregulates microglia phagocytosis and increases transient receptor potential vanilloid type 2 channel expression on the plasma membrane. Glia 2019, 67, 2294–2311. [Google Scholar] [CrossRef]
- Shibasaki, K.; Ishizaki, Y.; Mandadi, S. Astrocytes express functional TRPV2 ion channels. Biochem. Biophys. Res. Commun. 2013, 441, 327–332. [Google Scholar] [CrossRef]
- Cahoy, J.D.; Emery, B.; Kaushal, A.; Foo, L.C.; Zamanian, J.L.; Christopherson, K.S.; Xing, Y.; Lubischer, J.L.; Krieg, P.A.; Krupenko, S.A.; et al. A Transcriptome Database for Astrocytes, Neurons, and Oligodendrocytes: A New Resource for Understanding Brain Development and Function. J. Neurosci. 2008, 28, 264–278. [Google Scholar] [CrossRef] [Green Version]
- Cohen, M.R.; Johnson, W.M.; Pilat, J.M.; Kiselar, J.; DeFrancesco-Lisowitz, A.; Zigmond, R.E.; Moiseenkova-Bell, V.Y. Nerve Growth Factor Regulates Transient Receptor Potential Vanilloid 2 via Extracellular Signal-Regulated Kinase Signaling to Enhance Neurite Outgrowth in Developing Neurons. Mol. Cell. Biol. 2015, 35, 4238–4252. [Google Scholar] [CrossRef] [Green Version]
- Shibasaki, K.; Murayama, N.; Ono, K.; Ishizaki, Y.; Tominaga, M. TRPV2 Enhances Axon Outgrowth through Its Activation by Membrane Stretch in Developing Sensory and Motor Neurons. J. Neurosci. 2010, 30, 4601–4612. [Google Scholar] [CrossRef] [Green Version]
- Link, T.M.; Park, U.; Vonakis, B.M.; Raben, D.M.; Soloski, M.J.; Caterina, M.J. TRPV2 has a pivotal role in macrophage particle binding and phagocytosis. Nat. Immunol. 2010, 11, 232–239. [Google Scholar] [CrossRef] [Green Version]
- Doñate-Macian, P.; Gómez, A.; Dégano, I.R.; Perálvarez-Marín, A. A TRPV2 interactome-based signature for prognosis in glioblastoma patients. Oncotarget 2018, 9, 18400–18409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santoni, G.; Amantini, C. The Transient Receptor Potential Vanilloid Type-2(TRPV2) Ion Channels in Neurogenesis andGliomagenesis: Cross-Talk between Transcription Factors and Signaling Molecules. Cancers 2019, 11, 322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshikawa, F.; Sato, Y.; Tohyama, K.; Akagi, T.; Hashikawa, T.; Nagakura-Takagi, Y.; Sekine, Y.; Morita, N.; Baba, H.; Suzuki, Y.; et al. Opalin, a Transmembrane Sialylglycoprotein Located in the Central Nervous System Myelin Paranodal Loop Membrane. J. Biol. Chem. 2008, 283, 20830–20840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hainz, N.; Wolf, S.; Tschernig, T.; Meier, C. Probenecid Application Prevents Clinical Symptoms and Inflammation in Experimental Autoimmune Encephalomyelitis. Inflammation 2016, 39, 123–128. [Google Scholar] [CrossRef]
- Hainz, N.; Wolf, S.; Beck, A.; Wagenpfeil, S.; Tschernig, T.; Meier, C. Probenecid arrests the progression of pronounced clinical symptoms in a mouse model of multiple sclerosis. Sci. Rep. 2017, 7, 17214. [Google Scholar] [CrossRef]
- Hainz, N.; Becker, P.; Rapp, D.; Wagenpfeil, S.; Wonnenberg, B.; Beisswenger, C.; Tschernig, T.; Meier, C. Probenecid-treatment reduces demyelination induced by cuprizone feeding. J. Chem. Neuroanat. 2017, 85, 21–26. [Google Scholar] [CrossRef]
- Nave, K.A.; Lai, C.; Bloom, F.E.; Milner, R.J. Jimpy mutant mouse: A 74-base deletion in the mRNA for myelin proteolipid protein and evidence for a primary defect in RNA splicing. Proc. Natl. Acad. Sci. USA 1986, 83, 9264–9268. [Google Scholar] [CrossRef] [Green Version]
- Koeppen, A.H.; Barron, K.D.; Csiza, C.K.; Greenfield, E.A. Comparative immunocytochemistry of Pelizaeus-Merzbacher disease, the jimpy mouse, and the myelin-deficient rat. J. Neurol. Sci. 1988, 84, 315–327. [Google Scholar] [CrossRef]
- Vela, J.M.; González, B.; Castellano, B. Understanding glial abnormalities associated with myelin deficiency in the jimpy mutant mouse. Brain Res. Rev. 1998, 26, 29–42. [Google Scholar] [CrossRef]
- Lee, B.C.; Le, D.T.; Gladyshev, V.N. Mammals Reduce Methionine-S-sulfoxide with MsrA and Are Unable to Reduce Methionine-R-sulfoxide, and This Function Can Be Restored with a Yeast Reductase. J. Biol. Chem. 2008, 283, 28361–28369. [Google Scholar] [CrossRef] [Green Version]
- Monet, M.; Gkika, D.; Lehen’Kyi, V.; Pourtier, A.; Abeele, F.V.; Bidaux, G.; Juvin, V.; Rassendren, F.; Humez, S.; Prevarsakaya, N. Lysophospholipids stimulate prostate cancer cell migration via TRPV2 channel activation. Biochim. Biophys. Acta 2009, 1793, 528–539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petković, F.; Campbell, I.L.; Gonzalez, B.; Castellano, B. Astrocyte-targeted production of interleukin-6 reduces astroglial and microglial activation in the cuprizone demyelination model: Implications for myelin clearance and oligodendrocyte maturation. Glia 2016, 64, 2104–2119. [Google Scholar] [CrossRef] [PubMed]
- Golan, N.; Adamsky, K.; Kartvelishvily, E.; Brockschnieder, D.; Möbius, W.; Spiegel, I.; Roth, A.; Thomson, C.E.; Rechavi, G.; Peles, E. Identification ofTmem10/Opalinas an oligodendrocyte enriched gene using expression profiling combined with genetic cell ablation. Glia 2008, 56, 1176–1186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, W.; Yang, W.; Yang, W.; Zhang, J.; Pang, D.; Gan, L.; Luo, L.; Fan, Y.; Liu, Y.; Chen, M. Identification of Tmem10 as a Novel Late-stage Oligodendrocytes Marker for Detecting Hypomyelination. Int. J. Biol. Sci. 2013, 10, 33–42. [Google Scholar] [CrossRef]
- Kippert, A.; Trajkovic, K.; Fitzner, D.; Opitz, L.; Simons, M. Identification of Tmem10/Opalin as a novel marker for oligodendrocytes using gene expression profiling. BMC Neurosci. 2008, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- De Faria, O.; Dhaunchak, A.S.; Kamen, Y.; Roth, A.D.; Kuhlmann, T.; Colman, D.R.; Kennedy, T.E. TMEM10 Promotes Oligodendrocyte Differentiation and is Expressed by Oligodendrocytes in Human Remyelinating Multiple Sclerosis Plaques. Sci. Rep. 2019, 9, 3606. [Google Scholar] [CrossRef]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and Expression of Non-Neuronal Transient Receptor Potential (TRPV) Ion Channels in Rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Maksoud, M.J.; Tellios, V.; Xiang, Y.-Y.; Lu, W.-Y. Nitric oxide displays a biphasic effect on calcium dynamics in microglia. Nitric Oxide 2021, 108, 28–39. [Google Scholar] [CrossRef]
- Platten, M.; Ho, P.P.; Youssef, S.; Fontoura, P.; Garren, H.; Hur, E.M.; Gupta, R.; Lee, L.Y.; Kidd, B.A.; Robinson, W.H.; et al. Treatment of Autoimmune Neuroinflammation with a Synthetic Tryptophan Metabolite. Science 2005, 310, 850–855. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.; Anderson, T.; McCulloch, M.; Dickinson, P.; Vouyiouklis, D.; Griffiths, I. The early phenotype associated with the jimpy mutation of the proteolipid protein gene. J. Neurocytol. 1999, 28, 207–221. [Google Scholar] [CrossRef]
- Ruiz, M.; Bégou, M.; Launay, N.; Ranea-Robles, P.; Bianchi, P.; López-Erauskin, J.; Morató, L.; Guilera, C.; Petit, B.; Vaurs-Barrière, C.; et al. Oxidative stress and mitochondrial dynamics malfunction are linked in P elizaeus- M erzbacher disease. Brain Pathol. 2018, 28, 611–630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, C.; Jia, P.; Jia, Y.; Weissbach, H.; Webster, K.A.; Huang, X.; Lemanski, S.L.; Achary, M.; Lemanski, L.F. Methionine sulfoxide reductase A (MsrA) protects cultured mouse embryonic stem cells from H2O2-mediated oxidative stress. J. Cell. Biochem. 2010, 111, 94–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torkildsen, Ø.; Brunborg, L.A.; Myhr, K.-M.; Bø, L. The cuprizone model for demyelination. Acta Neurol. Scand. 2008, 117, 72–76. [Google Scholar] [CrossRef] [PubMed]
- Van Der Star, B.J.; Vogel, D.Y.; Kipp, M.; Puentes, F.; Baker, D.; Amor, S. In Vitro and In Vivo Models of Multiple Sclerosis. CNS Neurol. Disord. Drug Targets 2012, 11, 570–588. [Google Scholar] [CrossRef] [PubMed]
- Baker, D. Mouse Models of Multiple Sclerosis: Lost in Translation? Curr. Pharm. Des. 2015, 21, 2440–2452. [Google Scholar] [CrossRef]
- Yamate-Morgan, H.; Lauderdale, K.; Horeczko, J.; Merchant, U.; Tiwari-Woodruff, S.K. Functional Effects of Cuprizone-Induced Demyelination in the Presence of the mTOR-Inhibitor Rapamycin. Neuroscience 2019, 406, 667–683. [Google Scholar] [CrossRef]
- Plastini, M.J.; Desu, H.L.; Brambilla, R. Dynamic Responses of Microglia in Animal Models of Multiple Sclerosis. Front. Cell. Neurosci. 2020, 14, 269. [Google Scholar] [CrossRef]
- Gudi, V.; Gingele, S.; Skripuletz, T.; Stangel, M. Glial response during cuprizone-induced de- and remyelination in the CNS: Lessons learned. Front. Cell. Neurosci. 2014, 8, 73. [Google Scholar] [CrossRef] [Green Version]
- Zorzella-Pezavento, S.F.G.; Chiuso-Minicucci, F.; França, T.G.D.; Ishikawa, L.L.W.; da Rosa, L.C.; Marques, C.; Ikoma, M.R.V.; Sartori, A. Persistent Inflammation in the CNS during Chronic EAE Despite Local Absence of IL-17 Production. Mediat. Inflamm. 2013, 2013, 519627. [Google Scholar] [CrossRef]
- Mangiardi, M.; Crawford, D.K.; Xia, X.; Du, S.; Simon-Freeman, R.; Voskuhl, R.R.; Tiwari-Woodruff, S.K. An Animal Model of Cortical and Callosal Pathology in Multiple Sclerosis. Brain Pathol. 2011, 21, 263–278. [Google Scholar] [CrossRef] [Green Version]
- Saunders, C.I.; Kunde, D.; Crawford, A.; Geraghty, D.P. Expression of transient receptor potential vanilloid 1 (TRPV1) and 2 (TRPV2) in human peripheral blood. Mol. Immunol. 2007, 44, 1429–1435. [Google Scholar] [CrossRef] [PubMed]
- Girolamo, F.; Ferrara, G.; Strippoli, M.; Rizzi, M.; Errede, M.; Trojano, M.; Perris, R.; Roncali, L.; Svelto, M.; Mennini, T.; et al. Cerebral cortex demyelination and oligodendrocyte precursor response to experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2011, 43, 678–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kashani, I.R.; Chavoshi, H.; Pasbakhsh, P.; Hassani, M.; Omidi, A.; Mahmoudi, R.; Beyer, C.; Zendedel, A. Protective effects of erythropoietin against cuprizone-induced oxidative stress and demyelination in the mouse corpus callosum. Iran. J. Basic Med. Sci. 2017, 20, 886–893. [Google Scholar] [CrossRef] [PubMed]
- Packialakshmi, B.; Zhou, X. Experimental autoimmune encephalomyelitis (EAE) up-regulates the mitochondrial activity and manganese superoxide dismutase (MnSOD) in the mouse renal cortex. PLoS ONE 2018, 13, e0196277. [Google Scholar] [CrossRef] [Green Version]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Noble, P.G.; Antel, J.P.; Yong, V.W. Astrocytes and catalase prevent the toxicity of catecholamines to oligodendrocytes. Brain Res. 1994, 633, 83–90. [Google Scholar] [CrossRef]
- Thorburne, S.K.; Juurlink, B.H.J. Low Glutathione and High Iron Govern the Susceptibility of Oligodendroglial Precursors to Oxidative Stress. J. Neurochem. 1996, 67, 1014–1022. [Google Scholar] [CrossRef]
- Ohl, K.; Tenbrock, K.; Kipp, M. Oxidative stress in multiple sclerosis: Central and peripheral mode of action. Exp. Neurol. 2016, 277, 58–67. [Google Scholar] [CrossRef]
- Offen, D.; Gilgun-Sherki, Y.; Melamed, E. The role of oxidative stress in the pathogenesis of multiple sclerosis: The need for effective antioxidant therapy. J. Neurol. 2004, 251, 261–268. [Google Scholar] [CrossRef]
- Dowling, P.; Husar, W.; Menonna, J.; Donnenfeld, H.; Cook, S.; Sidhu, M. Cell death and birth in multiple sclerosis brain. J. Neurol. Sci. 1997, 149, 1–11. [Google Scholar] [CrossRef]
- Zipp, F. Apoptosis in multiple sclerosis. Cell Tissue Res. 2000, 301, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Suades, A.; Alcaraz, A.; Cruz, E.; Marimon, E.A.; Whitelegge, J.P.; Manyosa, J.; Cladera, J.; Perálvarez-Marín, A. Structural biology workflow for the expression and characterization of functional human sodium glucose transporter type 1 in Pichia pastoris. Sci. Rep. 2019, 9, 1203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, X.; Touhey, J.; Gaudet, R. Structure of the N-terminal Ankyrin Repeat Domain of the TRPV2 Ion Channel. J. Biol. Chem. 2006, 281, 25006–25010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doñate-Macián, P.; Álvarez-Marimon, E.; Sepulcre, F.; Vázquez-Ibar, J.L.; Perálvarez-Marín, A. The Membrane Proximal Domain of TRPV1 and TRPV2 Channels Mediates Protein—Protein Interactions and Lipid Binding In Vitro. Int. J. Mol. Sci. 2019, 20, 682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marignier, R.; Nicolle, A.; Watrin, C.; Touret, M.; Cavagna, S.; Varrin-Doyer, M.; Cavillon, G.; Rogemond, V.; Confavreux, C.; Honnorat, J.; et al. Oligodendrocytes are damaged by neuromyelitis optica immunoglobulin G via astrocyte injury. Brain 2010, 133, 2578–2591. [Google Scholar] [CrossRef]
- Bolte, S.; Cordelières, F.P. A guided tour into subcellular colocalization analysis in light microscopy. J. Microsc. 2006, 224, 213–232. [Google Scholar] [CrossRef]
- Acarin, L.; González, B.; Castellano, B.; Castro, A.J. Quantitative Analysis of Microglial Reaction to a Cortical Excitotoxic Lesion in the Early Postnatal Brain. Exp. Neurol. 1997, 147, 410–417. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of Structural Proteins during the Assembly of the Head of Bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Laemmli, U.; Favre, M. Maturation of the head of bacteriophage T4: I. DNA packaging events. J. Mol. Biol. 1973, 80, 575–599. [Google Scholar] [CrossRef]
- Valente, T.; Mancera, P.; Tusell, J.M.; Serratosa, J.; Saura, J. C/EBPβ expression in activated microglia in amyotrophic lateral sclerosis. Neurobiol. Aging 2012, 33, 2186–2199. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human Sample | Gender | Age (Years Old) | Post-Mortem Time (h) | Multiple Sclerosis Type | Neurological/Histopathological Evaluation |
|---|---|---|---|---|---|
| Control #1 | Male | 66 | 7 | - | Absence of histological lesions; Absence of neurological disease |
| Control #2 | Male | 70 | n.d. | - | Absence of histological lesions; Absence of neurological disease |
| Control #3 | Female | 74 | 3.33 | - | Absence of histological lesions; Absence of neurological disease |
| Control # 4 | Female | 81 | 23.5 | - | Few Aβ-plaques in entorhinal area; Absence of neurological disease |
| Multiple Sclerosis #1 | Female | 65 | 8 | Secondary progressive | Some chronic active lesions; Many chronic inactive lesions |
| Multiple Sclerosis #2 | Female | 48 | 8.25 | Secondary progressive | Few chronic active lesions; Many chronic inactive lesions |
| Multiple Sclerosis #3 | Male | 46 | 3.25 | Primary progressive | Many chronic active lesions; Few chronic inactive lesions |
| Multiple Sclerosis #4 | Male | 68 | 7 | Secondary progressive | Few chronic active lesions; Many chronic inactive lesions |
| Multiple Sclerosis #5 | Female | 52 | 3.5 | Secondary progressive | Very few chronic active lesions; Many chronic inactive lesions |
| Primary Antibody | Host | Dilution | Reference Manufacturer | Secondary Antibody | Dilution | Reference, Manufacturer |
|---|---|---|---|---|---|---|
| Slot blot | ||||||
| Opalin | Goat | 1:1000 | sc-163187, SantaCruz Biotechnology | Donkey anti-goat IgG-HRP | 1:2000 | sc-2033, SantaCruz Biotechnology |
| NTM | Rabbit | 1:1000 | sc-98979, SantaCruz Biotechnology | Goat anti-rabbit IgG-HRP | 1:2000 | sc-2030, SantaCruz Biotechnology |
| PLP | Rabbit | 1:1000 | sc-98781, SantaCruz Biotechnology | Goat anti-rabbit IgG-HRP | 1:2000 | sc-2030, SantaCruz Biotechnology |
| ICC | ||||||
| GFAP | Mouse | 1:1000 | G3893, Sigma-Aldrich | Alexa 555 donkey anti-mouse | 1:1000 | A31570, Invitrogen |
| CD68 | Rat | 1:250 | MCA1957, AbD Serotec | Alexa 555 goat anti-rat | 1:1000 | A21434, Invitrogen |
| PDGFRα | Rat | 1:100 | 558774, BD Biosciences | Alexa 555 goat anti-rat | 1:1000 | A21434, Invitrogen |
| TRPV2 | Rabbit | 1:200 | sc-31155, SantaCruz Biotechnology | Alexa 488 donkey anti-rabbit | 1:1000 | A21206, Invitrogen |
| Opalin | Goat | 1:50 | sc-163187, SantaCruz Biotechnology | Biotinylated horse anti-goat | 1:500 | BA-9500, Vector Laboratories |
| IHC | ||||||
| TRPV2 | Rabbit | 1:200 | ACC-032, Alomone | Biotinylated horse anti-rabbit | 1:500 | BA-1100, Vector Laboratories |
| MBP | Rabbit | 1:200 | A-623, Dako | Biotinylated horse anti-rabbit | 1:500 | BA-1100, Vector Laboratories |
| MSRA | Rabbit | 1:200 | ab16803, abcam | Biotinylated horse anti-rabbit | 1:500 | BA-1100, Vector Laboratories |
| Opalin | Goat | 1:50 | sc-163187, SantaCruz Biotechnology | Biotinylated horse anti-goat | 1:500 | BA-9500, Vector Laboratories |
| Double IF | ||||||
| TRPV2 | Rabbit | 1:100 | ACC-032, Alomone | Biotinylated horse anti-rabbit | 1:500 | BA-1100, Vector Laboratories |
| APC | Mouse | 1:200 | Calbiochem (#OP80) | Alexa 555 donkey anti-mouse | 1:500 | A31570, Invitrogen |
| WB | ||||||
| TRPV2 | Goat | 1:500 | PA5-18989, ThermoFisher | Mouse anti-goat IgG-HRP | 1:2000 | AP186P, Sigma-Aldrich |
| MSRA | Rabbit | 1:1000 | ab16803, abcam | Donkey anti-rabbit IgG-HRP | 1:5000 | A16035, ThermoFisher |
| β-actin | Mouse | 1:100,000 | A1978, Sigma-Aldrich | Goat anti-mouse IgG-HRP | 1:2000 | 62-6520, ThermoFisher |
| Target Gene | Primer Sequences (5′→3′) | Annealing Temp. (°C) |
|---|---|---|
| MSRA | F: GGC CAT CTA CCC GAC CTC T R: GCC ATT GGG GTT CTT GCT CA | 60 |
| TRPV2 | F: TCA GGT TGG AGA CAT TAG ATG GA R: TCG GTA GTT GAG GTT GAC TCT T | 60 |
| GAPDH | F: CAT GAG AAG TAT GAC AAC AGC CT R: AGT CCT TCC ACG ATA CCA AAG T | 60 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Enrich-Bengoa, J.; Manich, G.; Valente, T.; Sanchez-Molina, P.; Almolda, B.; Solà, C.; Saura, J.; González, B.; Castellano, B.; Perálvarez-Marín, A. TRPV2: A Key Player in Myelination Disorders of the Central Nervous System. Int. J. Mol. Sci. 2022, 23, 3617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073617
Enrich-Bengoa J, Manich G, Valente T, Sanchez-Molina P, Almolda B, Solà C, Saura J, González B, Castellano B, Perálvarez-Marín A. TRPV2: A Key Player in Myelination Disorders of the Central Nervous System. International Journal of Molecular Sciences. 2022; 23(7):3617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073617
Chicago/Turabian StyleEnrich-Bengoa, Jennifer, Gemma Manich, Tony Valente, Paula Sanchez-Molina, Beatriz Almolda, Carme Solà, Josep Saura, Berta González, Bernardo Castellano, and Alex Perálvarez-Marín. 2022. "TRPV2: A Key Player in Myelination Disorders of the Central Nervous System" International Journal of Molecular Sciences 23, no. 7: 3617. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23073617